
论植物养分获取的杂种优势
2020-07-06 09:00潘春桂
陕西农业科学 2020年5期
潘春桂
(安徽省东至县种子管理站,安徽 东至 247210)
从达尔文时代人类就已经认识到杂种优势现象,利用杂种优势增加经济效益以及研究杂种优势的形成机理也有近一个世纪。对其机理的解释,经典遗传学提出了显性、超显性、上位性效应假说,分子遗传学认为基因差异表达、表观遗传修饰包括DNA甲基化和组蛋白修饰参与杂种优势的形成[1],以上假说也获得了相关试验的支持。另外,一些学者也提出了遗传平衡、活性基因、遗传振动活成、基因网络系统及自组织理论等假说[2],不难看出,以上假说共同性在于将杂种一代内在相关过程的特异性等价为杂种优势的形成机理或产生原因,实际上它们仍然为杂种优势的表现或结果,就像杂种一代营养体较大,生物或经济产量较高一样,不能作为杂种优势的形成原因。可能这些研究潜在地认为亲本两套遗传物质相互作用就产生了杂种优势,排斥或弱化了环境因素在杂种优势形成中的地位。相同遗传类型在不同环境中具有生理、生化、形态过程的差异性,表明环境效应从遗传表达到形态建立各个环节中均产生重要影响。从能量平衡而言,杂种一代的优势应该与获得更多的外部环境养分有关,植株体获取养分的遗传过程如何随着外部环境而发生变化,笔者从一些基本假设出发,论述养分获取的杂种优势。
1 假设与验证
植株体对外部环境中养分吸收速度与遗传、环境养分浓度及植株体大小等有关,可用Logistic曲线来模拟植株体生长过程[3,4]。可近似认为,在一定条件之下,①植株体对各种营养元素的吸收存在饱和浓度或饱和期,此时增加各种养分的浓度或环境供给,植株体对它们的吸收速度不再增加[5],这里假定不考虑盐害浓度。②在饱和期杂种一代对各种元素的吸收速度为加性遗传。③植株体对各种元素吸收速度为固定比例,只与遗传类型有关。④各种营养元素均存在临界环境浓度或临界点,对某种元素而言,此时减少其环境浓度或供给,植株体对这种元素的吸收速度会下降,其它元素也协同下降,但吸收速度仍保持固定比例。基于以上假设,如果一些元素饱和期吸收速度在双亲间存在差异,则最大元素吸收速度被双亲所具有,因此,经过一段生长时间后,双亲均因为各自具有最大元素吸收速度比杂种一代可能更早地通过临界点,其后杂种一代对各种元素吸收速度便超过中亲值,因此,各器官或单株重的杂种优势便开始形成。

表1 不同元素吸收速度一种变化模型
表1假设ti+1-ti= △t(i =0.1.2),a1>a2,b2>b1,父本在t1时刻元素A穿过临界点,母本在t2时刻B元素穿过临界点,其后均线性下降;杂种一代一直处于饱和期。v<1,u<1,从t0时刻开始统计,可算得t3时刻两种元素的吸收量。
父本:(2+v)×(a1+b1)△t
母本:(2.5+0.5u)×(a2+b2)△t
杂种一代:0.5×3×(a1+b1)△t+0.5×3(a2+b2)△t
根据上文假设条件,易得2F1/(p1+p2)>1,即此时段已形成正向中亲优势或杂种优势。当然,对于某一种元素而言,不同生长时期饱和吸收速度存在差异,临界环境临度浓度也不同,以上分析假设饱和吸收速度不变。上文养分获取的杂种优势形成机制可产生一些一般性关系。下面通过一个模拟试验的结果进行分析。为了简化计算过程,假设各种养分吸收过程可分为二个阶段:固定速度吸收期(饱和期)及停止吸收期(中止期),当某一种养分环境消耗率达到了一定比例时,便从固定速度吸收期转为中止期,转折点称为临界点(时刻)。这里认为各种养分临界环境消耗率相同,杂种一代为加性遗传;饱和期各种养分吸收速度也为加性遗传。设置的6个亲本P1~P6,尽量代表不同的遗传类型。P1~P4在饱和期有较大的养分吸收速度,P5饱和期养分吸收速度很小,P6养分临界消耗率较低。
表2中6个亲本配置半完全双列杂交15个组合。先简单介绍试验数据的取得方法,首先设环境中三种养分A、B、C总供给(vt)分别为5、4、3,当任一种养分临界消耗率为80%时,三种养分临界消耗量(vt)分别为4、3.2、2.4,其它临界消耗率时类推。对于P1,三种养分A、B、C的吸收速度(v)分别为1.3、2、1.5,临界消耗率为80%,经过时间1.6(t)时,A、B、C三种养分消耗量(vt)分别为2.08、3.2、2.4,可看出B、C两种养分已达到临界消耗量,1.6(t)就是临界点(时刻),这时个体停止对各种养分的吸收,即由饱和期进入中止期,养分吸收总量停止在2.08+3.2+2.4=7.68(vt)上。对于P2类似计算,经过1.6(t)时,养分C达到临界消耗量,总吸收量停止在(1.8+1.5+1.5)×1.6=7.68(vt),当然在临界点之前养分吸收总量为三种养分吸收速度之和与时间乘积。P1×P2三种养分A、B、C吸收速度(v)分别为1.55、1.75、1.5,临界养分消耗率为80%,在1.6(t)时,养分C达到临界点,总吸收量停止在(1.55+1.75+1.5)×1.6=7.68(vt)。按相同方法计算出所有亲本和组合各时期个体养分吸收总量,经统计可得出以下结果:
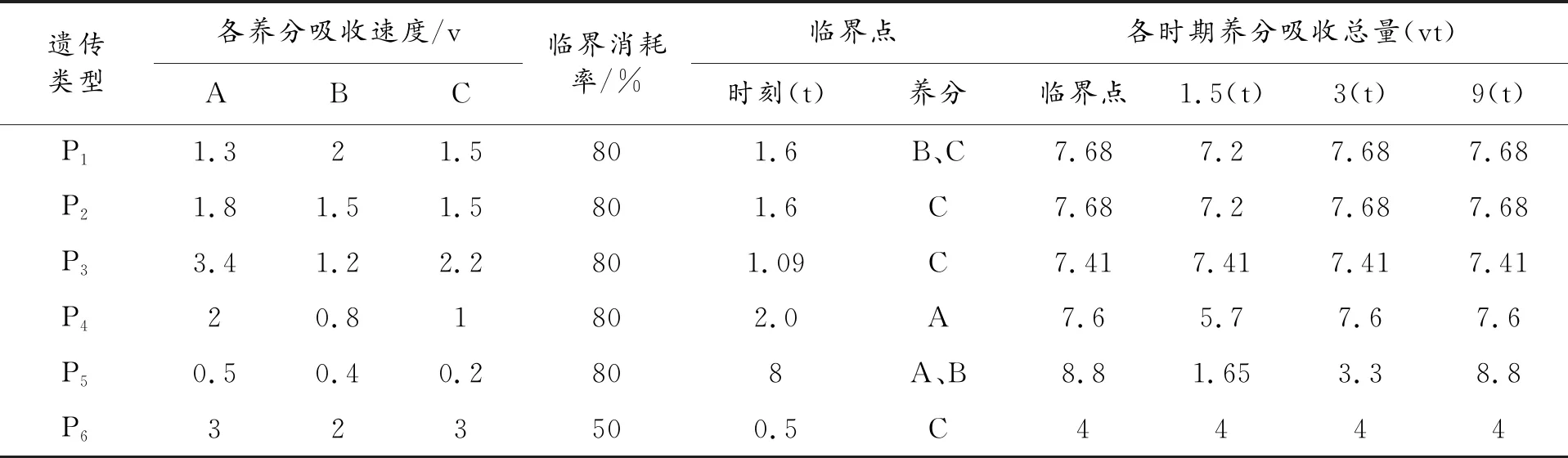
表2 各种养分吸收速度及个体养分总吸收量变化过程
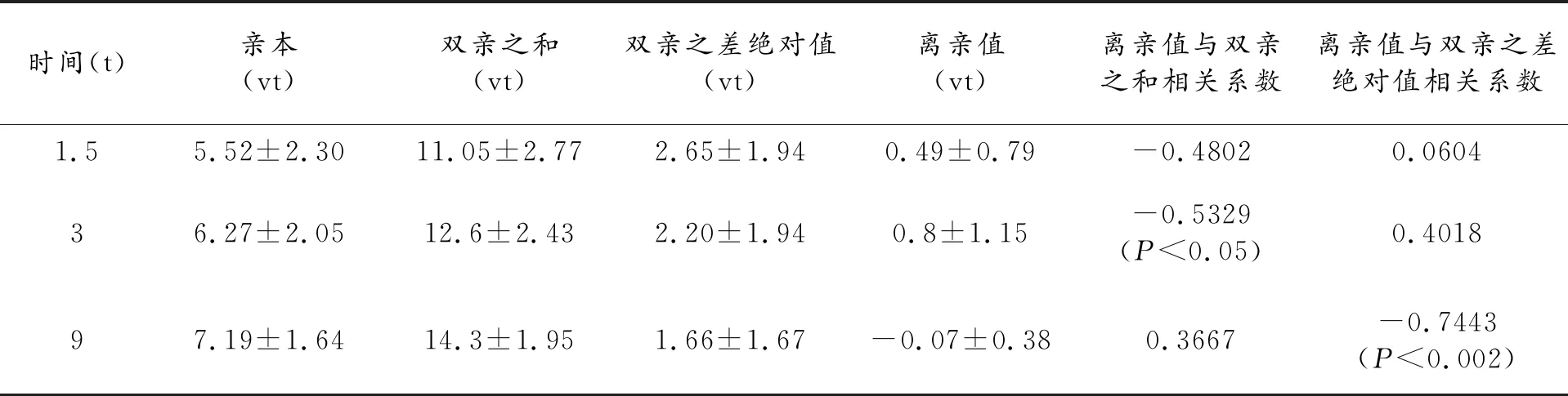
表3 个体养分吸收总量及相关统计
离亲值指杂种一代值与中亲值(双亲平均值)之差,这里用以表示杂种优势的大小。另外,值得注意的是与个体总养分吸收量有关的另外几个统计量,1.5(t)时双亲之和、双亲之差绝对值与3(t)时离亲值相关系数分别为-0.6979(P<0.005)及0.7270(P<0.005),1.5(t)到3(t)时,增加量离亲值与1.5(t)时、3(t)时双亲之差绝对值相关系数分别为0.7015(P<0.005)及0.5199(P<0.05)。可见生长(取样)时期对统计结果有明显影响。
从以上表一养分吸收模型和模拟试验可产生如下推论:
(1)在养分供应充足和植株生长很早的时期,杂种优势活跃度很低,即平均值很小或趋于零,变化也较小。因为这个时期可能很短等原因以致实际情型中不易被分辨出来。
(2)如果植株生长时间足够长或各种养分已进入较显著的环境胁迫期(短缺期),这时杂种优势表现活跃。植株体与环境相容性好易获得较大的重量(干重或鲜重,下文同)。相容性包括各种养分饱和吸收速度、各种养分进入临界点时间同步性、养分临界消耗率等方面。相容性好指这几方面均表现较好。以上几方面其中一种或多种同时表现较差时则为环境相容性差,使植株体重量变小。由于以外部环境作共同参照,显然,双亲重量越小,双亲遗传差异总体上较大,但在不同组合间可有一定变化;双亲重量越大,则遗传相似性越好;环境相容性好的双亲各种营养元素吸收速度的衰减(相对于饱和吸收速度衰减,下文同)和杂种一代相近,衰减时间也和杂种一代较同步,因此不易形成较大的杂种优势;环境相容性差且遗传差异较大时,可导致双亲间各种养分平均衰减幅度更大,平均衰减时间更早,易形成较大的杂种优势。因此在大多数情况下,双亲重量之和或中亲值与杂种优势或离亲值呈显著的负相关。上文模拟试验可见到这种结果。
(3)也可以呈现其它结果,如植株重量双亲之差绝对值与杂种优势正相关,甚至双亲之和与杂种优势正相关,这与试验条件及取样时期有关。下文通过文献数据的计算结果来证明以上推论。马谦的试验数据表明[6],大田条件下水稻播种后第20天的幼苗干重杂种优势为-5.8±19.4(%),而第30天为23.41±19.1(%),第20天到第30天幼苗干重增加量杂种优势与第20天双亲干重差绝对值呈正相关,相关系数为0.3799(P<0.05),与第30天双亲干重差绝对值弱正相关,相关系数为0.2699,这与上文模拟试验结果也相似;与第30天干重双亲之和呈负相关,相关系数为-0.5272(P<0.005)。以上结果表明,第20天时几乎所有亲本仍处于饱和期或在临界点附近或者说各种养分供给很充分。第30天时,所有亲本均已通过临界期。余丽霞棉花杂交种愈伤组织培养试验数据计算表明[7],第35天到第65天愈伤组织重量增加量杂种优势与第35天、第65天、第80天双亲重量之和相关系数分别为-0.5082(p<0.01)、-0.7141(p<0.001)、-0.7089(p<0.001)。另外,亲本愈伤组织重量(g)在第35天、第65天时、第80天时分别为1.32±0.51、2.13±0.88、2.75±1.16,亲本间差异性变化趋势与上文模拟试验相反,与这可能与试验材料及取样时间等有关;但以上结果也表明,愈伤组织重量亲本间变化较大时,双亲之和与杂种优势相关性较强,这和模拟试验是一致的。第65天到第80天愈伤组织重量增加量杂种优势与第35天、第65天、第80天双亲重量之和相关系数分别为-0.5442(p<0.005)、-0.5331(p<0.01)、-0.5903(p<0.002),可能生长后期外界养分已较严重亏缺,杂种一代对各种养分吸收速度也衰减严重,杂种优势活跃度下降,双亲遗传差异大小与愈伤组织增加量杂种优势相关性减弱。王章奎营养液栽培小麦试验表明[8],拔节期单株根系干重杂种优势与双亲之和相关系数为-0.7505(P<0.001),离亲值与中亲值相关系数为-0.7049(P<0.001),说明这时营养液各种养分的供给已进入较重的胁迫阶段。温玲甜瓜杂交种试验表明[9],4叶一心期单株根系干重杂种优势与双亲之和相关系数为0.7435(P<0.05),离亲值与中亲值相关系数为0.7894(p<0.02);这与上文推论③有关。大田作物经过一个生长季节,土壤养分已被较多地消耗,这时作物产量杂种优势与双亲之和易为显著或极显著负相关。另外,上文已经提及,双亲之和越小时,双亲遗传差异在不同组合间可变化较大。因此,从线性相关的计算原理可知,由于亲本选配的随意性,有些情况下去除双亲之和最小的一些组合,可显著提高杂种一代离亲值或杂种优势与中亲值相关性。吴利民小麦杂交试验原有杂交组合20个[10],单株产量杂种一代离亲值与中亲值相关系数为-0.4639(p<0.05),现去除单株产量双亲之和最小的7个组合,重新计算出以上相关系数为-0.7266(p<0.01)。桑世飞油菜杂交试验原有组合46个[11],去除产量双亲之和最小的6个组合,杂种一代产量离亲值与中亲值相关系数由原-0.4548(p<0.002)变为-0.5643(p<0.0005)。张锡顺蓖麻杂交试验原有杂交组合30个[12],现去除单株生产力双亲之和最小的13个组合,单株生产力杂种一代离亲值与中亲值相关系数由原-0.4420(p<0.02)变为-0.7953(p<0.001)。在上文模拟试验中,在1.5(t)时去除个体养分吸总量双亲之和最小的9个组合,也即P5和P6参与的所有组合,1.5(t)时双亲之和与3(t)时离亲值相关系数由原-0.6979(p<0.005)变为-0.9681(p<0.002)。由于植株体主要由大量元素构成,但它对大量元素和少(微)量元素的获取是相互协同和相互制约的关系,因试验环境、遗传背景、目标性状及取样时间不同,杂种优势与亲本生物体重量之和的相关性存在可解释的变化,但在生长后期或环境养分相对亏缺时,负相关性几乎是必然的,有兴趣的读者可以去验证。
2 讨论
单株截获光能等于单株叶面积和单位叶面积截获光能的乘积。一般而言,由于叶层相互遮掩,单株叶面积与单位叶面积截获光能呈反向变化关系,也即单株截获光能具有自然或本能的反极因子效应,或者说其正向中亲优势即杂种优势是必然发生的[13]。再加上文章中养分获取的杂种优势形成机理,使植物体整体或部分重量存在正向中亲优势或杂种优势从偶然性转变为近似于一种本能或必然的自然现象。这从公开发表的各种作物杂交试验的有关文献中能够发现,几乎所有杂交试验中,各组合中亲优势平均值大于零或者正向中亲优势的组合比例大于50%,占很大比例的试验中均接近100%。上文表1基本假设和模拟试验强调了环境变化的作用,只假设有关性状为加性遗传,即可发现杂种优势或超亲优势广泛存在,也得到了引文试验数据统计结果的验证,这值得重视。很显然,环境相容性包括养分的供给数量、空间分布及供给时期等方面,也包括与养分供给无关的其它外界因素如温度、光照、病害等。养分获取的杂种优势不完全等同于经济产量的杂种优势,有必要深入地研究。
3 结论
养分获取具有天然或本能的杂种优势,杂种优势是遗传与环境相互作用下动态形成的,应重视环境效应在杂种优势研究中的地位。当各种养分进入环境胁迫阶段时,杂种优势才可能产生;植株体重量主要由其环境相容性所决定。随着环境胁迫不断加重,植株体整体或部分重量的杂种优势与双亲之和呈显著或极显著负相关。
猜你喜欢
中国饲料(2022年5期)2022-04-26
蔬菜(2021年7期)2021-11-27
东坡赤壁诗词(2020年5期)2020-11-06
散文诗世界(2019年10期)2019-09-10
农业科技与装备(2017年9期)2018-03-02
小学生时代·综合版(2016年4期)2016-11-19
家庭医药·快乐养生(2016年9期)2016-09-08
江苏农业科学(2015年1期)2015-04-17
发明与创新·大科技(2009年5期)2009-05-31
青年文摘·上半月(1994年7期)1994-01-01
