
紫苞鸢尾的种子休眠特性研究
2020-07-09 11:16古力尼哥塔依尔阿马努拉依明尼亚孜王振振迪利夏提哈斯木
种子 2020年6期
古力尼哥·塔依尔, 阿马努拉·依明尼亚孜, 王振振, 迪利夏提·哈斯木
(新疆农业大学草业与环境科学学院,自治区草地资源与生态重点实验室, 乌鲁木齐 830052)
种子休眠是指具有活力的种子在适宜的条件下不能萌发的现象,是植物在长期进化过程中形成的抵抗外界不良环境条件的适应性[1-2],对保持物种自身繁衍具有重要意义[3]。为了适应不同的环境,植物在长期的进化过程中选择形成了不同的种子休眠特性[4]。Baskin等[1]把种子休眠归纳为形态生理休眠、形态休眠、生理休眠、物理休眠和复合休眠五种类型。影响种子休眠的因素有种皮机械阻碍、胚发育不全、抑制物存在以及外界环境条件限制等[3]。
目前,有关鸢尾属种子休眠的研究集中在种子休眠的原因、休眠类型以及打破休眠[5-8]的方法等方面。相关研究表明,该属植物种子大多具有较低的萌发率[9],多数种子的休眠属于生理休眠和形态生理休眠[2]。紫苞鸢尾(Irisruthenica)是鸢尾科鸢尾属多年生草本克隆植物,鲜草有毒,在春夏季节家畜不采食,在新疆北部退化山地草原上大面积蔓延[10-11]。对紫苞鸢尾种子萌发实验表明,其种子存在休眠[12],而有关休眠类型和休眠特性的研究未见报道。本研究以紫苞鸢尾为材料,采用野外观测与控制实验相结合的方法,对种子进行室内干藏、赤霉素(GA3)和野外埋藏处理,观测各处理的萌发率,以明确种子休眠类型和休眠特点,为控制该物种在新疆退化山地草原上种群的扩张及蔓延提供基础实验数据。
1 研究材料与地点
供试种子于2017年7月在新疆乌鲁木齐县庙尔沟乡谢家沟山地草原采集,采集地点:87°37′21 E,43° 47′07 N,海拔高度1 600~1 800 m。年平均气温3.3 ℃,最高温度为42 ℃,最低温度为-41 ℃,年均无霜期179 d,属于中温带大陆性气候。
2 研究方法
2.1 室内干藏对打破休眠的影响
将在室温条件下贮藏0(新采收的种子)、2、4、6个月和12个月的种子消毒处理后,置于垫有2层滤纸、直径9 cm的培养皿中,每个培养皿内放置25粒种子,模拟原生境不同季节的平均日最高和最低温度,在30 ℃/15 ℃、25 ℃/15 ℃、20 ℃/10 ℃、15 ℃/2 ℃和5 ℃/2 ℃光照/黑暗(12 h光周期)及全黑暗条件下进行培养,4个重复。每24 h统计1次萌发数,持续观测28 d;在全黑暗条件下培养的种子待萌发结束后(第28天)统计最终萌发数。
2.2 GA3处理对打破休眠的影响
在不同时间段,将贮藏0、2、4、6个月和12个月的种子,置于GA3浓度分别为0.1、1 mmol·L-1和5 mmol·L-1的培养皿中,每个培养皿内放置25粒种子,4个重复。将以上3种处理的种子分别在5个变温、光照/黑暗及全黑暗条件下进行种子萌发观察,对萌发状况进行统计(方法同2.1)。为了防止水分和GA3溶液的蒸发,将培养皿用封口膜进行密封。
2.3 野外埋藏对打破休眠的影响
为了确定野外埋藏及埋藏时间对打破休眠的影响,取10 000粒室内干藏的种子并将其分别装入10个尼龙袋中,将尼龙袋浅埋至盛满沙子的花盆中,深度约为3 cm,置于野外实验地,保证土壤湿度。将埋藏1、2、4、6个月和12个月的种子定期取回,分别在5个变温、光照/黑暗及全黑暗条件下进行种子萌发观察,对萌发状况进行统计(方法同2.1)。

图1 干藏处理光照/黑暗及全黑暗条件下的种子萌发状况
2.4 数据分析
对所测数据用SPSS 23.0统计分析软件进行处理分析。
3 结果与分析
3.1 室内干藏对打破休眠的影响
室温条件下贮藏0个月(新采收的种子)种子的萌发试验显示,5个温变(光照/黑暗)的萌发率分别为34.4%、82.2%、14.4%、5.4%和0%(图1),25 ℃/15 ℃为该物种最适萌发温度。光照/黑暗条件下,储藏0~6个月,对25 ℃/15 ℃种子萌发率的影响不显著,储藏12个月后,其萌发率达到100%;随着贮藏时间的延长,30 ℃/15 ℃、20 ℃/10 ℃、15 ℃/2 ℃和5 ℃/2 ℃光照/黑暗条件下种子萌发率呈显著增加趋势,储藏12个月后,以上4个变温条件下的种子萌发率均超过80%。在全黑暗条件下,随着贮藏时间的延长,25 ℃/15 ℃下种子萌发率呈显著增加趋势,12个月后,其萌发率达到80%;贮藏时间对30 ℃/15 ℃和20 ℃/10 ℃全黑暗条件下的种子萌发率有所影响,而对15 ℃/2 ℃和5 ℃/2 ℃种子的萌发率影响不显著(图1)。
3.2 GA3处理对打破休眠的影响
0.1、1 mmol·L-1和5 mmol·L-1浓度GA3处理对储藏0、2、4、6个月和12个月种子萌发率变化的实验结果表明: 1) 用1 mmol·L-1浓度GA3处理后,各变温、光照/黑暗及全黑暗条件下的萌发率有显著提升,且随着储藏时间的延长,储藏12个月种子萌发率有显著提升; 2) 用0.1 mmol·L-1浓度GA3处理后,光照/黑暗储藏6个月和12个月25 ℃/15 ℃种子有显著变化,所有25 ℃/15 ℃全黑暗条件下种子萌发率有较显著变化,其它种子变化不明显; 3) 用5 mmol·L-1浓度GA3处理后,对光照/黑暗以及全黑暗条件下种子萌发率有显著及较显著的抑制作用; 4) 贮藏12个月后,25 ℃/15 ℃(光照/黑暗)GA30.1、1 mmol·L-1和5 mmol·L-1浓度下的萌发率分别为98.8%、99%和77.5%,在全黑暗条件下的萌发率分别为81%、99%和60%(图2)。
3.3 野外埋藏对打破休眠的影响
野外埋藏1、2、4、6个月和12个月的种子在光照/黑暗和全黑暗5个变温条件下种子萌发率实验结果表明:光照/黑暗25 ℃/15 ℃埋藏1、2、4个月和30 ℃/15 ℃、20 ℃/10 ℃埋藏6个月种子的萌发率有显著提升;埋藏1、2、4、6个月25 ℃/15 ℃全黑暗种子萌发率有显著提升;其它处理条件下的种子萌发率无显著变化;贮藏12个月25 ℃/15 ℃光照/黑暗条件下的种子萌发率为100%,贮藏12个月25 ℃/15 ℃全黑暗条件下的种子萌发率为91%。说明种子萌发率随着野外埋藏时间的增加呈现上升趋势(图3)。
4 讨 论
种子休眠与环境因子以及物种特有的属性密切相关[6,13,14]。多数鸢尾属植物的种子具有休眠现象,且不同种的休眠类型存在较大差异[5,7]。导致鸢尾属植物休眠的原因主要有种皮厚、种子内存在抑制物质、胚的发育存在后熟作用以及外界环境条件限制等[5,7,8]。室内干藏、低温层积、野外埋藏和赤霉素处理是确定其休眠类型的重要依据[15,16]。紫苞新鲜种子仅在25 ℃/15 ℃条件下萌发率为82.2%,而其它变温条件下的萌发率均低于34.4%,说明其种子存在休眠现象;同时,该物种种子具有较好的吸水性[11],说明种皮不是造成休眠的主要原因,种子可能不存在物理休眠;室内干藏试验结果显示,该物种种子随着室内干藏时间延长,光照及全黑暗各变温条件下种子的萌发率均呈现不同程度的增加(图1),可能是由于该物种种子存在胚的后熟作用,在储藏过程中随着胚的成熟种子萌发率不断提高。植物激素是调节种子休眠与萌发的关键因子,是解除种子休眠的重要方法[17-19]。用0.1、1、5 mmol·L-1浓度GA3处理储藏0、2、4、6个月和12个月种子萌发率变化的实验结果表明:用GA3处理紫苞鸢尾种子对打破其休眠有一定的作用,作用大小随着种子储藏时间的延长而增加。种子的生理休眠是最为普遍的休眠类型,可分为非深度生理休眠、中度生理休眠和深度生理休眠[17,19]。综上表明,该物种种子存在休眠,其种子休眠类型为非深度生理休眠。
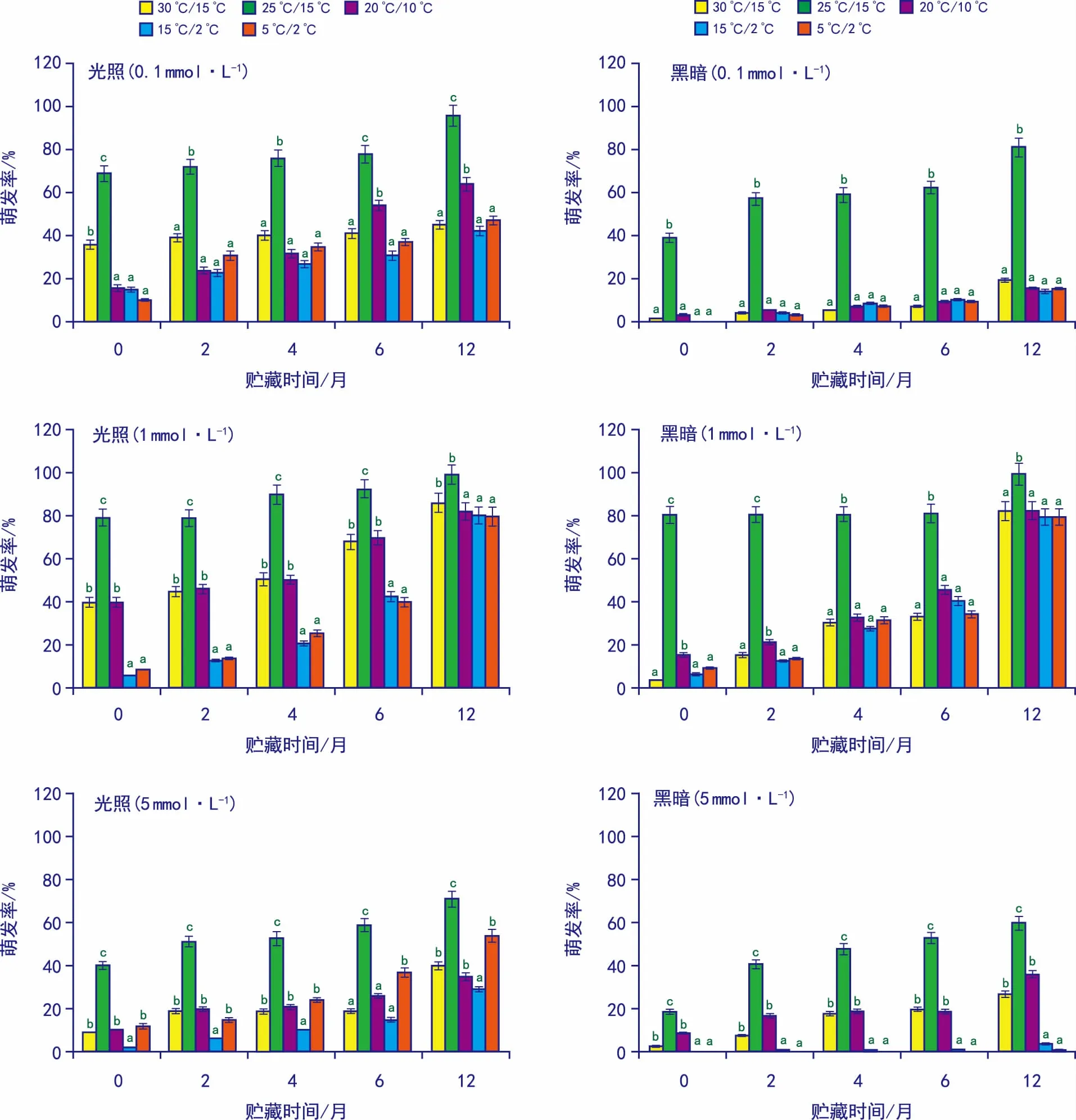
图2 赤霉素0.1、1、5 mmol·L-1处理光照/黑暗及全黑暗条件下种子萌发状况

图3 野外埋藏处理光照/黑暗及全黑暗条件下种子萌发状况
种子休眠可以在不适宜幼苗生长的情况下阻止种子的萌发,减少子代的风险,一定程度上保障了种群的延续[4,14]。紫苞鸢尾在新疆分布于气候多变的恶劣环境区域,该区域4月初气温迅速升高,在4月底至5月初受北部寒流影响,通常有大幅度降温或暴雪天气,恶劣气候影响该物种的开花结实率,对其幼苗产生毁灭性打击。试验结果显示,该物种与鸢尾属其它物种相比具有较高的萌发率,其种子随着室内干藏及野外埋藏时间的延长种子萌发率均呈现上升的态势(图3),这可能是该物种种子躲避山区的恶劣环境,形成土壤种子库,分散种子萌发的时间段,保证幼苗成活率,增加种群数量,实现该物种在新疆退化草地上种群扩张蔓延的适应性。
5 结 论
鸢尾属植物种子与同属其它物种相比,在光照/黑暗25 ℃/15 ℃条件下的萌发率较高,达82.2%,而其它温度条件下萌发率均低于35%;该物种种子存在休眠,属于非深度生理休眠;种子随着室内干藏时间延长,光照及全黑暗各变温条件下种子的萌发率均呈现不同程度的增加,种子可能存在后熟作用;用GA3处理紫苞鸢尾种子对打破其休眠有一定的作用,作用大小随着种子储藏时间的延长而增加;种子萌发率随着野外埋藏时间的增加呈上升趋势。这些休眠特性可能是由该物种种子长期适应分布区多变的气候条件,实现种群在新疆退化山地草原上扩张的一种适应。
猜你喜欢
沈阳农业大学学报(2022年2期)2022-06-04
体育科技文献通报(2022年3期)2022-05-23
种子(2022年1期)2022-02-24
中老年保健(2021年7期)2021-08-22
发电技术(2020年3期)2020-06-29
天天爱科学(2019年5期)2019-09-10
热带农业科学(2018年2期)2018-06-21
农村百事通(2018年24期)2018-02-22
江苏农业科学(2016年1期)2017-05-17
农村农业农民·B版(2017年2期)2017-03-11
