
河西走廊不同生态型芦苇种群生殖分株的生物量分配与异速生长1)
2020-06-27 16:08独肖艳焦润安焦健陈丽张晓玮陈国鹏
东北林业大学学报 2020年6期
独肖艳 焦润安 焦健 陈丽 张晓玮 陈国鹏
(甘肃农业大学,兰州,730070)
在漫长的发展和进化中,植物与环境相互作用,逐渐形成了许多内在生理和外在形态方面的适应对策,以最大程度地减小环境的不利影响。植物在外在形态方面的适应对策即为生态可塑性。对生态可塑性的认识,更多来自于植物响应变化的环境条件个体基因型所产生的表型的范围[1]。植物个体生长发育所处生境的差异导致植物表现出丰富的可塑性调节[2],不仅影响到各自在整个生育期的生长发育过程,而且最终都要表现在大小、多少等数量性状上[3]。植物形态可能随植株大小的变化而变化,因此,异速生长分析对研究植物生长形态非常必要[4]。植物的异速生长与生物量分配、植物对资源的利用、对异质环境的形态适应有密切关系[5]。
河西走廊地处巴丹吉林沙漠南缘荒漠地带,近几十年来在人类干扰和区域气候变化双重驱动下,该区域地下水位逐年下降,土壤旱化、盐渍化,植被生长受到限制[6],急需进一步加强区域植被保护工作,维护生态安全。在内陆荒漠气候条件下,土壤水、盐条件对河西走廊植物群落数量特征的影响具有同等重要性[7]。
芦苇(Phragmitescommunis),根茎型无性系禾草,生长于池沼、河岸、河口、湖滨、荒漠、盐碱区等地[8]。世界各地均有生长,河西走廊也广泛分布,是该地区植被的主要建群种之一,约占耕地总面积的50%[9-10]。芦苇萌生能力强,众多根茎串联交织,增加了下垫面的粗糙度,具有降低风速和减轻风沙危害的作用,对河西走廊沙漠化的扩张形成有效的遏制作用。芦苇有较强的生态适应性,其生态可塑性也比较可观。以往对芦苇的研究多集中于湿润半湿润地区的水生芦苇[11-15],对于不同生态型芦苇有干旱、盐碱的适应性比较[16]及分子生物学[17-18]方面的研究报道,近年来有少量关于河西走廊芦苇光合作用[19-20]及群落格局[21]研究,但河西走廊地区不同生态型芦苇种群生态可塑性方面的研究还鲜见报道。张承烈等[8,22]对河西走廊不同生境中的芦苇划分为种下不同生态型,证实了其合理性,本研究依据其划分选择湿地生境的湿生芦苇、沙地生境的沙生芦苇以及盐碱生境的盐碱芦苇,对3种生境不同生态型芦苇生殖分株的生物量分配、异速生长和其对土壤因子的响应进行分析研究,籍以揭示芦苇种群逆境适应策略及其生态可塑性,为河西走廊芦苇种群的保护和植被恢复提供理论依据。
1 研究区概况
以位于巴丹吉林沙漠的甘肃省临泽县为定位研究区。临泽县处于99°51′~100°30′E,38°57′~39°42′N,位于河西走廊中部,属大陆性荒漠草原气候。年平均气温为7.7 ℃,年均降水量118.4 mm,蒸发量1 830.4 mm。光照和水分配置极不均衡,土壤次生盐渍化严重,植物的生长环境十分恶劣。该地区芦苇起源于沼泽生境中,长期生长于沙丘或盐碱化土壤中,其形态特征与各自生长环境相适应,产生了的极大变异[23]。
2 研究方法
2018年9月依据生境土壤条件差异,选择湿地、沙地和盐碱地设置样地,3种生境样地分布于5 km以内,故认为其气候条件相同。以湿地为对照样地,每种生境设3个20 m×20 m的样地:①湿地生境(H1),调查样地设在临泽县平川镇三一村黑河岸边,生长湿生芦苇,种群密度较大;②沙地生境(H2),样地设于鸭暖镇古寨村沙地,生长沙生芦苇,种群密度最小;③盐碱生境(H3),位于临泽县鸭暖镇马营村盐碱地,生长盐碱芦苇,种群密度最大。
在不同生境设置的各样地上,均随机取20株芦苇生殖分株,每个生境3个样地,共60株,齐地面剪下,混匀后按不同生境随机抽取30株,测量各生殖分株的株高和花序长,随后带回实验室,放置烘箱内80 ℃烘至恒质量,分别测量各个分株的叶片干质量、叶鞘干质量,花序干质量和茎干质量。
由于芦苇根茎分布在1 m以内土层,将各生境样地按土层间隔10 cm深度分为10层,用土钻(直径2 cm)进行土壤取样,3次重复,将之混匀带回实验室,用烘干法测定土壤含水量,质量法[24]测定土壤含盐量(土水比为1∶5)。
生殖生长比率、生殖分配、生产分配和茎生物量分配计算公式[25]分别为:
生殖生长比率(RRG)=(花序长/株高)×100%;
生殖分配(RA)=(花序生物量/总生物量)×100%;
生产分配(PA)=(叶生物量/总生物量)×100%;
茎生物量分配(SA)=(茎生物量/总生物量)×100%。
用SPSS 24.0软件对数据进行分析处理,对不同生境间芦苇分株构件指标的差异进行单因素方差分析;分株各构件指标与土壤因子的关系进行Pearson相关分析。分析异速生长关系时用幂函数Y=βXα来表示,其中,β为标准化常数,α为异速指数。当α=1时,为等速关系,而当α≠1时,为异速关系[26];分析生殖生长量与分株高关系时,在线性、指数和幂3种函数中,选用相关性最高的作为性状间关系定量分析模型,方程的拟合优度用R2和P的值进行检验。
3 结果与分析
3.1 生殖分株构件特征
湿生芦苇,沙生芦苇和盐碱芦苇生殖分株各构件指标均有显著性差异(P<0.05),表明其生态可塑性良好;3种生态型芦苇各构件指标的变异系数除叶鞘生物量和茎生物量外,由大到小均呈现盐碱芦苇、沙生芦苇、湿生芦苇的特征,叶鞘生物量的变异系数由大到小排序为沙生芦苇、湿生芦苇、盐碱芦苇,茎生物量变异系数由大到小为沙生芦苇、盐碱芦苇、湿地芦苇(表1)。
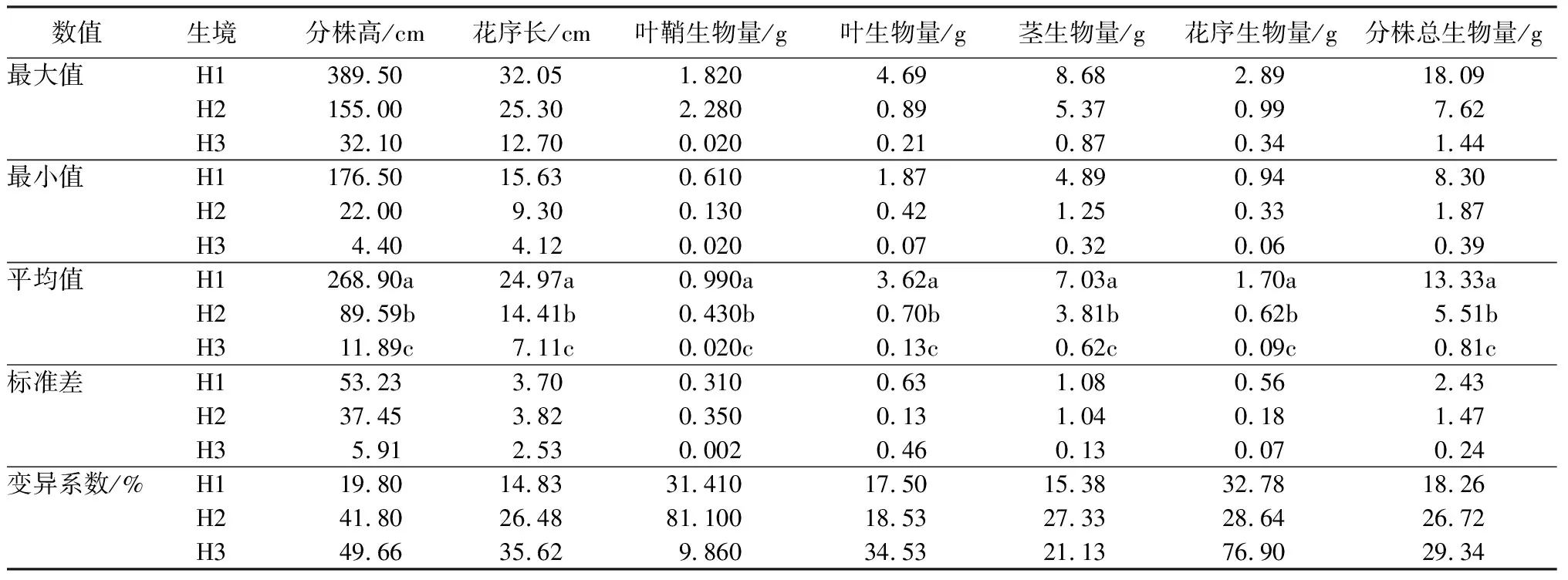
表1 3种生境芦苇种群生殖分株的特征
注:H1为湿地生境;H2为沙地生境;H3为盐碱地生境;同列不同小写字母表示差异显著(P<0.05)水平。
各生态型芦苇分株叶、茎和花序生物量的分配均有显著性差异(P<0.05);沙生芦苇和湿生芦苇的生殖生长比率不具有显著性差异(P>0.05),但盐碱芦苇与湿生芦苇具有显著性差异(P<0.05);各生态型芦苇生殖生长比率和生产分配变异系数由大到小均呈现沙生芦苇、盐碱芦苇、湿生芦苇的趋势,而生殖分配和茎生物量分配由大到小为盐碱芦苇、沙生芦苇、湿生芦苇(表2),表明不同生态型芦苇生殖分株构件生物量分配有一定差异性。
表23种生境芦苇种群生殖分株叶、茎和花序生物量分配特征
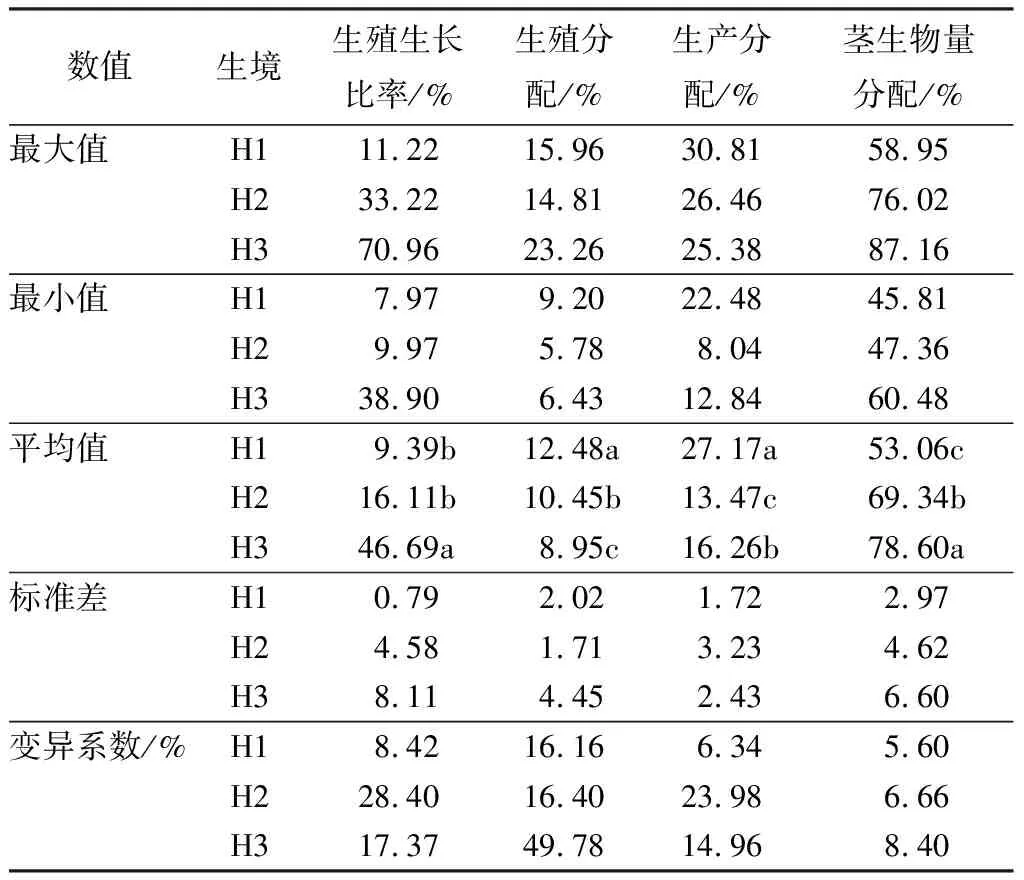
数值生境生殖生长比率/%生殖分配/%生产分配/%茎生物量分配/%最大值H111.2215.9630.8158.95H233.2214.8126.4676.02H370.9623.2625.3887.16最小值H17.979.2022.4845.81H29.975.788.0447.36H338.906.4312.8460.48平均值H19.39b12.48a27.17a53.06cH216.11b10.45b13.47c69.34bH346.69a8.95c16.26b78.60a标准差H10.792.021.722.97H24.581.713.234.62H38.114.452.436.60变异系数/%H18.4216.166.345.60H228.4016.4023.986.66H317.3749.7814.968.40
注:H1为湿地生境;H2为沙地生境;H3为盐碱地生境;同列不同小写字母表示差异显著(P<0.05)水平。
3.2 生殖生长量与分株高度的关系
3种生态型芦苇分株的花序长与株高呈现显著正相关关系(P<0.01),沙生芦苇和湿生芦苇生殖生长比率与株高呈现显著负相关关系(P<0.01),盐碱芦苇生殖生长比率与株高无显著负相关关系(P>0.05)(表3);随着分株高度的增加,花序长度的增长幅度由大到小为盐碱芦苇、沙生芦苇、湿生芦苇,生殖生长比率的降低幅度为沙生芦苇大于湿生芦苇(表3)。
表3湿地、沙地、盐碱生境芦苇生殖生长量(y)与分株高度(x)的拟合方程及显著性检验
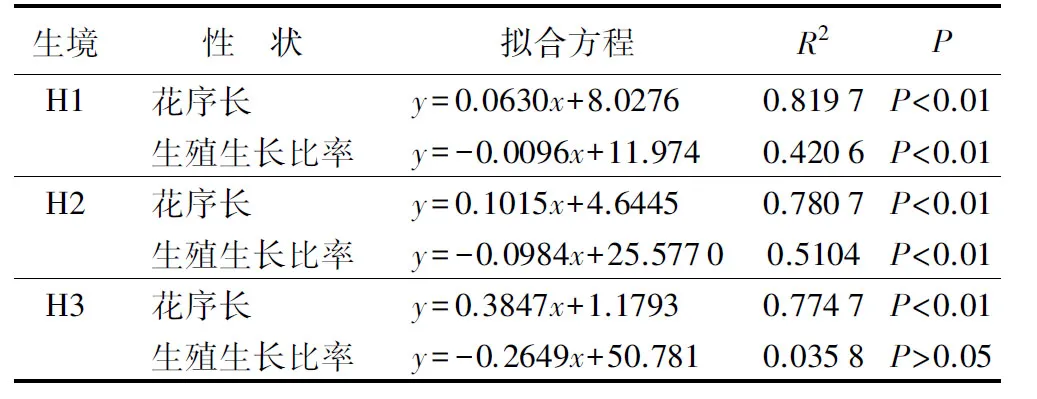
注:H1为湿地生境;H2为沙地生境;H3为盐碱地生境。
3.3 分株异速生长分析
3种生态型芦苇分株总生物量、茎和叶鞘生物量、叶生物量以及花序生物量与分株高度之间的关系具有相似性,均随分株高度的增加呈现幂函数增长规律;随分株高度的增加,除茎和叶鞘生物量增长速率由大到小为湿生芦苇、沙生芦苇、盐碱芦苇,其余构件生物量均由大到小为湿生芦苇、盐碱芦苇、沙生芦苇;湿生芦苇各构件生物量的增长速率均明显大于其余两种生态型芦苇;同一生态型芦苇花序生物量增长速率明显大于其余构件指标,最大幂值达到1.531 5;所有方程的确定系数R2在0.630 7~0.932 9(表4)。
表4湿地、沙地、盐碱生境芦苇分株生物量(y)与分株高度(x)的拟合方程及显著性检验
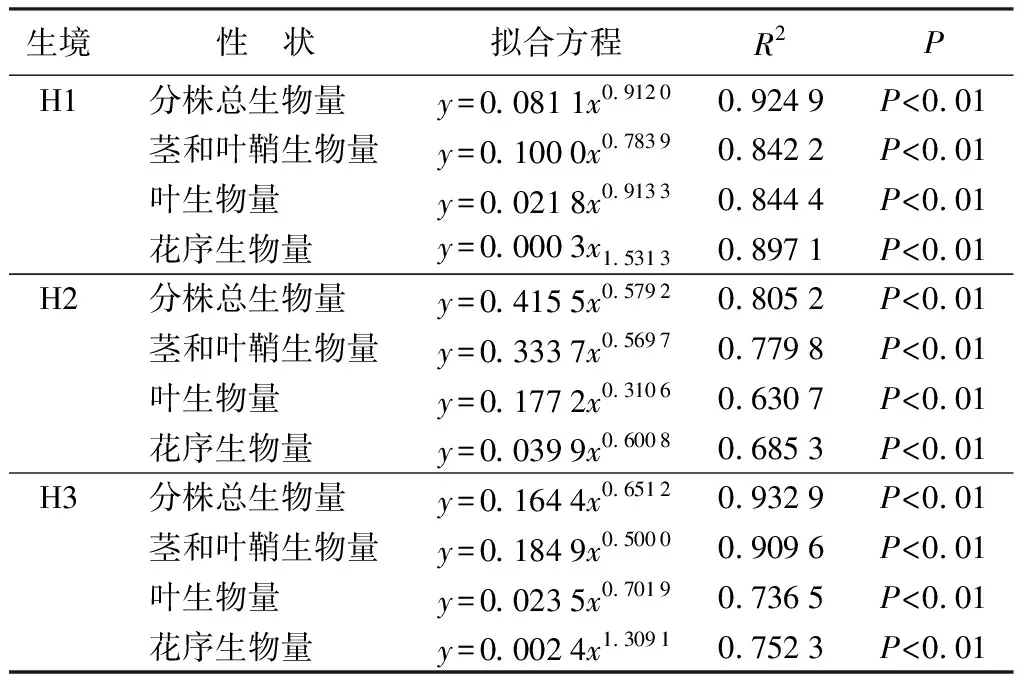
生境性 状拟合方程R2PH1分株总生物量y=0.0811x0.91200.9249P<0.01茎和叶鞘生物量y=0.1000x0.78390.8422P<0.01叶生物量y=0.0218x0.91330.8444P<0.01花序生物量y=0.0003x1.53130.8971P<0.01H2分株总生物量y=0.4155x0.57920.8052P<0.01茎和叶鞘生物量y=0.3337x0.56970.7798P<0.01叶生物量y=0.1772x0.31060.6307P<0.01花序生物量y=0.0399x0.60080.6853P<0.01H3分株总生物量y=0.1644x0.65120.9329P<0.01茎和叶鞘生物量y=0.1849x0.50000.9096P<0.01叶生物量y=0.0235x0.70190.7365P<0.01花序生物量y=0.0024x1.30910.7523P<0.01
注:H1为湿地生境;H2为沙地生境;H3为盐碱地生境。
3种生态型芦苇茎和叶鞘生物量、叶生物量以及花序生物量均随分株生物量的增加呈现幂函数增长规律(表5);随分株总生物量的增加,茎和叶鞘生物量的增长速率由大到小为沙生芦苇、湿生芦苇、盐碱芦苇,其幂值在0.764 8~0.994 3,而叶生物量、花序生物量的增长速率均由大到小为盐碱芦苇、湿生芦苇、沙生芦苇,叶生物量幂值在0.495 6~1.128 1,花序生物量幂值在1.314 4~2.932 6;所有方程的确定系数R2在0.668 9~0.989 4(表5)。
表5湿地、沙地、盐碱生境芦苇分株组分生物量(y)与总生物量(x)的拟合方程及显著性检验
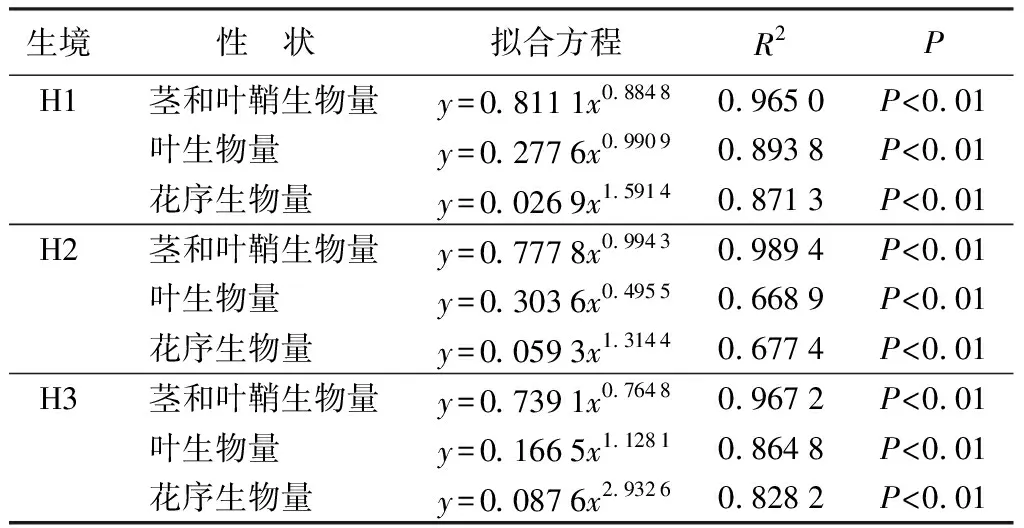
生境性 状拟合方程R2PH1茎和叶鞘生物量y=0.8111x0.88480.9650P<0.01叶生物量y=0.2776x0.99090.8938P<0.01花序生物量y=0.0269x1.59140.8713P<0.01H2茎和叶鞘生物量y=0.7778x0.99430.9894P<0.01叶生物量y=0.3036x0.49550.6689P<0.01花序生物量y=0.0593x1.31440.6774P<0.01H3茎和叶鞘生物量y=0.7391x0.76480.9672P<0.01叶生物量y=0.1665x1.12810.8648P<0.01花序生物量y=0.0876x2.93260.8282P<0.01
注:H1为湿地生境;H2为沙地生境;H3为盐碱地生境。
3.4 土壤水、盐含量对分株各构件的影响
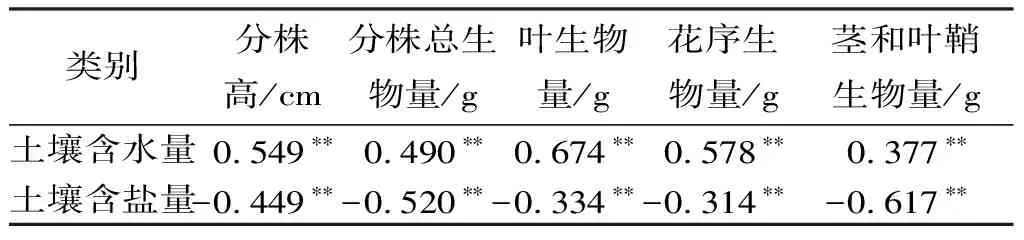
不同生境样地土壤含水量有显著性差异(P<0.05),湿地土壤含水量最大(51.8%),盐碱地次之,沙地最小(9.2%);盐碱生境土壤含盐量和湿地、沙地生境均有显著性差异(P<0.05),但湿地和沙地两生境间无显著性差异(P>0.05),土壤含盐量以盐碱地最高,湿地次之,沙地最低(表6)。分株高、分株总生物量、叶生物量、花序生物量、茎和叶鞘生物量与土壤含水量呈极显著正相关(P<0.01),与土壤含盐量呈极显著负相关(P<0.01)(表7)。可以认为,土壤水、盐含量为影响芦苇分株生长发育的驱动因素。

表6 不同生境样地土壤水、盐含量
注:H1为湿地生境;H2为沙地生境;H3为盐碱地生境,同列不同小写字母表示差异显著(P<0.05)水平,表中数据为平均值±标准差。
表7芦苇种群分株高、各构件生物量与土壤水、盐含量间的相关系数
类别分株高/cm分株总生物量/g叶生物量/g花序生物量/g茎和叶鞘生物量/g土壤含水量0.549∗∗0.490∗∗0.674∗∗0.578∗∗0.377∗∗土壤含盐量-0.449∗∗-0.520∗∗-0.334∗∗-0.314∗∗-0.617∗∗
注:** 表示P<0.01差异极显著。
4 讨论
4.1 不同生境芦苇生物量分配
盐碱芦苇分株总生物量较湿生芦苇相差16.5倍,分株高度相差22.6倍;沙地芦苇分株总生物量较湿生芦苇相差2.4倍,分株高度相差3.0倍。湿生芦苇分株高和分株总生物量的生态可塑性差异不大,沙生芦苇和盐碱芦苇分别有15.1%和20.3%的差异。分株高的生态可塑性要高于分株总生物量,这与杨允菲[27],焦德志[13]在松嫩平原对芦苇的研究有所区别。河西走廊地区芦苇受干旱和盐碱的交互影响。张爽等[28]通过不同盐度水进行芦苇的灌溉处理,表明盐度高于0.6%时,株高会出现显著的抑制效果。罗文泊等[29]研究表明,随着水位的上升,湿生植物会将更多的光合作用产物向地上器官分配,使茎干伸长,增强对O2、CO2及光资源的利用。基于前人研究结果,结合本研究我们可以认为,盐碱环境对株高的影响较干旱更为明显。植物高度的生态可塑性变化,可以大大增加对环境的综合适应能力[30]。分株高的生态可塑性高于分株总生物量是芦苇植株适应河西地区逆境的结果。
3种生态型芦苇,叶生物量、花序生物量和茎生物量占分株总生物量的比重以茎为最大,这与刘香秀等[25]、杨允菲等[27]在松嫩平原测定的叶生物量占比重最大有所区别。究其原因,河西地区光照充足,降雨量少,蒸发量大,且处于盐碱环境,植物叶片扩展性受到一定的抑制,进而使得植物水分不会严重的流失[31],从而造成叶生物量的减少。茎具有贮藏、养分输导和支撑叶片扩展生长空间的功能[32],3种生态型芦苇分株茎生物量所占比重最大,体现芦苇凭借茎的伸长生长,提高分株对空间以及光资源的竞争力,使芦苇植株能够有效的使用环境方面的压力[27]。
分株的茎节伸长属于营养生长,花序节伸长属于生殖生长,其生殖生长量可用花序长度和生殖生长比率来衡量,前者反映出生殖分株的营养生长和生殖生长按比例伸长的协调性,后者反映出其营养生长和生殖生长的分配比例[33]。芦苇分株的花序长与株高呈现明显的正相关关系,湿生芦苇和沙生芦苇生殖生长比率与株高呈现负相关关系。湿生芦苇种群密度较大且水分条件最好,芦苇分株的生殖生长比率会降低,为了增加其竞争力,相对的会增加茎节的生长;沙生芦苇密度最小且水分条件最差,芦苇分株会增大生殖生长比率来提高潜在的生育力;盐碱芦苇种群密度最大,水分条件介于上述两生态型芦苇之间,但其土壤含盐量达到0.61%,对芦苇植株的抑制作用显著,其生殖生长和营养生长均处于较低水平。这一结论与张春华等[34]对光稃茅香生殖生长量和李红等[35]对羊草生殖生长量的研究结果相似,蕴含着植株生长与物质分配相协调原则。
4.2 不同生境芦苇异速生长
同速生长型和异速生长型是植物生长过程中的两种模式。同速生长型往往体现在数量性状间的关系为直线函数形式,而异速生长可通过幂函数和指数函数来刻画数量性状之间的关系,由遗传物质所决定,也受外界条件的影响[36]。用拟合方程的确定系数R2来估计遗传因子的控制水平,用1-R2来估计环境因素的影响水平[27,37-38]。芦苇在分株生物量与分株高度的异速生长中,沙生芦苇受环境影响最大,其中叶的R2达到0.630 7,受环境影响水平达到36.93%;在茎和叶鞘、叶、花序的相对生长中,叶和花序受环境影响最大的是沙生芦苇,最小的是湿生芦苇,而茎和叶鞘生物量受环境影响最小的是沙生芦苇,湿生芦苇和盐碱芦苇水平相当。从整体水平来说,遗传因子对芦苇分株的控制力更大,但沙生芦苇受环境因素的影响较为明显;在芦苇分株各组分异速生长中,叶受环境因素的影响较大。依据生长模型界定无性系空间发展策略:指数采取的是无性系内部空间的优先补充策略,线性增长采取的是无性系外部空间的优先扩展策略,幂函数增长采取的则是无性系内、外部空间兼顾的补充和拓展策略[39]。由此我们可知:河西走廊不同生境芦苇采用的是无性系内、外部空间兼顾的补充和拓展策略。
4.3 分株构件对土壤水、盐含量的响应
河西走廊地处沙漠-绿洲过渡带,该地区在很小的尺度上形成了环境的异质性,主要是土壤条件的异质性。本研究选取的3种生境除沙地生境和湿地生境土壤含盐量不具有显著性差异,其余两两间土壤水、盐含量均有极显著性差异,是造成该地区芦苇种下不同生态型的驱动因素。以往的研究也表明土壤环境因子对芦苇数量特征影响较大。土壤含水量是影响银川平原芦苇生态特征的最关键因子,其次是pH和全盐含量[40],扎龙湿地影响芦苇分株生态可塑性变异的主要驱动因子是土壤含水量[41],黄河三角洲芦苇的高度和茎粗与水深显著相关[42]。可见,尽管芦苇种群生长发育是生境条件综合作用的结果,但不论是在大尺度或者小尺度水平上,土壤环境因子都是研究芦苇生长发育必不可少的因素。
5 结论
同一构件生物量在不同生态型芦苇间均有显著性差异(P<0.05),分株对叶、茎和花序生物量的分配也存在显著性差异(P<0.05),且对茎生物量的分配比重最大;分株株高的生态可塑性要高于分株总生物量,盐碱环境对株高的影响较干旱更为明显;芦苇分株的花序长与株高呈显著正相关关系,湿生芦苇和沙生芦苇生殖生长比率与株高呈现负相关关系,盐碱芦苇生殖生长比率与株高无显著负相关关系。
3种生态型芦苇分株总生物量和各组分生物量随分株高度的增加,及茎和叶鞘、叶、花序的生物量随分株总生物量的增加均呈现幂函数异速增长规律。不同生态型芦苇分株之间,同一生态型芦苇不同组分之间的生长规律存在差异,具有较大的可调节性和生态可塑性,蕴含着植株生长与物质分配相协调原则。河西走廊不同生境芦苇采用的是无性系内、外部空间兼顾的补充和拓展策略。
沙地生境和湿地生境土壤含盐量不具有显著性差异(P>0.05),其余两两生境间土壤水、盐含量均有极显著性差异(P<0.05);土壤水、盐含量是造成河西地区芦苇种下不同生态型的驱动因素。
猜你喜欢
现代装饰(2022年5期)2022-10-13
作物学报(2022年8期)2022-05-29
中国农村水利水电(2021年11期)2021-11-29
金桥(2021年11期)2021-11-20
现代畜牧科技(2021年4期)2021-07-21
风景园林(2021年2期)2021-04-24
落叶果树(2021年6期)2021-02-12
北方水稻(2020年6期)2020-12-12
现代装饰(2020年8期)2020-08-24
商周刊(2018年12期)2018-07-11
