
松弛素及其相关肽的构效关系和生物学功能研究进展
2020-07-09 03:10李纯锦张博琦
黑龙江动物繁殖 2020年4期
陈 通,李纯锦,赵 云,张博琦,王 楠,周 虚
(吉林大学 动物科学学院动物繁殖调控实验室,长春 130062)
1926年F.L.Hisaw[1]发现,将妊娠豚鼠或者家兔血清注射到初情期豚鼠体内,血清中存在的某种因子会导致初情期豚鼠耻骨韧带松弛,这种因子被命名为松弛素(relaxin)。次年在猪黄体和家兔胎盘中发现相同功能因子。1930年首次从猪黄体中分离提纯得到松弛素。研究者随后在其他动物组织也发现了这种松弛因子及其类似物。松弛素及其相关肽被命名为松弛素肽家族。1945年后关于松弛素生理功能的研究成为热点,发现了松弛素在动物生殖和生殖以外的生物学作用。1980年开始利用DNA重组克隆技术探究松弛素肽家族的分子生物学结构和特性。2000年之后随着新技术的出现和发展,研究者发现了松弛素肽家族的受体RXFP1-4[2],并对松弛素与受体间的构效关系进行了研究。松弛素及其类似物已经在临床和畜牧生产中成为一些疾病的治疗药物,并且有望在未来作为某些疾病的预测、评估指标,对新药物开发和利用具有重要意义。
1 分子结构特性
研究表明松弛素肽家族可能是从胰岛素样肽中分化出来的[3],松弛素肽家族包括两个亚家族a和b,见图1。a家族包括胰岛素样肽3(INSL3)、胰岛素样肽4(INSL4)、胰岛素样肽6(INSL6)、人松弛素1(H1)和松弛素[即人松弛素2(H2)],非灵长类动物体内松弛素等价于人松弛素2。b家族包括胰岛素样肽5(INSL5)和人松弛素3(H3)。这表明在肽链结构和基因的角度上松弛素与胰岛素是具有同源性的。
1.1 松弛素的分子结构
松弛素属于水溶性多肽类激素,由α(也称A)及β(B)两个多肽链组成,其中α链存在链内二硫键,两条链之间以两个二硫键相连结,分子质量为6 000~23 000 Da。松弛素前体是一种包含信号肽、β链、连接肽和α链四种结构的多肽链[4],前体经历信号肽S和链接肽C的去除得到成熟松弛素(见图2)。
1.2 松弛素相关肽的分子结构与生物学特性
松弛素相关肽的基本结构与松弛素相似,成熟的肽都由α和β两条链构成。但对比这些多肽和松弛素的初级序列和分子结构发现,松弛素和这些多肽之间除了形成三个二硫键的半胱氨酸(cys)和肽的整体折叠相似外,几乎没有序列同源性[5]。松弛素肽家族之间分子结构的差异可能是由于连接肽C的氨基酸残基组成和转化酶识别位点存在物种特异性,导致松弛素在修饰过程中剪切位点不同造成的。
2 来源与分布
2.1 松弛素的来源与分布
早期的研究发现,松弛素主要存在于妊娠期哺乳动物的生殖相关组织中[3],黄体和胎盘是哺乳动物体内松弛素的主要来源(见表1)。
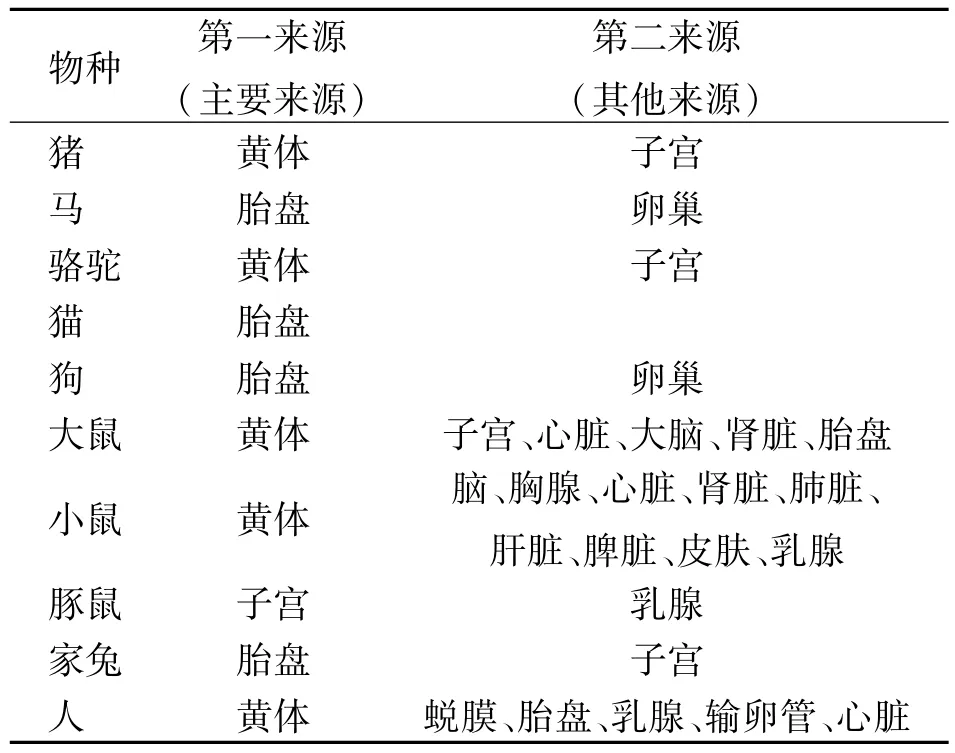
表1 常见哺乳动物中已知的松弛素来源
值得注意的是在反刍动物中松弛素是否存在以及来源问题仍然存在争议,在牛和绵羊基因组联合分析中,编码卵巢松弛素的基因缺失,但外源松弛素仍然可以在其体内发挥与其他哺乳动物相似的功能[6-7]。
此外,在哺乳动物生殖以外的组织中,如猪的垂体[8],大鼠的脑、心脏、肾脏[9],人的心脏[10]等处也检测到松弛素的表达。
2.2 松弛素相关肽的来源与分布
松弛素相关肽的来源也基本得到了验证,其中表达H1的基因只在高等灵长类动物中被发现[11]。H3在人脑和睾丸组织中大量表达[12],H3被认为是松弛素的祖先肽。
人胎盘、胎儿睾丸间质细胞和哺乳动物的睾丸间质细胞中都发现了INSL3的表达[13-14]。值得注意的是INSL3在反刍动物奶牛[15]和绵羊[16]卵巢和黄体中的表达与其他哺乳动物中松弛素的表达模式相似,推测可能在反刍动物中INSL3具有等同于松弛素的生物学功能。INSL4在人胎盘和胚胎组织中表达,可能与胎儿的组织发育有关。INSL5在小鼠和人的结肠中高表达,在肾脏、心脏和卵巢中也有表达。INSL6在人和大鼠睾丸组织中的表达最先被验证,最近的一项研究表明,在小鼠骨骼肌中也有INSL6的表达[17]。
总之,松弛素在哺乳动物组织中广泛表达,且在动物体内进行全身性循环,而其他多肽的表达和分布则具有物种差异性。反刍动物体内INSL3可在某种程度上替代松弛素。
3 松弛素与受体间的构效关系
3.1 松弛素及其相关肽的受体
松弛素及其相关肽的受体RXFP1-4属于G蛋白偶联受体家族。其中松弛素受体RXFP1和INSL3受体RXFP2属于富含亮氨酸重复序列(LRR)亚家族(见图3),在胞外区域存在一个包含LRR结构域和低密度脂蛋白(LDLa)模块的氨基末端,其中LRR结构域负责与松弛素高效率结合,LDLa则与受体的信号转导和反应易位有关[2]。H3受体RXFP3和INSL5受体RXFP4则不包含胞外的长氨基末端,其与受体结合并且被激活的确切机制尚不明确。
3.2 松弛素与受体的构效关系
松弛素是一种多效性激素,松弛素与受体之间的结构效应是松弛素发挥功能的重要基础。利用核磁共振技术对不同物种松弛素的氨基酸序列进行比对,结果表明,在松弛素β链中间区域存在三个相似的残基R-B13、R-B17和I-B20[5]。这三个功能残基构成“松弛素结合盒子”结构域,其中第三个残基I-B20可以是缬氨酸(V)或者异亮氨酸(I)[18]。松弛素中这一区域主要与受体RXFP1上细胞外一段富含亮氨酸的重复序列(LRR)相互作用[19]。最新的研究表明,在α链中同样存在Y-A13、L-A20和F-A23三个残基,对松弛素与受体的结合起重要作用[20]。L.M.Haugaard-Kedstrom等[21]将α和β链的C末端酰胺化后,发现松弛素依然具有生物活性。这表明自由C末端对于松弛素活性没有影响。
3.3 松弛素相关肽与受体的构效关系
H3作为一种神经肽,主要通过RXFP3受体发挥生物学作用。与松弛素一样,H3肽链中具有与受体相互作用相关的氨基酸残基(R-B8、RB12、I-B15、R-B16和F-B20),与松弛素不同的是H3的β链自由C末端对H3本身的活性具有重要作用[22-23]。
INSL3主要在间质细胞中表达,通过与受体RXFP2相互作用发挥生物学功能,INSL3的两条肽链中均含有与受体结合相关的氨基酸残基,并且在α链中存在对RXFP2受体的激活至关重要的氨基酸残基(R-A8、Y-A9)[24]。INSL5是一种在肠道中高度表达的新型肽激素,通过RXFP4受体发挥生物学作用,其肽链上同样具有与受体结合相关的氨基酸残基。与其他多肽不同的是,INSL5的β链末端存在刚性α-螺旋,它可与受体结合,具有很强的特异性[25]。这可能与刚性螺旋的存在有很大关系,并且β链的自由C末端对其生物活性具有重要影响[26]。
4 生物学功能
松弛素在非妊娠的哺乳动物中以自分泌或旁分泌的方式发挥作用,在妊娠的雌性动物中则可以通过内分泌的方式进行全身性循环[27]。松弛素在不同物种中来源不同,且在不同物种中同源性较低,因此松弛素的某些生理功能只存在于特定物种中。
4.1 松弛素在生殖方面的作用
4.1.1 耻骨间韧带 松弛素被敲除的小鼠耻骨间韧带发育失败[28],表明松弛素对于小鼠耻骨间韧带发育至关重要。松弛素会通过促进胶原蛋白分解和抑制胶原蛋白合成提高耻骨间韧带的转化能力。在许多物种中(如豚鼠、小鼠、蝙蝠和人类),分娩时都会伴随着骨盆韧带的改变(柔软,有弹性)[29],但这种现象在大鼠和绵羊中不会出现。松弛素的这种使耻骨间韧带软化的作用表明,松弛素在临床中可能成为辅助治疗初产或者难产患者的有效手段。
4.1.2 子宫颈和阴道 松弛素也会促进妊娠期雌性动物生殖道生长和软化,如在大鼠中松弛素可以抑制细胞凋亡,促进上皮细胞和基质细胞积累,刺激细胞增殖,导致子宫颈和阴道延展性发生变化[30]。这表明松弛素作用于生殖道可以增加延展性,有助于分娩时胎儿的排出。
4.1.3 子宫 在小鼠、大鼠、猪、豚鼠、仓鼠[31]等哺乳动物体内注射松弛素可以显著降低子宫肌层收缩频率和幅度;雌激素和孕酮刺激会增加大鼠和猪子宫肌层对松弛素的敏感性[32]。研究表明,松弛素可能通过与雌激素、孕酮的协同作用降低妊娠动物子宫的自发性收缩。
4.1.4 乳腺 松弛素在雌性动物乳腺正常发育中至关重要,通过注射特异性抗体抑制了大鼠内源性松弛素之后,大鼠乳腺发育受阻[33]。松弛素可能在乳腺发育中起促进乳腺组织生长的作用。
松弛素在雄性生殖中也至关重要。最新的研究表明,松弛素通过减少细胞凋亡和增加线粒体活性、过度激活cAMP水平以及顶体反应来影响体外人类精子的各项指标[34]。
4.2 松弛素在生殖以外的作用
研究人员在动物体内越来越多的组织中检测到松弛素基因的表达。越来越多的研究表明,松弛素除了在生殖功能上有重要作用外,在非生殖方面也有重要作用,如抗纤维化、伤口愈合、心脏和脉管系统保护等方面。
4.2.1 抗纤维化 松弛素可以通过减少Ⅰ型和Ⅲ型胶原蛋白合成从而抗纤维化,从而抵抗由转化生长因子-β诱导的人真皮成纤维细胞、肺脏成纤维细胞和心脏成纤维细胞的纤维化[35]。
4.2.2 伤口愈合和损伤修复 伤口愈合是一个复杂的生理过程,其中包括了炎症、颗粒细胞组织形成、再上皮化和组织重塑等一系列需要充足血液供应的过程。松弛素可以减轻或者缓解脂多糖诱导的急性肺脏损伤和由缺血再灌注造成的急性肾脏损伤[36-37]。研究表明,松弛素可能通过增加在伤口组织周围供血来实现伤口愈合和损伤修复。
4.2.3 心脏和脉管系统保护 使用纯化的猪松弛素或者重组H2松弛素可以在心脏、肾脏和肝脏等组织器官中诱导血管扩张[38];重组人松弛素不仅可以增加血管内皮生长因子、基质金属蛋白酶的表达,还可以直接通过一氧化氮的作用松弛血管平滑肌,增加肾脏血浆流量[39]。
4.2.4 其他作用 松弛素可以抑制肥大细胞和嗜碱性粒细胞的颗粒胞吐和组胺释放[40],延缓大鼠由衰老引起的炎症反应[41]。
4.3 松弛素相关肽生物学功能
与松弛素相比,松弛素家族其他多肽类的生物学功能有待考证,现就已知的生物学功能进行阐述。
INSL3在雌性动物中被证实对腔前卵泡生长和激素的分泌具有重要作用。INSL3可以通过抑制细胞凋亡减缓小鼠卵巢卵泡闭锁和黄体溶解[42];卵泡膜细胞分泌的INSL-3以自分泌或旁分泌的方式作用于卵泡膜细胞或颗粒细胞上的RXFP2受体,刺激其产生17α-羟化酶[43],该酶负责从孕酮到雄烯二酮(类固醇激素的前体)的转化;INSL-3也会促进有腔卵泡产生雌二醇[3]。此外,INSL3在雄性生殖方面同样具有重要作用。雄性小鼠敲除INSL-3后出现隐睾[44],表明INSL-3与睾丸沉降有关。这说明INSL3可能与动物的一系列生殖疾病有关,如雌性动物的多囊卵巢综合征、卵巢早衰、雄性动物的睾丸发育失败等,推测INSL3可用于调节卵泡发育,或者预防雄性动物睾丸发育失败的激素治疗方案。
H3在动物大脑中高表达,在应激反应的认知信号具有重要作用,在治疗认知、情感和情绪障碍或神经系统疾病方面具有很好的应用前景。INSL4在动物胎盘和胎儿组织中表达,可能与胎儿的组织器官发育以及骨骼生长有关;近期的一项研究表明,INSL4异常表达可能会支持肺癌细胞的生长[45]。INSL5主要在动物结肠中表达,可能与能量代谢有关。最新的研究表明,INSL5可能在调节胰岛素分泌和胰岛β细胞稳态中起着重要作用;并且INSL5敲除雄性和雌性小鼠分别表现出精子活力下降和发情周期不规则[46]。INSL6在心脏应激的小鼠动物模型中表现出缓解心脏收缩障碍和抑制心肌纤维化的生理功能[47],因此INSL6在未来心脏疾病中可以作为一种新型药物缓解心肌纤维化和心脏收缩障碍。
5 结语
松弛素作为一种血液循环的激素,其受体在动物体内多数组织中都有表达,不仅在生殖方面具有重要功能,在生殖以外如减缓炎症、抗纤维化和脉管系统保护等方面也具有重要作用。近几年来基因编辑技术的成熟将给松弛素功能研究带来更加便利和可信的技术手段,对于松弛素及其受体的结构效应研究(如生产分子结构更简单、生物效率更高更稳定的类似物肽等)还有广阔的前景。
猜你喜欢
生物化学与生物物理进展(2022年7期)2022-07-25
生物化学与生物物理进展(2022年6期)2022-07-21
中国典型病例大全(2022年11期)2022-05-13
西南农业学报(2021年10期)2021-12-14
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19
中国生殖健康(2020年4期)2021-01-18
故事作文·高年级(2019年10期)2019-10-16
北方农业学报(2016年6期)2016-12-16
中学生数理化·七年级数学北师大版(2008年6期)2008-10-22
