
盐、干旱胁迫对拟南芥WRKY71基因突变体种子萌发的影响
2020-07-04 03:06徐金鹏祁亚男于延冲
山东农业科学 2020年3期
徐金鹏 祁亚男 于延冲
摘要:WRKY71是影响拟南芥开花和分枝发育的重要转录因子,而其在种子萌发中的作用尚不清楚。本研究以拟南芥野生型(Col-0)、WRKY71过表达突变体(D27)和T-DNA插入突变体(wrky71)种子为材料,分析正常条件、盐胁迫、干旱胁迫处理对种子萌发的影响。结果表明:正常条件下三种材料萌发一致;盐胁迫下尤其是LiCl处理的wrky71种子萌发率优于Col-0,而D27则低于Col-0;干旱胁迫1 d时,D27明显低于Col-0,随后三者的萌发率达到一致。这说明,D27种子萌发对盐和干旱胁迫较Col-0敏感,而wrky71相对不敏感。
关键词:拟南芥;WRKY71;盐;干旱;突变体;种子萌发
中图分类号:S567.210.1文献标识号:A文章编号:1001-4942(2020)03-0034-04
Abstract WRKY71 is an important transcription factor which regulates the process of flowering and branch development of Arabidopsis, however, its function on seed germination is not clear. In this study, Arabidopsis wild type(Col-0), WRKY71 overexpression mutant(D27) and T-DNA insertion mutant(wrky71) were used to analyze the effects of normal conditions, salt stress and drought stress on seed germination. The results showed that the germination rates of the three materials were the same under normal condition. Under salt stress, especially under LiCl treatment, the germination rate of wrky71 was higher than that of Col-0, while that of D27 was lower compared with Col-0. The germination rate of D27 was obviously lower than that of Col-0 after one-day drought stress, then that of different materials was similar. The results indicated that the seed germination of D27 was more sensitive to salt stress and drought stress compared with Col-0, while that of wrky71 was relatively insensitive.
Keywords Arabidopsis;WRKY71; Salt; Drought; Mutant;Seed germination
WRKY是近20年來发现的一类植物特异转录因子[1],因其N端含有近乎绝对保守的WRKYGQK氨基酸序列而得名[2]。大量研究表明WRKY转录因子家族主要在植物抗病和非生物胁迫应答中有重要作用[2,3],此外也参与到其它许多生理和发育进程,如叶片衰老、开花和分枝发育、表皮毛发育和糖代谢等[4-8]。
种子萌发是生长发育的起点,在植物整个生命周期中占有至关重要的地位[9,10],也是一个十分复杂的过程,受到许多内源和外源因素的联动影响。不同生境中的种子感知外界环境信号,启动内部激素信号,调控内部基因表达,进而控制种子在合适的时机萌发[11]。众多研究表明,盐和渗透胁迫会抑制或者延迟种子的萌发[12,13]。
许多研究表明,转录因子在种子萌发调控中起到重要作用,WRKY转录因子就是其中之一。大麦中,HvWRKY38会抑制GA诱导的α-淀粉酶的表达,进而抑制种子的萌发[14]。同样水稻中,OsWRKY71和OsWRKY51协同作用也可以抑制GA诱导的α-淀粉酶的表达,进而影响种子的萌发[15]。烟草中过表达小麦TaWRKY10可以增加烟草种子在NaCl和甘露醇处理下的萌发率[16]。ABA处理下拟南芥WRKY2突变体种子萌发率显著低于野生型[17];相反,拟南芥WRKY41突变体种子的萌发则对ABA不敏感,进一步研究发现,WRKY41可以直接调控ABA信号基因ABI3的表达进而影响种子萌发[18]。
本团队之前的研究表明,WRKY71可以与FT和LFY的启动子区域相结合,并上调其表达,从而加速拟南芥开花[8]。而且WRKY71过表达植株在高盐度条件下依然早花,表明WRKY71可以加速拟南芥完成整个生命史而逃避盐害[19]。然而WRKY71是否也可以加速拟南芥种子的萌发尚未可知。为此,本试验以拟南芥野生型(Col-0)、WRKY71过表达突变体(D27)和T-DNA插入突变体(wrky71)为材料,研究其在盐和干旱胁迫下的种子萌发情况,以期为研究WRKY71对拟南芥生活史的调控提供新内容,并为通过分子生物学手段影响作物种子的萌发提供参考。
1 材料与方法
1.1试验材料
哥伦比亚野生型(Col-0)拟南芥(Arabidopsis thaliana L.)种子为本实验室保存,WRKY71过表达突变体(D27)和T-DNA插入突变体wrky71为之前研究所用[19]。植物材料在光照培养箱(上海新苗医疗器械创造有限公司产品,型号GLX-3006S-III)中培养,培养条件:相对湿度40%~60%、温度22℃和16 h/d光照、8 h/d黑暗。
1.2 种子消毒
将新收获的(不超过30 d)拟南芥种子在无菌工作台上先用含0.03% triton 的75%酒精清洗1 min,再用70%酒精清洗两次,每次1 min,然后将种子置于无菌滤纸上晾干备用。
1.3 种子萌发与试验处理
将Col-0、D27和wrky71干种子撒于1/2MS培养基上作对照,另将三种种子分别播于65 mmol/L LiCl、250 mmol/L KCl、200 mmol/L NaCl和200 mmol/L甘露醇的1/2MS培养基上进行胁迫试验。每个平皿撒150粒,封口膜密封平皿,低温(4℃)黑暗环境下春化3 d,然后放到光照培养箱中进行萌发试验。
培养皿放置到光照培养箱后,每隔24 h进行一次萌发率统计(种子露出白色胚根即为成功萌发),共统计5 d。试验重复3次。萌发率(%)= 萌发种子数/培养皿中全部种子数×100。
1.4 数据处理
利用Microsoft Excel进行数据处理与作图。
2 结果与分析
2.1 盐胁迫对WRKY71突变体种子萌发率的影响
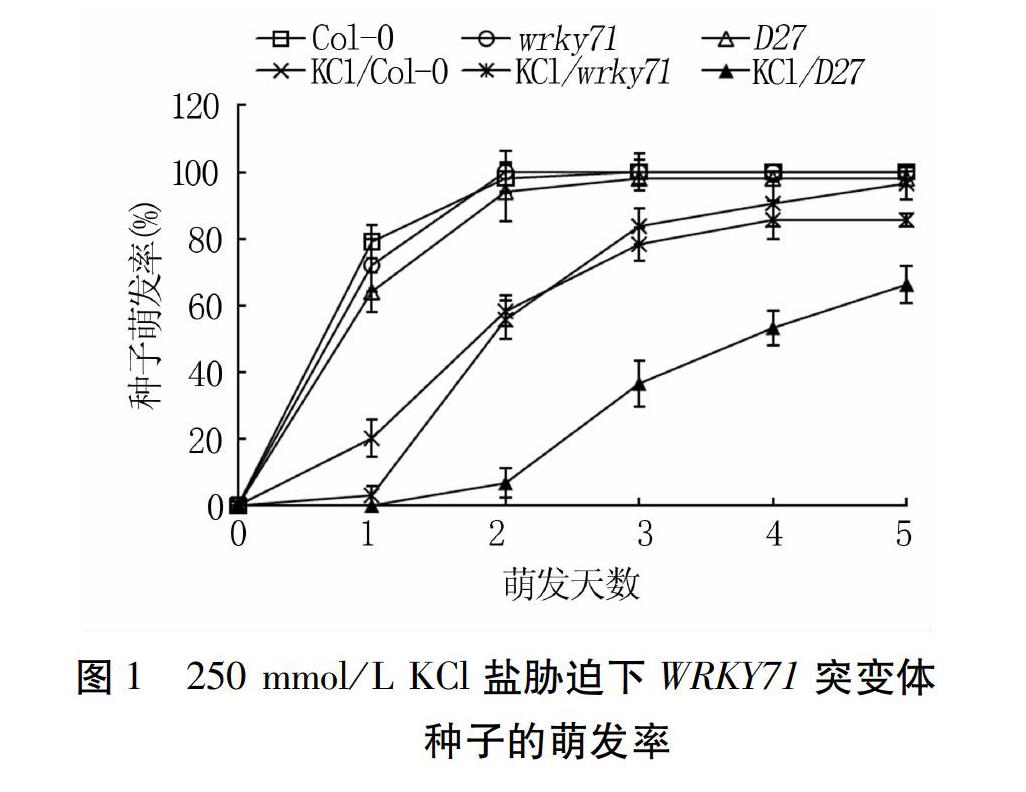
Col-0、wrky71和D27三种拟南芥种子对照的最终萌发率一致,均为100%,但wrky71第2 d达到最大萌发率,Col-0和D27为第3 d(图1)。250 mmol/L KCl胁迫下D27的萌发率受到明显抑制,第5 d时仅为66.2%,比对照降低31.8个百分点;Col-0受抑制程度较D27轻,第5 d时萌发率为85.6%,降低14.4个百分点;wrky71萌发尽管前期受到明显抑制,但第5 d时萌发率达965%,仅降低3.5个百分点(图1)。由此看出,D27种子萌发相比Col-0对KCl敏感,而wrky71相对最不敏感。
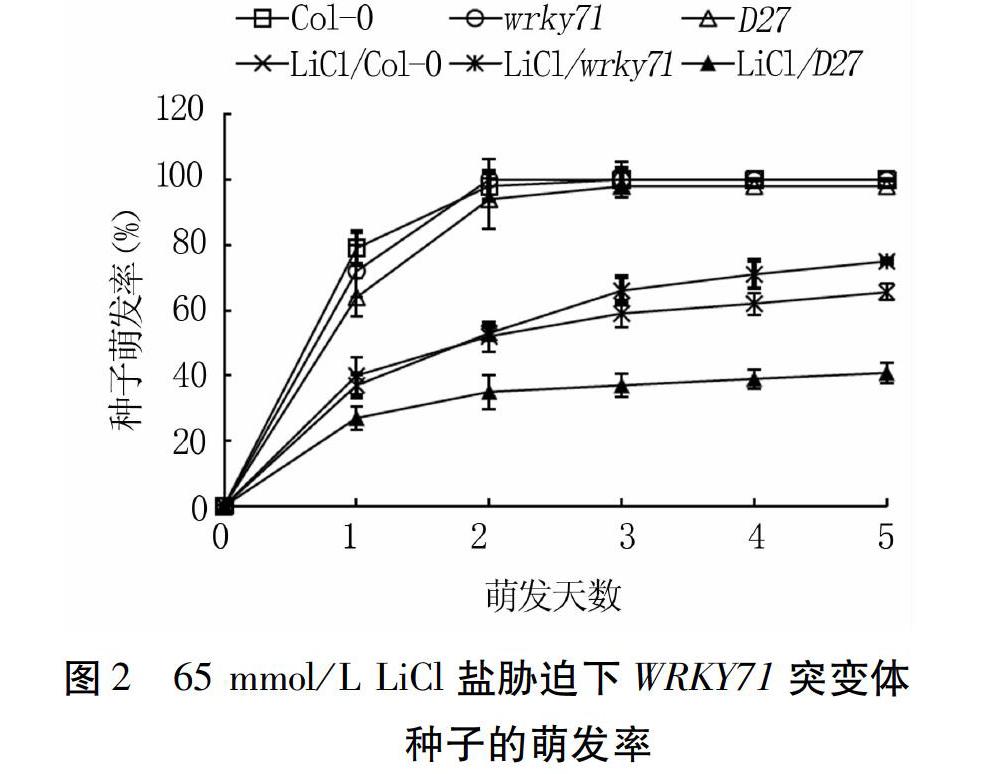
65 mmol/L LiCl处理下,Col-0、wrky71和D27种子萌发率受到明显抑制,其中5 d内D27的萌发率始终低于Col-0和wrky71,第5 d时其萌发率仅有40.8%,比对照降低57.2个百分点。试验前期(1 d和2 d),Col-0和wrky71的萌发率相当,第3 d始后者的萌发率显著高于前者,第5 d时二者的萌发率分别为65.5%和75.0%,比对照分别降低34.5、25.0个百分点(图2)。表明D27种子萌发相比于Col-0对LiCl敏感,而wrky71相对最不敏感。
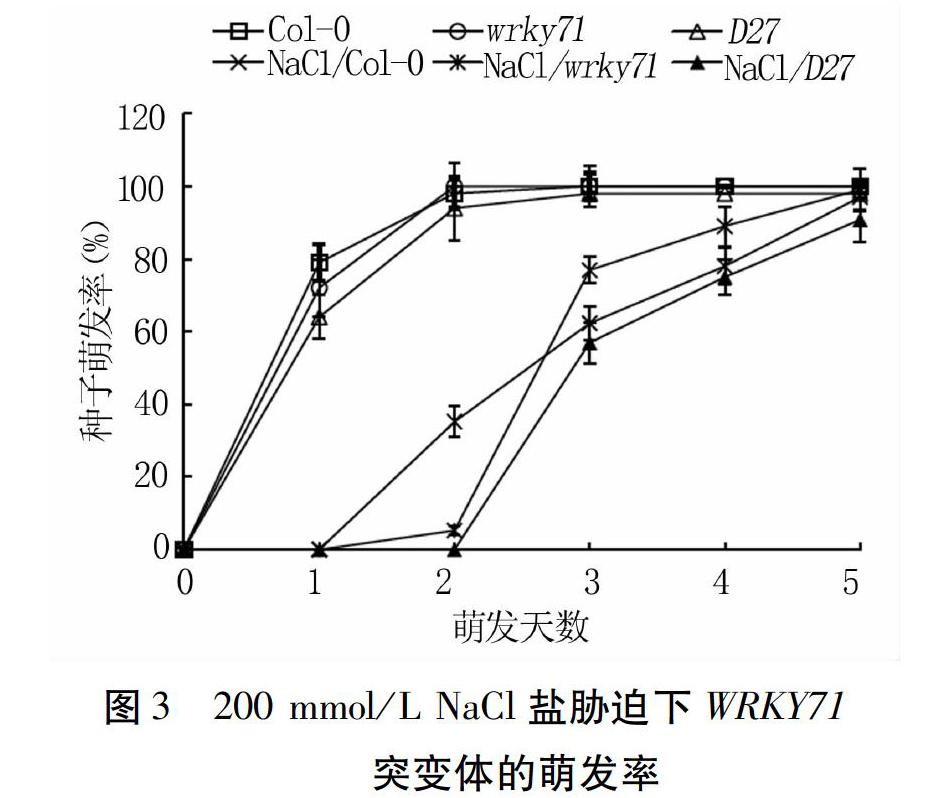
200 mmol/L NaCl处理下,Col-0、wrky71和D27种子萌发在试验前期(1~2 d)受到明显抑制,后期萌发率显著提升,第5 d时三者的萌发率分别为97.0%、99.0%和90.8%,分别减少3.0、1.0、7.2个百分点(图3),表明D27对盐最敏感。试验第2 d,Col-0萌发率高于wrky71,第3 d之后wrky71则显著高于Col-0,说明wrky71相对于野生型对NaCl适应较慢但耐受能力强。
综上所述,不同盐胁迫条件下,拟南芥三种种子的萌发时间和萌发数均受到一定程度的延迟和抑制,对照3 d时萌发率均达到100%,而盐处理达到最大萌发率时需要5 d。D27种子萌发受到三种盐胁迫的显著抑制,而盐胁迫下wrky71的萌发率相对于野生型有所延迟,但其最终耐受能力较高。
2.2 干旱胁迫对WRKY71突变体种子萌发率的影响
用含200 mmol/L甘露醇(Man)的1/2MS培养基进行干旱胁迫试验,结果表明,Col-0、wrky71和D27均在第1 d开始萌发,萌发率分别是51.3%、41.4%和14.3%,三者萌发率均受到一定影响,均低于对照,D27最明显。第2 d时,三种拟南芥种子的萌发率均达到100%(图4)。以上结果表明,在早期,D27种子萌发对干旱胁迫敏感性显著高于Col-0和wrky71。
3 讨论与结论
众所周知,植物生长发育过程中会遇到多种多样的生物与非生物因素影响,如土壤盐分和水分含量等,它们或有利于植物的生长发育或对其产生危害,甚至导致植物死亡。在长期的生物进化过程中,植物进化出一套相對完整的趋利避害机制,从而使植物在激烈的生存竞争中取得优势并扩大种群数量。在植物众多调控机制中,有一类被称为WRKY家族的调控因子,其许多成员已被证明是植物响应非生物胁迫的重要因子。WRKY71作为WRKY家族的重要一员,目前关于其在非生物胁迫下对种子萌发的影响未见报道。
前期研究表明,WRKY71的过表达会促进拟南芥开花、缩短生活史以帮助拟南芥逃避盐害。本研究发现,盐胁迫下尤其是LiCl胁迫下,Col-0、wrky71和D27三者的种子萌发均受到抑制,但D27受到的抑制更明显,wrky71受到的抑制最小(图1~图3)。因此推测,这一方面可能是因为WRKY71的过表达会使得植物细胞内活性表达上升、与生命活动相关的氧化酶活性增加,从而导致拟南芥能够提前开花,同时由于WRKY71的过表达使得细胞代谢活跃度增强,导致植物细胞可以更多地吸收外部环境中的离子,从而对D27种子造成更大的离子胁迫,其萌发受到显著抑制;另一方面,可能是因为D27属盐敏感型,相较于Col-0和wrky71表现出不抗盐的表型,因此盐胁迫下萌发最慢。到目前为止,WRKY71突变体种子内哪些因子参与了对盐胁迫的响应以及如何响应目前还不清楚,这也是今后进一步研究的目标。
干旱胁迫下D27种子早期萌发显著低于Col-0和wrky71,说明WRKY71的过表达使拟南芥种子萌发对干旱胁迫更为敏感,而WRKY71基因的敲除并没有使其种子萌发对干旱胁迫不敏感,推测可能与基因功能冗余有关,这与WRKY71在开花方面的功能冗余类似[8,19]。
本團队前期及本试验研究结果表明,WRKY71尽管能促进拟南芥开花以逃避盐害,但其在种子萌发中的作用则相反,其过表达会抑制种子的萌发。这也可能是一种逃避机制,但具体情况尚不清楚,需要进一步研究。此结果可为WRKY71转录因子的生物功能研究提供新思路,也可为通过分子生物学手段影响作物种子的萌发研究提供参考。
参考文献:
[1] Jiang J J,Ma S H,Ye N H,et al. WRKY transcription factors in plant responses to stresses[J]. Journal of integrative plant biology,2017,59 (2):86-101.
[2] Rushton P J,Somssich I E,Ringler P,et al. WRKY transcription factors (Review)[J]. Trends in Plant Science,2010,15(5):247-258.
[3] Chen L G,Song Y,Li S J. The role of WRKY transcription factors in plant abiotic stresses[J]. Biochimica et Biophysica Acta(BBA)-Gene Regulatory Mechanisms,2012, 1819(2):120-128.
[4] Johnson C S,Kolevski B,Smyth D R.TRANSPARENT TESTA GLABRA2,a trichome and seed coat development gene of Arabidopsis,encodes a WRKY transcription factor[J].The Plant Cell,2002,14(6):1359-1375.
[5] Sun C X,Palmqvist S,Olsson H,et al. A novel WRKY transcription factor,SUSIBA2,participates in sugar signaling in barley by binding to the sugar-responsive elements of the iso1 promoter[J]. The Plant Cell,2003,15:2076-2092.
[6] Jiang Y J,Liang G,Yang S Z. Arabidopsis WRKY57 functions as a node of convergence for jasmonic acid- and auxin-mediated signaling in jasmonic acid-induced leaf senescence[J]. The Plant Cell,2014,26(1):230-245.
[7] Guo D S,Zhang J Z,Wang X,et al. The WRKY transcription factor WRKY71/EXB1 controls shoot branching by transcriptionally regulating RAX genes in Arabidopsis[J]. The Plant Cell,2015,27(11):3112-3127.
[8] Yu Y C,Liu Z H,Wang L,et al. WRKY71 accelerates flowering via the direct activation of FLOWERING LOCUS T and LEAFY in Arabidopsis thaliana[J]. The Plant Journal,2016,85(1):96-106.
[9] 郭君洁,贵红霞,武欣,等. 盐胁迫下过表达BoRACK1基因对羽衣甘蓝种子萌发的影响[J]. 植物生理学报,2018,54(8):1365-1371.
[10]Bai M J,Sun J J,Liu J Y,et al. The B-box protein BBX19 suppresses seed germination viainduction of ABI5[J]. The Plant Journal,2019,99 (6):1192-1202.
[11]Kim S G,Lee A K,Yoon H K,et al. A membrane-bound NAC transcription factor NTL8 regulates gibberellic acid-mediated salt signaling in Arabidopsis seed germination[J].The Plant Journal,2008,55(1):77-88.
[12]Jisha K C, Vijayakumari K,Puthur J T. Seed priming for abiotic stress tolerance: an overview[J]. Acta Physiologiae Plantarum,2013,35(5):1381-1396.
[13]Ibrahim E A. Seed priming to alleviate salinity stress in germinating seeds[J]. Journal of Plant Physiology,2016,192:38-46.
[14]Xie Z,Zhang Z L,Hanzlik S,et al. Salicylic acid inhibits gibberellin-induced alpha-amylase expression and seed germination via a pathway involving an abscisic-acid-inducible WRKY gene[J]. Plant Molecular Biology,2007,64 (3):293-303.
[15]Xie Z,Zhang Z L,Zou X,et al. Interactions of two abscisic-acid induced WRKY genes in repressing gibberellin signaling in aleurone cells[J]. The Plant Journal,2006,46(2):231-242.
[16]Wang C,Deng P Y,Chen L L. A wheat WRKY transcription factor TaWRKY10 confers tolerance to multiple abiotic stresses in transgenic tobacco[J]. PLoS ONE,2013,8(6):e65120.
[17]江文波,余迪求. 擬南芥WRKY2转录调控因子可能参与调控渗透胁迫反应[J]. 云南植物研究,2009,31(5):427-432.[JP]
[18]Ding Z J,Yan J Y,Li G X,et al. WRKY41 controls Arabidopsis seed dormancy via direct regulation of ABI3 transcript levels not downstream of ABA[J]. The Plant Journal,2014,79(5):810-823.
[19]Yu Y C,Wang L,Chen J C. WRKY71 acts antagonistically against salt-delayed flowering in Arabidopsis thaliana[J]. Plant and Cell Physiology,2018,59(2):414-422.
猜你喜欢
热带作物学报(2020年7期)2020-09-02
山东农业科学(2017年12期)2018-01-09
现代农业科技(2017年1期)2017-03-06
现代农业科技(2016年21期)2017-03-06
现代经济信息(2016年4期)2016-06-20
企业文化·中旬刊(2015年12期)2016-03-09
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
热带农业科学(2014年8期)2014-09-23
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
