
Neuromechanism of acupuncture regulating gastrointestinal motility
2020-07-10 07:10ZhiYu
World Journal of Gastroenterology 2020年23期
Zhi Yu
Abstract
Key words: Acupuncture; Gastrointestinal motility; Neuromechanism; Afferent fibers;Autonomic nervous system; Central nervous system
INTRODUCTION
Acupuncture, as a complementary and alternative medical treatment, is used worldwide due to its advantages in terms of efficacy and safety. A literature analysis summarized the clinical diseases and symptoms that could be treated by acupuncture and moxibustion by collecting and analyzing published randomized controlled studies in the PubMed database. This analysis identified more than one hundred diseases and symptoms that were unanimously considered effectively treated, among which gastrointestinal (GI) diseases accounted for a quarter of the diseases[1-3]. GI motility disorder is a common pathological feature in many clinical diseases[4,5],including gastroparesis, irritable bowel syndrome (IBS), functional dyspepsia,pseudo-obstruction, and chronic constipation, causing premature satiety, nausea,vomiting, abdominal pain, bloating, loss of appetite, and other clinical symptoms[6-10].Taking diabetic gastroparesis as an example, as many as 25%-55% of patients with type 1 diabetes have gastroparesis, with a slightly higher incidence rate in patients with type 2 diabetes, causing great suffering and poor quality of life in patients[11]. GI motility disorder is becoming a serious public health problem and a significant burden to society; however, current treatment options for gut dysmotility are limited because of serious side effects[12,13].
In previous decades, many studies confirmed the efficacy of acupuncture for functional GI disorders and more recent medical research has made some progress in determining its mechanisms. The nervous system plays a key role in the benign regulation of GI movement in acupuncture. GI motility is the result of the combined effects of the nervous system, humoral factors, and the myoelectric activity of the GI tract itself, in which the nervous system plays a main role. Neural regulation of GI function depends on the following three levels: The local enteric nervous system(ENS) (endogenous regulation), the autonomic nervous system (ANS), and the central nervous system (CNS) (exogenous regulation). The aim of this review is to discuss the possible neuromechanisms of acupuncture in GI motility, based on its afferent signals and the neural regulation of GI motility.
AFFERENT FIBERS AND RECEPTOR MECHANISM OF ACUPUNCTURE
The effect of acupuncture is based on the somatic afferent signals induced by the insertion of needles into the body manually (manual acupuncture, MA) or electrically(electroacupuncture, EA). Somatic afferent nerve fibers are classified into several types according to their diameter, including A-α, A-β, A-δ, and C-fibers, and it is widely accepted that these fibers underlie the neural mechanism of acupuncture[14].MA stimulation can activate all four groups of somatic afferents and deliver the signal to the CNS, while the type of fibers activated by EA stimulation depends on the intensity[15]. Nevertheless, only when the intensity of stimuli is over the threshold of Aδ and/or C-fibers can acupuncture obviously modulate GI motility[16].
A strong mechanical stimulus applied to the abdominal skin induces an inhibitory effect on jejunal motility, and this effect was proven to be produced by excitation of group IV (unmyelinated) cutaneous afferent nerve fibers in the T10 spinal nerve.Similar stimuli applied to the skin of the upper chest, neck, forepaws, or hind paws elicited a facilitative jejunal reflex that was evoked by excitation of mainly group III(A-δ) cutaneous afferent nerve fibers[17]. Another study indicated that duodenal motility was inhibited by EA at intensities of more than 5.0 mA (suprathreshold of group IV afferent excitation) at acupoints located in the abdominal area, and facilitated by EA at acupoints located in the hind paw with intensities of more than 2.0 mA (subthreshold of group IV, and suprathreshold for groups II + III afferent excitation)[18].
A variety of somatosensory receptors are present in the skin and muscles, including mechanoreceptors, thermoreceptors, and nociceptors. Among them, acid-sensing ion channel 3 (ASIC3) is mainly located in Aβ-fibers innervating the skin and muscle to respond to mechanical stimuli, and transient receptor potential vanilloid-1 (TRPV1) is co-expressed with C-fibers rather than with myelinated nociceptors as a heat and mechanical sensor[19,20]. EA at acupoints located in the abdominal area markedly inhibits gastric motility in an intensity-dependent manner, and this inhibitory effect is alleviated by capsazepine injection (an antagonist of the TRPV1 receptor) orTRPV1 gene knockout with no intensity dependence, indicating that TRPV1 is partially involved in the EA-mediated modulation of gastric motility[21,22]. A similar result for EA was observed on jejunal motility[23]. InAsic3gene deleted mice, the effect of EA at ST36 and CV12 on gastric motility was slightly decreased, with no statistical significance compared with that in wild-type mice[24]. The mean temperature was around 43°C, referring to the thermal activation of Aδ- and C-fibers[25-27]. Nociceptive 43°C and 45°C heat stimuli at ST36 and CV12, rather than non-nociceptive 41°C,produced significant regulatory effects on gastric motility, and this effect was also decreased inTrpv1-/-mice, but not inAsic3-/-mice[28,29]. Results from morphological observations confirmed the role of TRPV1 and ASIC3. Western blotting and immunofluorescence results demonstrated an abundance of TRPV1, TRPV4, and ASIC3 in the anatomical layers of ST36[30]. Components of calcium wave propagation(CWP, the proposed downstream sensing pathway) were also co-expressed with TRPV1. However, only TRPV1 is regarded as a responding channel for acupuncture by sensing peripheral information and conducting signalingviathe CWP and the excitatory phosphorylated glutamate ionotropic receptor NMDA type subunit 1-phosphorylated calcium/calmodulin dependent protein kinase II pathway, offering a comprehensive understanding of the physical stimulation by acupuncture of neurological signaling[31].
AUTONOMIC NERVOUS SYSTEM MECHANISM OF ACUPUNCTURE
The parasympathetic and sympathetic nervous system
Neural networks to control GI motility are positioned at three levels: The parasympathetic and sympathetic nervous system (PNS, SNS), the ENS, and the CNS.Although the intestines are capable of functioning in the absence of extrinsic innervation, movements of the upper GI tract are much more dependent on extrinsic neural pathways, and the motility of the small and large intestines are mainly monitored by the ENS[32,33]. The PNS and SNS comprise one of the factors affecting extrinsic innervation of GI contractility. The SNS provides a predominantly inhibitory effect on GI motility, and in contrast, the PNS exerts both excitatory and inhibitory control over GI motility.
The role of the somatoautonomic reflex, which was demonstrated by Sato and his colleagues in the 1960s[34,35], is now considered the neurological basis of acupuncture in GI motility modulation[36,37], and this modulation exerts a regional-specific effect[38,39](Figure 1). Stimulating acupoints in the abdominal area inhibits gastric, duodenal, and jejunal motility by increasing sympathetic efferent fiber activity, and stimulating acupoints in the limb, which, in contrast, facilitates the above-mentioned gut motility by exciting vagal efferent fiber activity[18,40-43]. The effect of acupuncture on GI motility could be attenuated by vagotomy or sympathectomy[16,44-46]. The β1 and β2 receptors are expressed in the GI tract and serve as the predominant subtypes in the inhibitory effects of the SNS on GI motility[47,48], while the M2 and M3 receptors serve as the predominant subtypes in the excitatory effects of the PNS on GI motility[49,50]. The effect of acupuncture on GI motility could also be reduced by the corresponding antagonist[51-54]and deletion of the gene encoding the β1&2receptor or M2&3receptor[55-57].In addition to the excitement effect, lower extremity acupoints also suggest an inhibitory effect on GI movement, and the inhibitory rate of ST37/LI11 on gastric movement fluctuates between 6.67%-1.33%[58,59]. Moreover, the effect of ST36 on gastric movement is reversed to inhibition after gene knockout of M2 and M3 receptor subtypes[45], suggesting the involvement of other receptors. This might be caused by activation of the other vagus pathway, the inhibitory non-adrenergic, noncholinergic(NANC) pathway, whose transmitter is nitric oxide (NO) or vasoactive intestinal polypeptide (VIP), to cause smooth muscle relaxation[60]. Zhu and his team explained this type of region-specific effect using the spinal segmental innervation theory[46].Stimulation at homotopic acupoints (with the same spinal segment from the innervated visceral organs) decreases GI pressureviathe sympathetic pathways,while stimulation at heterotopic acupoints (with different spinal segments from the innervated visceral organs) facilitates GI motilityviathe parasympathetic pathways[41,56]. This effect applied not only to normal conditions, but also to rats with diarrhea and constipation. It should be noted that this region- specific effect seems to apply only to the regulation of GI movements. When it comes to the effects of acupuncture on modulating the cardiovascular system, stimulations of the limbs,chest, and abdomen all produce a similar effect in terms of heart rate and blood pressure, yet whether the effect is excited or suppressed depends on the intensity of the stimulus (or what type of fiber is activated) and the tissue structure (skin or muscle)[61-64].
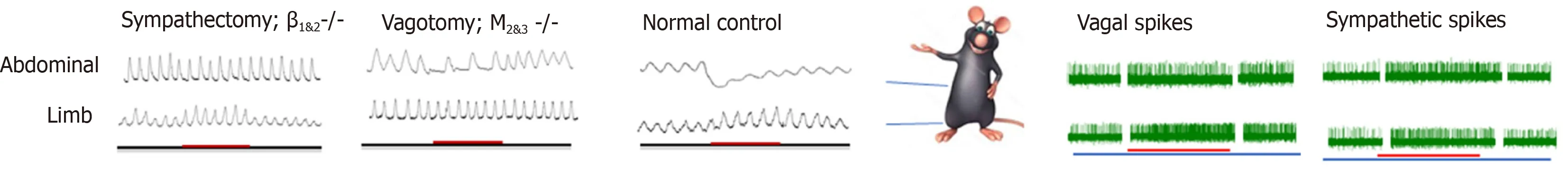
Figure 1 Regional-specific effect of acupuncture on gastrointestinal motility and its autonomic nervous system mechanism. Stimulating the abdominal acupoints inhibits gastric/duodenal/jejunal motility by increasing sympathetic efferent fiber activity; and stimulating acupoints in the limb facilitates motility by exciting vagal efferent fiber activity. The effect of abdominal acupoints on gastrointestinal motility could be attenuated by sympathectomy or deletion of the gene encoding the β1&2 receptor. The effect of limbic acupoints on gastrointestinal motility could be attenuated by vagotomy or M2&3 receptor gene knock out.
The response time of acupoints at different sites might be another feature of the regional specificity of the somatoautonomic reflex; however, this feature is rarely noticed. According to a previous study[65], the abdominal acupoints for gastric motility could be effective within 30 s with a shorter response time, while the acupoint of the hindlimb was effective after 30 s with a longer response time. ST36 showed an excitatory effect in a fast onset manner in sympathectomy rats and β1&2knockout mice,suggesting that sympathetic inhibition might be one of the key factors for the delayed onset of ST36 and β1 and β2 receptors may underlie its receptor mechanisms[44,45].Undoubtedly, the sympathetic pathway is activated by acupoints in the lower limbs;evidence shows that EA at ST36/ST37 increased sympathetic activity in healthy volunteers using a heart rate variability analysis or microneurography evaluation of the left peroneal nerve[66,67]. This mechanism is fully elucidated in terms of the cardiovascular system and anti-inflammatory effects[68,69]; however, it is rarely mentioned in the regulation of GI function. Therefore, the role of sympathetic nerve pathways in regulating GI function by limbic acupoints is greatly underestimated and further evidence is required.
When it comes to colonic motility, the situation is a bit more complicated, because the effects of acupoints located in different areas on the proximal and distal colon are inconsistent. Acupuncture at the acupoints of the forelimb (LI1-LI6 and LI11),abdomen (ST25), and back (BL25) had no significant effects on the proximal colonic motility; and acupoints of the hind paw (ST37) increased proximal colonic pressure[70-72]. Stimulation of the limbs (LI11, ST37, and ST36), abdomen (ST25), and back (ST37) all produced an augmented effect on distal colonic motility regardless of normal, constipated, or diarrheic state[73]. The mechanism underlies the cholinergic pathway[74]. Apart from the segmental innervation theory, the regional neuro-feature between the proximal and distal colon might offer a partial explanation. Vagal innervation to the colon varies among species. In humans, the vagal efferent to the large intestine ends at the splenic flexure, and the remaining part receives parasympathetic input from the pelvic nerves[75-77]. However, studies in rodents suggest that the efferent fibers of the vagus distribute throughout the entire colon[78,79],which is supported by evidence from a retrograde neuronal tracing study[80].Moreover, the distal colon is innervatedviadual parasympathetic nerves, because the pelvic nerve projects from the rectum through the mid colon. In addition, there are regional differences in the innervation of the vagus nerve between the proximal and distal colon[81-84],i.e., the fiber density and distribution of neurotransmitters, such as acetylcholine, within the ganglions is different from each other. It is possible that the regional difference underlies the inconsistency of EA in regulating the motility of the proximal and distal colons. Besides, the regional complexity in the enteric neuron[85]and pelvic ganglion (contain both postganglionic sympathetic and parasympathetic neurons) innervated colon might also add to the complexity of acupuncture-mediated modulation of GI motility[86-88].
ENS MECHANISM OF ACUPUNCTURE
In contrast to the innervation of other peripheral organs, the GI tract has an extensive intrinsic nervous system (the ENS), which modulates its functions, including motility,secretion, blood flow, immunity, and maintenance of the integrity of the epithelial barrier. The major components of the ENS are myenteric plexuses and submucosal plexuses, which contain numerous enteric neurons and glial cells. The myenteric plexuses distribute from the upper esophagus to the internal anal sphincter, forming a continuous network to control GI motility. The submucosal plexuses are closer,allowing them to sense the lumen environment and regulate GI blood flow, as well as controlling epithelial cell functions and secretion[89]. Basically, enteric neurons can be classified into three types according to their function: Motor neurons (including inhibitory neurons and excitatory neurons), sensory neurons, and interneurons.Different neurons in the ENS neural network regulate GI movement by releasing different transmitters. These transmitters include excitatory transmitters, like acetylcholine, opioid peptides, and 5-HT; and inhibitory transmitters such as VIP and NO.
Enteric motor neurons
The importance of the ENS in the neuropathy of GI dysmotility is highlighted by a wide range of diseases that are grouped as congenital, acquired (such as slow transit constipation), and secondary disorders (such as diabetic gastroparesis)[32]. Wedelet al[90]found that the number of nerve cells decreased significantly within the myenteric plexus and external submucous plexus in patients with idiopathic slow-transit constipation by observing the neuronal marker Protein Gene Product 9.5 (PGP9.5),confirming the role of ENS in GI motility. PGP9.5-positive cells were also markedly decreased in the jejunum, ileum, and proximal colon myenteric plexus of mice with constipation[91]. EA has shown a benign regulatory effect in STC; however, much less is known about its mechanism with reference to the ENS, and a series of studies suggested that EA could restore GI motility by controlling excitatory and inhibitory neurons. ST37, an acupoint in the limb, improved the carbon propulsion rates and defecation time and increased the total PGP9.5 expression in the jejunum, ileum, and proximal colonic myenteric plexus[92]. Further results indicated the restoration of enteric neuron function by downregulating the level of nitric oxide synthase (nNOS,secreted by inhibitory neurons to induce relaxation) and upregulating the expression of choline acetyltransferase (ChAT, secreted by excitatory neurons to induce contraction). While ST25, located in the abdomen, could only downregulate the expression of nNOS; therefore, the regulatory effect of EA on enteric neurons seems to act in a region-specific manner[72](Figure 2).
Loss of enteric neurons is one the most important neuropathies that contribute to GI dysmotility in diabetic rats, and this change may be mediated, in part,viaa reduction of glial cell derived neurotrophic factor (GDNF) and its main downstream signaling pathway phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3K)-protein kinase B (AKT), which is a survival signal for enteric neurons[93,94]. Decreases in PGP9.5, nNOS, and ChAT neurons in the colon of diabetic rats led to disordered gastric emptying and intestinal transit[95,96]. After long-term intervention, EA at ST36 induced regeneration of lost enteric neurons[97]. Meanwhile, the mRNA and protein level of GDNF, an important neurotrophic factor of enteric neurons, and p-AKT in the colon were upregulated, suggesting the role of GDNF and the PI3K-AKT signal pathway in the mechanism of EA-induced regeneration of impaired enteric neurons.
Enteric sensory neurons
Purinergic P2X receptors contribute to neurotransmission in subsets of enteric neurons of the GI tract and P2X3 receptor immunoreactivity was found in the myenteric neurons of the esophagus, stomach, and small and large intestines of rodents, of which the major types of neurons are excitatory, inhibitory muscle motor neurons, and interneurons[98,99]. However, the evidence for P2X3 receptor involvement in GI motility is limited. Studies have reported thatP2X3gene knockout mice exhibit a feature of impaired gut peristalsis[100,101]. P2X3 subunits are localized to after hyperpolarizing neurons[102], which are likely to be intrinsic intestinal sensory neurons. The impairment of peristalsis caused byP2X3gene deletion was independent of the changes in the muscarinic receptor or nicotinic receptor function in the longitudinal muscle layer, suggesting that the P2X3 receptor underlies the sensory mechanisms in the myenteric plexus that mediate peristalsis, and plays an important role in mechanosensitivity and hypersensitivity[103]. The therapeutic effect of EA on IBS has been confirmed by a substantial body of literature from clinical data,and the mechanism involves alleviating mechanosensitivity and hypersensitivity[104],where P2X receptors might serve as novel targets[105]. Electroacupuncture has been suggested to regulate the P2X3 receptor located in the neurons of the colonic myenteric plexus of IBS rats[106]. Whether this has a crucial role in the other parts of the GI tract has not yet been reported, and the results of this study[104]provide clues for further research.
Enteric glia cells
The ENS contains a large amount of enteric glial cells (EGCs) surrounding neurons to regulate neuronal activity and protect the gut in different ways, including maintaining the intestinal barrier function and the inflammatory process, which facilitate the control of GI motility[107]. Animals with genetic elimination of enteric glia develop jejuno-ileitis inflammation[108]and intestinal dysmotility in a sex-dependent manner[109], confirming the pivotal role of EGCs in GI motility disorders[110].
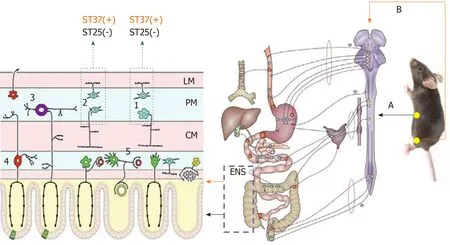
Figure 2 The enteric nervous system mechanism of acupuncture. Stimulation at ST37 and ST25 had different effects on colonic motility in terms of the effects on excitatory and inhibitory neurons in the enteric nervous system. A: Sympathetic pathway; B: Parasympathetic pathway; 1: Inhibitory neurons; 2: Excitatory neurons; 3:Myenteric intrinsic primary afferent neurons; 4: Submucosal intrinsic primary afferent neurons; 5: Secretomotor/vasodilator neurons; LM: Longitudinal muscle; MP:Myenteric plexus; CM: Circular muscle; (+): Effective, (-): Ineffective[72]; ENS: Enteric nervous system.
Hemorrhagic shock is frequently accompanied by bowel stasis following intestinal ischemia and reperfusion injury. The initiated inflammatory response, including the production of cytokines and recruitment of leukocytes within the intestinal muscularis, contributes to impaired muscle contractility and mucosal barrier function[111]. In addition to promoting gastric emptying, EA at ST36 was also reported to decrease intestinal injury and alleviate inflammation to prevent intra-abdominal adhesion, which serves as one of the main causes of intestinal obstruction[112,113]. This protective effect provided by EA acts partlyviaactivation of EGCs to secrete substances that regulate the gut functionviaactivation of the cholinergic antiinflammatory pathway[114,115], because both blockage of the cholinergic α7nAChR subunit by α-bungarotoxin and vagotomy could weaken or eliminate the effects of EA at ST36.
Apart from the above-mentioned function, enteric glia are emerging as novel regulators of enteric reflex circuits that contribute to the regulation of GI motility[116].Delvalleet al[117]provided the first evidence that EGCs have effects on gut motility through modulation of excitatory reflexes involved in GI motility, and activation of glial M3 and M5 subtypes of muscarinic receptors contributes to this kind of physiological regulation. Genetic ablation of M2 and M3 receptors diminished the enhancement of GI motility by EA at ST37[53]; however, whether the subtypes of muscarinic receptors of EGCs are involved requires more evidence.
CNS MECHANISM OF ACUPUNCTURE
As described previously, the extrinsic neural pathway participates in the modulation of GI functions. The CNS, as another important factor of extrinsic innervation,provides its extrinsic neural inputs to control GI motility in a more widespread and integrated manner, which involves the spinal cord, medulla, midbrain, and mesencephalon. The regulatory effect of acupuncture on GI movement also requires the participation of the CNS at multiple levels by changing the activities of the nuclei related to GI movement, including the dorsal motor nucleus (DMV), the solitary tract nucleus (NTS), the raphe nuclei, the lateral hypothalamic area (LHA), and the paraventricular nucleus of the hypothalamus (PVN), which have been confirmed by neuroanatomical tracing following the injection of a neuronal tracer into the stomach and ST36, respectively[118,119]. The role of acupuncture on the nuclei of the CNS is summarized in Table 1.
Brain stem
The dorsal vagal complex: A large volume of sensory information of the GI tract, both mechanical and chemical, is transmitted mainly to the NTSviathe vagal afferent fibers. This information is integrated by NTS neurons and then conveyed to the motor DMV, using γ-aminobutyric acid (GABA), glutamate (Glu), and norepinephrine as the main neurotransmitters to modulate vagal efferent innervation and GI motility through the vagal ACh or NANC pathways[120-122]. The NTS and DMN constitute the dorsal vagal complex (DVC), the main neuroanatomical structure of vagovagalreflexes. The role of DVC in acupuncture-mediated regulation of GI function is supported by multiple pieces of evidence from morphological, neuroelectrophysiological, and neurochemical studies. While regulating the electro activity of the stomach, EA at ST36 and ST37 could modulate the firing of the neurons in the NTS and DMV, and increase their Fos expression[46,123-126]. Moreover, with the help of a lesion-making device, the DVC was damaged, which resulted in GI tract dysfunction[127], and EA lost its regulatory effect on gastric motility, confirming the important role of DVC in EA-mediated regulation of GI function[123,128].

Table 1 The role of acupuncture on the nuclei of the central nervous system
Brain-gut peptides play an important role in GI function. There is a long list of GI peptides that are capable of regulating GI motility. They are secreted by endocrine cells of the gut as neurotransmitters and neuromodulators, and distribute to the CNS to regulate GI functions[129-131]. A fewer years later, another study demonstrated that EA at RN12 and BL21 enhanced the production of GI hormones, such as motilin and gastrin, in the DVC[126]. In addition, acupuncture stimulation at ST36 could modulate the concentration of VIP, motilin, ghrelin, and gastrin in serum[132]. Microinjection of substance P (SP), another candidate brain-gut peptide, into the DVC inhibited gastric motilityviathe NK-1 receptor, whereas EA at ST36 enhanced gastric myoelectric activity and simultaneously inhibited the release of SP in the DVC, suggesting that SP in the DVC is involved in the effect of EA on gastric myoelectric activity[133]. These data implied the role of neurons in the DVC in the effect of EA on GI movement. The activities of brain-gut peptides are complicated, and their mechanisms differ from each other. In the detection of the same brain-gut peptide in patients, different scholars may have drawn different conclusions, and have not mentioned their complex interconnections. An in-depth study of brain-gut peptides and their relationship with acupuncture would be conducive to a better understanding of the mechanism of acupuncture in GI motility.
Glia constitute almost half of the cells in the CNS. Given the emerging evidence,glial cells have been highlighted in many aspects of CNS function, rather than acting as static bystanders as was long believed. Continued elucidation of glial cell biology,and the dynamic interactions of neurons and glia, will enrich our understanding of nervous system health and diseases[134]. Research on acupuncture-mediated regulation of glial cells is increasing year by year, in such fields as analgesic processes, antiinflammation, and GI function[135-137]. Studies suggest that EA at ST36 significantly increased the expression of astrocytic (glial fibrillary acidic protein) and microglial(OX-42) marker cells in the DVC and regulated gastric activity. Propentofylline (PPF,a blocker of both microglial cells and astrocytes) attenuated the activation by EA of gastric activity, suggesting that EA regulation of gastric activity might be related to the glia in the DVC[138]. Although the function of DVC glia in the effect of EA on GI motility has been suggested, the data remains very limited, and the hypothesis must be verifiedviafurther studies.
Vagovagal neurocircuits comprise specific nuclei, including the NTS and DMV.NTS neurons receive and converge the somatic and visceral sensory information[139-141],whereas the DMV is the origin of vagal efferent fibers, modulating gastric motility as well as other visceral functions (e.g., vagal neurocircuitry and its influence on gastric motility). GABA is the main neurotransmitter that transmits information from the NTS to the DMV. Injection of L-Glu into the DMV might lead to a change in gastric activity[142-144], and whether the effect is excited or suppressed depends on the injection site (rostrum or posterior)[145]. Microinjection of GABA into the DMV decreased gastric motility, whereas microinjection of bicuculline, a GABA antagonist, increased gastric motility[146,147]. EA at PC6 showed no marked change after the injection of L-Glu;however, gastric movement and the parasympathetic nerve firing after injection of GABA were enhanced, indicating that GABA transmission of the DMV underlies the facilitation of EA at PC6 on gastric motility[44]. EA at ST36[45]is capable of modulating gastric motility after the injection of L-Glu and GABA, suggesting that both Glu and the GABA receptor in the DMV participate in promoting the gastric motility effect of EA at ST36,i.e., EA at ST36 and PC6 modulate gastric motilityviathe vagovagal reflex.
The rostral ventrolateral medulla: The rostral ventrolateral medulla (RVLM), the final area for the integration of sympathetic nerve activity (SNA)[148], processes a large volume of pre-sympathetic (sympathoexcitatory) neurons that project directly into the intermediolateral (IML) column of the spinal cord, and modulates the sympathetic control of visceral functions[149]. Takahashiet al[150]reported that acupuncture at ST-36,but not at ST25, increased the number ofc-Fosimmunopositive cells at the NTS and DMV, while acupuncture at ST25 specifically increased the number of cells at the NTS and RVLM. This suggested that somatic afferents activated by acupuncture at ST36 are conveyed to the NTS and then stimulate the DMV neurons. In contrast, somatic afferents activated by acupuncture at ST25 are conveyed to the NTS and stimulate RVLM neurons. This deduction was supported by anatomical and functional evidence that demonstrated a robust excitatory input from NTS neurons to neurons within the RVLM[151]. In spinal functional magnetic resonance imaging research, multiple activations of grey matter (including the spinal dorsal horn and IML) in T8 and T9 vertebrae showed a response to stimulation on ST25[152]. Another study found that ST25, ST37, and LI11 could all change the firing rate of the IML; however, ST25 showed the strongest effect. It could be concluded that the NTS-RVLM-IMLsympathetic and NTS-DMV-parasympathetic pathways, respectively, underlie the neural anatomical evidence of regional specific effects of acupuncture on gastric motor function in rats. However, RN12, BL 21, and ST25 are located in the abdomen and back, and share similar segment innervation, yetc-fosexpression in the NTS and DMV was statistically increased by EA stimulation at RN12 and BL21; therefore, this anatomical evidence of a regional specific effect requires further research for verification.
The cerebellum
The fastigial nucleus: The cerebellum is traditionally viewed as a subcortical motor center; however, behavioral, anatomical, and neuroelectrophysiological studies have shown that the cerebellum also has an important regulatory role in the visceral activities of the body[153]. The fastigial nucleus (FN) is the oldest nucleus in the cerebellum and inhabits a key position in the regulation of nonsomatic functions,including the GI movement and feeding, among the nuclei of the cerebellum[154,155].Electrical stimulation of the cerebellum can affect GI movement through the vagal and sympathetic nerve pathways[156-158]: Activation of adrenergic fibers to release catecholamines, and activation of cholinergic fibers to inhibit or facilitate gastric movement. In addition, studies have shown that electrical stimulation of the FN has a protective effect on the gastric mucosa by reducing gastric ischemia-reperfusion injury and apoptosis of gastric mucosal cells in rats. Needling at ST36, LI11, BL21, and CV12 could elevate the spontaneous discharge frequency of cerebellar FN neurons;however, CV12 seems to have an advantageous role in regulating the cerebellar FN,partly confirming the role of the FN in the central pathway of acupuncture-mediated regulation of GI function[159].
The direct bidirectional pathways between the cerebellum and the hypothalamus
have been demonstrated in a series of neuroanatomical investigations, forming a part of the circuits underlying cerebellar autonomic modulation[160,161]. The FN is one of the originating nuclei of the cerebellar-hypothalamic projection, and is the main structure that modulates GI activities in the cerebellum. Recently, the FN-LHA loop has been investigated[162], and is likely to serve as the neural basis for regulating GI functions[163].Gastric vagal and cerebellar fastigial nuclear signals converge on glycemia-sensitive neurons of the LHA in rats[164]. Electrical and chemical stimulation of the FN had neuroprotective effects on gastric mucosal stress injury and gastric ischemiareperfusion damage in rats, and pre-damage of the FN or LHA could eliminate the protective effect of the FN[165-169]. Extracellular recording results suggested that the signal of gastric distention (GD) and acupuncture at ST36 converged in the LHA and FN, and acupuncture could modulate the activities of different types of GD-sensitive neurons in the LHA and FN, indicating that the LHA-FN circuit might participate in the central integration mechanism of acupuncture’s effects on gastric function[170].However, whether the LHA-FN circuit, which underlies the effect of acupuncture on GI function, acts through the sympathetic nerve or parasympathetic nerve requires further research.
The vestibular nucleus: The vestibular nucleus (VN) is the initial structure of the central processing of vestibular orientation information and receives projections from the vestibular nerve, playing a special role in regulating GI function. The work of Gagliusoet al[171]and Ruggieroet al[172]demonstrated the neuroanatomy of the vestibulo-solitary pathway, providing an anatomical explanation for the autonomic changes observed, including GI responses, after peripheral vestibular activation[171,172].The medial vestibular nucleus (MV) is the area with the largest volume and cell number in the vestibular nucleus. Direct fiber projection also exists in the MV and dorsal vagus nucleus[173,174]. The technique of retrograde axonal transport of horseradish peroxidase in the MV identified a group of neurons sending axons to the"stomach" region of a single tract nucleus, and local irritation of the neurons initiated relaxation of the stomach wall[175]. GD and acupuncture at different acupoints (ST36,LI11, and BL21) modulated the activity of the neurons within the MVN, suggesting that the MVN is partly involved in the central integration mechanism underlying acupuncture-mediated regulation of gastric functions[176].
The hypothalamus
The hypothalamus is located on the ventral thalamus and contains distinct groups of nuclei and bundles that regulate a variety of autonomic functions by interconnecting with other parts of the CNS. Among them, the PVN, the LHA, and the arcuate nucleus (ARC) are particularly important in the regulation of GI functions.
The paraventricular nucleus of the hypothalamus: The PVN is located near the periventricular zone of the medial hypothalamus, and emerges as one of the most important autonomic control centersviaits secretion of a variety of peptides[177,178].Among them, corticotrophin-releasing hormone (CRH) and oxytocin (OT),synthesized by magnocellular and parvocellular neurons within the PVN for release into the hypophysial portal system or projection to other central nuclei, are particularly important in the regulation of GI functions, especially stress-induced alterations of GI motility[179,180]. Studies have shown that corticotropin-releasing factor(CRF) activates DMV neurons and decreases gastric motilityviaactivation of the vagal NANC pathway[181]. Few data are available focusing on acupuncture-mediated regulation of CRH function, and these data suggest that acupuncture could activate CRH-like neurons and modulate the expression of CRH and the glucocorticoid receptor (GR) in the PVN[182]. Rinamanet al[183]showed that the PVN is the sole source of OT-immunopositive fibers and terminals in the DVC. Microinjection of OT into the DMV or PVN resulted in inhibition of gastric motility; however, this effect was ameliorated by vagotomy or microinjection of an OT receptor antagonist into the PVN[184]. Llewellyn’ s work provided the anatomical basis for the functional pathways in GI motility regulation by OT: As a neurotransmitter, OT is transmitted from the PVN to the DVC, where it activates motor neurons of the brainstem vagus nerveviathe NANC pathway, thereby inhibiting GI movement[185,186]. EA at the limbic acupoints ST36 and PC6 increased the intragastric pressure and decreased the numbers of double-labeled OT neurons andc-fosneurons within the PVN[187]. However, the effect of acupuncture was significantly decreased after dorsolateral funiculus or spinal cord transection, suggesting that the dorsolateral funiculus is the main conduction bundle responsible for acupuncture signal transduction into the PVN[188]. Knockout of theOTgene accelerated gastric emptying, and influenced the effect of acupuncture, further confirming the paraventricular nucleus OT neurons as the initiating device of autonomic nervous pathways, which might participate in EA-mediated regulation of gastric motility, in the form of neural and neuroendocrine stimuli[189]. Thus,acupuncture signals gather in the DVC and PVN. PVN lesions cause a decrease in gastric motility, and EA is unable to restore it, demonstrating that the PVN is another central target of acupuncture’s regulation of gastric motility[190]. Taken together, these data suggest that the acupuncture signal is transmitted to the solitary nucleus through the spinal cord, and after initial integration by the solitary nucleus, the ascending projection activates the PVN. The descending projection is performed by the PVNDVC-vagal nerve pathway, through the nerve system and neuroendocrine system to regulate GI function.
The lateral hypothalamic area: The LHA is linked to celiac sympathetic and parasympathetic nerves, and regulates metabolic, behavioral, and autonomic functions, and serves as the feeding center[191]. Its influence on GI function and feeding behavior might be mediated by the DVC. As early as the 1980s, with the help of tracing techniques, an anatomical connection was found between the LHA and the dorsal DVC[192,193]. Moreover, this pathway involves multiple neurotransmitters and neuropeptides, including the projection of ghrelin neurons[194-196]. Ghrelin has central and peripheral actions in gut motility, and its receptors are regarded as targets for novel motility drugs[197,198]. Ghrelin fibers originating in the LHA project into the DVC.The electrical stimulation of the LHA promotes gastric motility and GD neurons in the DVC, and this excitatory effect is partially blocked by pretreatment with a ghrelin receptor antagonist and is diminished by an electrical lesion of the DVC. Cisplatintreatment could weaken this excitatory effect of electrical stimulation in the LHA.
Somatic input from ST36 abolished the inhibitory reaction of the LHA induced by gastric distention[170,199]. Acupuncture could modulate the levels of noradrenaline,serotonin (5-HT), and the activity of ATPases in the LHA[200]. However, ST36 and ST25 are able to alter the activity of the glucose-sensitive neurons within the LHA, which is also the candidate target of ghrelin[201-203]. Increasing numbers of investigations have focused on the role of acupuncture in the peripheral actions of ghrelin in gut motility in the clinic[204,205]. The level of plasma ghrelin was measured in patients with GI cancer and obesity[206,207]and in mice. The results suggested that acupuncture has a benign effect on the modulation of ghrelin secretion. However, the literature on the central role of acupuncture on ghrelin secretion is limited. It has been confirmed that acupuncture can activate the TRPV1 receptor[21,23,30,31], which can also be activated by capsaicin during GI motility,i.e., acupuncture has a similar effect to capsaicin.Capsaicin regulates the central action of ghrelin[194-196]; therefore, we speculated that acupuncture also has a central regulatory effect on ghrelin, which requires further data to prove this.
Hypothalamic arcuate nucleus: The ARC is located at the base of the hypothalamus and adjacent to the third ventricle. It receives extensive afferent inputs and processes efferent fibers projecting to other central regions, underlying its role in the regulation of autonomic functions, including GI functions[208-210]. Stimulation of neurons in the ARC has been demonstrated to increase colonic motility[211]and inhibit gastric phasic contractions[212]as well as gastric acid secretionviavagal pathways[213]. Its effect on the colon could be blocked by microinjection of CRF antagonists into the PVN, suggesting that an Arc-PVN neurocircuit might be involved in the regulation of GI functionsviavagal efferent fibers.
The role of ARC in the modulation of acupuncture has been demonstrated;however, most research has been confined to the area of analgesia[214]. Applying the tracing technique, the afferent pathways of CV4 were explored. After the injection of Pseudorabies virus (PRV) into the acupoint of CV4, nuclei such as ARC and PVN showed PRV-immunoreactive neurons in normal rats[215]. Other results from Fos-CreER-based genetic mapping indicated that acupuncture at PC6 activates a series of regions, including the ARC and PVN[216]. These studies confirmed that ARC could receive the somatic fibers of acupuncture and modulate its activity. Following ligation of the ARC, the effect of acupuncture on the electrogastrography of rabbits was attenuated[217], providing anatomical and physiological evidence for the ability of acupuncture to regulate GI motilityviathe ARC. EA treatment increased the peptide levels of the melanocyte-stimulating hormone receptor (α-MSH) and the mRNA levels of its precursor proopiomelanocortin (POMC) in the ARC of hypothalamic neurons of rats with diet-induced obesity[218]. In addition, acupuncture also regulates the expression of neuropeptide Y, a cocaine and amphetamine-regulated transcript peptide[219,220], together with POMC neurons in the ARC, playing an important role in feeding and gastric emptying, which is potentially in a pathological state[221,222];therefore, feeding-related peptides in the ARC might underlie another mechanism by which acupuncture regulates GI motility.
CONCLUSION
Although marked progress has been made in revealing the neural mechanism underlying acupuncture’s effects on the GI tract, there is still a lot of work to be done to unravel the complex interaction between the peripheral and CNSs, as well as the distinct CNS areas related to the integration of GI homeostatic functions. Acupuncture is a physical stimulus. When the needle penetrates the body, the mechanical force generated directly or indirectly acts on the acupoint area. The mechanical stimulation translates into neurochemical signals, inducing a somatic afferent signal; however,only when the intensity of the stimulus is over the threshold of the Aδ and/or Cfibers can acupuncture obviously modulate GI motility, whose receptor mechanism mainly relies on TRPV1. In the process of acupuncture, a regional-specific effect is exhibited in the modulation of GI motility, the neurological basis of which is now considered to be the somatoautonomic reflex, and the spinal segmental innervation hypothesis could be borrowed to explain it: Stimulation at homotopic acupoints (with the same spinal segments from the innervated visceral organs) decrease the GI pressureviathe sympathetic pathways (mainly involving β1 and β2 receptors), while stimulation at heterotopic acupoints (with different spinal segments from the innervated visceral organs) induces GI facilitationviaparasympathetic pathways(both cholinergic and NANC vagal efferents are involved). Other than that, the ENS,including enteric neurons and glia cells, also adds to the effect of acupuncture on the modulation of GI motility. The CNS provides its extrinsic neural inputs to control GI motility in a more widespread and integrated manner. The regulatory effect of acupuncture on GI movements also requires the participation of the CNS at multiple levels. In this review, only some nuclei of the brain stem, cerebellum, and hypothalamus are discussed, because of the complexity of the CNS and the blurred relationship between distinct nuclei in the CNS and acupuncture. Those connections among the nuclei constitute the anatomical and functional basis controlling GI motility. The acupuncture signals over GI functions were also analyzed and integrated through this connection. Applying modern and constantly updated technology is conducive to gaining a better understanding of the mechanism of acupuncture’s effects on GI motility.
 World Journal of Gastroenterology2020年23期
World Journal of Gastroenterology2020年23期
- World Journal of Gastroenterology的其它文章
- Liver-directed therapies for liver metastases from neuroendocrine neoplasms: Can laser ablation play any role?
- Potential of the ellagic acid-derived gut microbiota metabolite - Urolithin A in gastrointestinal protection
- Endosonographic diagnosis of advanced neoplasia in intraductal papillary mucinous neoplasms
- Medications in type-2 diabetics and their association with liver fibrosis
- Pancreatic necrosis and severity are independent risk factors for pancreatic endocrine insufficiency after acute pancreatitis: A long-term follow-up study
- Impact of a national basic skills in colonoscopy course on trainee performance: An interrupted time series analysis