
舟山近海3 种鰧科鱼类及其DNA 条形码研究
2020-06-19 17:04任米佳俞正森徐胜勇徐开达徐汉祥高天翔
浙江海洋大学学报(自然科学版) 2020年5期
任米佳,俞正森,徐胜勇,徐开达,徐汉祥,高天翔
(1.浙江海洋大学水产学院,浙江舟山 316022;2.中国海洋大学水产学院,山东青岛 266003;3.浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江舟山 316021)
鰧科Uranoscopidae 鱼类隶属于硬骨鱼纲Osteichthyes、鲈形目Perciformes、龙鰧亚目Trachinoidei[1]。Fishbase(Froese and Pauly,2019)资料显示现生鰧科鱼类共有8 属53 种,在中国已知有3 属7 种。在《浙江海洋鱼类志》[1]755中记载浙江省鰧科鱼类共有3 属5 种,分别为日本鰧Uranoscopus japonicusHouttuyn,1782、土佐鰧Uranoscopus tosae(Jordan &Hubbs,1925)、少鳞鰧Uranoscopus oligolepisBleeker,1878、披肩鰧Ichthyscopus lebeck(Bloch &Schneider,1801) 和青鰧Xenocephalus elongatus(Temminck &Schlegel,1843)。鰧科鱼类作为经济鱼类[1],少量出现于兼捕渔获物中,目前仅见一些鰧科鱼类分类学研究介绍[2-5],而关于鰧科鱼类生物学、遗传学的研究报道极少。
线粒体DNA (mitochondrial DNA) 是线粒体中的遗传物质。鱼类线粒体DNA 包括13 个蛋白质编码基因、22 个转运RNA、2 个核糖体RNA、1 个非编码区和1 个轻链复制起始区。它们具有自我复制、转录和翻译的能力[6]。2003 年HEBERT,et al[7]提出DNA 条形码(DNA barcoding)作为物种鉴定的标记,以此建立DNA 序列和生物物种间的对应关系。DNA 条形码是指生物体内能够代表物种的、标准的、有足够变异的、易扩增且相对较短的DNA 片段[8],目前已被广泛应用于物种鉴定等研究。例如,2006 年YOSHIDA,et al[9]利用线粒体12S rRNA 及COⅠ序列探讨乌贼科Sepiidae 的系统发育关系;2013 年李渊等[10]利用COⅠ基因对鲳属Pampus鱼类进行分类研究;2015 年王燕平[11]基于COⅠ序列探讨蛇鲻属Saurida鱼类的系统发育关系;2016 年张静等[12]利用DNA 条形码技术对中国沿海分布的6 种棱鳀属Thryssa鱼类样品进行了物种鉴定;2018 年赵娜等[13]基于DNA 条形码可以对除南极小带腭鱼Cryodraco antarcticus和罗斯海小带腭鱼Cryodraco atkinsoni之外的南极鱼类物种进行鉴定。2015 年MIYA,et al[14]设计了12S rRNA 基因片段引物作为metabarcoding,主要用于海洋鱼类多样性监测研究。
本研究对近年来采集的鰧科鱼类进行形态特征分析,并测定线粒体12S rRNA 和COⅠ基因片段,结合GenBank 中同源序列进行比较分析,探讨鰧科鱼类种间遗传变异程度及其系统发育关系,以期为鰧科鱼类分类的深入研究提供基础资料。
1 材料与方法
1.1 实验材料
于2017 年4 月-2019 年10 月在浙江舟山近海采集日本鰧、土佐鰧和青鰧标本。形态学实验所用日本鰧27 尾,体长范围为148.5~261.9 mm,体质量范围为25.73~337.9 g;土佐鰧5 尾,体长范围为142.5~183.5 mm,体质量范围为131.1~234.3 g;青鰧24 尾,体长范围为239.5~392.5 mm,体质量范围为378.9~1 807.9 g(表1)。所有标本保存于浙江海洋大学渔业生态与生物多样性实验室。

表1 本研究所用鰧科样品信息Tab.1 Information of Uranoscopidae samples in this study
1.2 实验方法
1.2.1 形态学研究
对样品进行形态学测定[10]。用游标卡尺测量体长、头长、吻长、眼径、眼间距、眼后头长、尾柄长、尾柄高、第一背鳍基长、第二背鳍基长、胸鳍基长、腹鳍基长和臀鳍基长,精确到0.1 mm,各测量指标见图1(以日本鰧为例)。对背鳍鳍条数、臀鳍鳍条数、尾鳍鳍条数、胸鳍鳍条数和腹鳍鳍条数进行计数。用电子天平测定样品质量,精确到0.1 g。

图1 日本鰧的形态学测量指标Fig.1 Morphometric characters for U.japonicus
1.2.2 DNA 提取、扩增和测序
剪取样品肌肉,采用苯酚-氯仿法提取DNA[15]。将乙醇沉淀后的基因组DNA 溶解于100 μL 灭菌水中,并保存于4 ℃冰箱备用。所得的DNA 用1.5%的琼脂糖凝胶电泳检测,并用于PCR 扩增。
本研究扩增的基因片段为12S rRNA 及COⅠ基因片段。线粒体DNA 的12S rRNA 基因扩增引物为Mifish-U-F:5’-GTCGGTAAAACTCGTGCCAGC-3’和Mifish-U-R:5’-CATAGTGGGGTATCTAATCCCAGTTTG-3’[11],COI 基因扩增引物为F1:5’-TCAACCAACCACAAAGACATTGGCAC-3’,R1:5’-TAGACTTCTGGGTGGCCAAAGAATCA-3’及R2:5’-ACTTCAGGGTGACCGAAGAATCAGAA-3’[16]。PCR 反应总体积25 μL,其中10×buffer 2 μL、正反向引物各1 μL、DNTPs 2 μL、ExTaq酶0.15 μL、灭菌水17.5 μL、ExTaq酶0.15 μL 以及DNA 模板1 μL。反应条件为:94 ℃预变性5 min;94 ℃变性45 s,54 ℃退火30 s,72 ℃延伸45 s,共35 个循环;最后72 ℃延伸10 min。扩增产物经电泳检测后,送上海美吉生物科技有限公司进行双向测序。
1.2.3 数据分析
将测得的mtDNA 12S rRNA 及COⅠ基因片段用DNAStar(DNASTAR,Inc)软件包中的Seqman 软件进行序列比对,并辅以人工校正;采用NCBI 在线BLAST 与网上序列对比;使用Arlequin 软件计算碱基组成、多态位点、单倍型多样性(haplotype diversity,h)、核苷酸多样性(nucleotide diversity,π)等数据,使用MEGA 6.0 计算Kimura-2-parameter 遗传距离,并用黄鳍刺虾虎鱼Acanthogobius flavimanus作为外群,基于邻接法(Neighbor-Joining)构建系统树分析3 种鰧科鱼类的亲缘关系,并对所得系统树进行自展法检验(1 000 次重复)。
2 结果
2.1 形态描述
在形态特征中,日本鰧、土佐鰧与青鰧的差异主要表现在背鳍和肱棘特征上。日本鰧和土佐鰧都具有2 个背鳍,且肱棘强大,而青鰧只具有1 个背鳍,且肱棘不明显。3 种鰧科鱼类可数、可量性状比较见表2。

表2 3 种鰧科鱼类可数、可量性状比较Tab.2 Meristic characters and proportional measurements of the three Uranoscopidae species
2.1.1 日本鰧Uranoscopus japonicusHouttuyn,1782
测量样品27 尾,全长范围为175.6~251.1 mm,体长范围为136.4~192.6 mm,D.Ⅳ-14;P.18;V.Ⅰ-5;A.15;C.16。
体长为体高的4.2~5.7 倍,为头长的3.9~4.1 倍,为体宽的5.8~6.0 倍;头长为吻长的6.9~8.2 倍,为眼径的10.9~11.1 倍,为眼间距的3.7~4.9 倍;眼间距为眼径的2.2~3.0 倍;体长为腹鳍基长的10.3~10.9 倍,为胸鳍基长的30.6~32.5 倍;体长为尾柄长的8.4~8.9 倍;尾柄长为尾柄高的1.04~1.21 倍。
体延长,头粗大,吻短。体背面中央与眼间隔间有1 凹窝。眶前骨下缘有2 个短骨突。鼻孔2 个,位于吻前端。鳃盖后方胸鳍上方具2 个肱棘,后棘特别尖长。前鳃盖骨下缘有4~5 个尖棘,下鳃盖骨下方有1埋在皮内的尖棘。
体被小圆鳞,鳞斜向后下方。头部、颈背部、胸腹部、背鳍基均无鳞。侧线1 条,位高。体背侧呈黄褐色,腹部白色,体两侧及背面具白色斑点。第一背鳍黑色,基底白色;第二背鳍淡黄色。臀鳍白色。胸鳍黄色。腹鳍淡红色。尾鳍黄色,后缘白色。
日本鰧分布于西北太平洋海域,我国沿海均有分布,属广温性底层中小型鱼类,多栖息于沙泥质浅海水域。详细体态特征见图4。

图2 日本鰧(192.6 mm SL,226 g)Fig.2 Uranoscopus japonicus Houttuyn,1782
2.1.2 土佐鰧Uranoscopus tosae (Jordan &Hubbs,1925)
测量样品5 尾,全长范围为188.5~228.5 mm,体长范围为142.5~183.5 mm,D.Ⅳ-13;P.19;V.Ⅰ-5;A.13;C.17。
体长为体高的3.26~4.57 倍,为头长的2.8~2.9 倍,为体宽的2.9~3.2 倍;头长为吻长的8.5~8.7 倍,为眼径的5.8~9.3 倍,为眼间距的3.7~4.5 倍;眼间距为眼径的1.6~2.1 倍;体长为腹鳍基长的5.1~5.3 倍,为胸鳍基长的17.0~18.2 倍;体长为尾柄长的10.7~13.1 倍;尾柄长为尾柄高的1.2~1.4 倍。
体长形,头粗大,稍平扁,近四棱形,吻短钝。眼间隔中央微凹。头部中央凹入,形成1 凹刻。眶前骨在吻前端有2个短棘突。头部两侧各有1 低纵骨棱,鳃盖后方胸鳍上方具2 个肱棘。鼻孔2 个,位于吻前端。上、下颌缘边均有1行小须状突起。前鳃盖骨下缘有6 个短棘,下鳃盖骨下端有1 短小棘。
体被圆鳞,细小,半埋于皮下,呈斜行排列。仅喉部到肛门间和胸鳍基部附近无鳞。侧线完全,位高,自鳃盖后上方起沿背缘向后延伸,在尾柄中部斜折至尾鳍基中部。体背侧黄褐色,身上无特殊斑纹,腹侧灰白色,第一背鳍黑色,下缘黄白色,第二背鳍、胸鳍与尾鳍灰黄色,腹鳍与臀鳍白色。口腔与鳃腔白色。
土佐鰧分布于西北太平洋海域,我国产于南海、东海及台湾海域,属于暖水性底栖中小型鱼类,栖息水深可达420 m。详细体态特征见图3。

图3 土佐鰧(193.1 mm SL,235 g)Fig.3 Uranoscopus tosae (Jordan &Hubbs,1925)
2.1.3 青鰧Xenocephalus elongatus(Temminck &Schlegel,1843)
测量样品24 尾,全长范围为312.3~470.5 mm,体长范围为289.5~432.1 mm,D.13;P.19~20;V.Ⅰ-5;A.18;C.12~13。
体长为体高的6.6~7.7 倍,为头长的3.9~4.2 倍,为体宽的7.3~8.4 倍;头长为吻长的8.6~10.4 倍,为眼径的8.9~10.4 倍,为眼间距的4.2~5.4 倍;眼间距为眼径的3.0~3.2 倍;体长为腹鳍基长的12.2~12.7 倍,胸鳍基长的35.0~41.2 倍;体长为尾柄长的15.3~17.2 倍;尾柄长为尾柄高的1.10~1.16 倍。
体长形,头中大,吻短钝。眼间隔中央凹入。鼻孔2 个,位于吻前缘,前鼻孔后缘有1 皮瓣。口缘有1 行短毛状皮突。鳃盖后方胸鳍上方无肱棘。鳃孔大。峡部有2 个黑点。
体被很小圆鳞,大多埋于皮下。头部、胸部和腹部均无鳞。侧线1 条,位高。体背部灰青绿色,腹部淡青灰色,体背部及两侧上方具许多不规则褐色小斑,背鳍淡黄色。臀鳍、胸鳍及腹鳍淡棕色。尾鳍灰青色。
青鰧分布于西北太平洋海域,我国沿海均有分布。属于广温性较深海底栖中型鱼类,主要栖息于大陆架边缘泥沙质底的较深海区。详细体态特征见图4。

图4 青鰧(254.7 mm SL,498 g)Fig.4 Xenocephalus elongatus (Temminck&Schlegel,1843)
2.2 DNA 条形码分析
2.2.1 mtDNA 12S rRNA 基因片段序列
本实验共测得41 条12S rRNA 序列,其中日本鰧22 条,土佐鰧5 条,青鰧14 条,所有序列未出现碱基插入与缺失,片段长度均为173 bp。3 种鰧科鱼类的4 种碱基平均含量为:A 为34.65%,T 为16.83%,G为24.23% C 为24.24%,平均A+T 含量(51.53%)略高于G+C 含量(48.47%)(表3)。41 个序列中共有8 个单倍型,其中日本鰧5 个单倍型,土佐鰧2 个单倍型及青鰧1 个单倍型。在173 个位点中,保守位点129个,变异位点44 个,简约信息位点41 个,单一突变位点3 个。日本鰧和土佐鰧单倍型多样性分别为0.337 7±0.127 8 和0.400 0±0.237 3,核苷酸多样性分别为0.002 2±0.002 4 和0.002 4±0.003 0。

表3 3 种鰧科鱼类12S rRNA 基因序列特征Tab.3 Characters of 12S rRNA gene fragments of the three Uranoscopidae species
2.2.2 mtDNA COⅠ基因片段序列
本实验共测得20 条线粒体COⅠ基因片段序列,其中日本鰧2 条,土佐鰧5 条,青鰧13 条,所有序列未出现碱基插入与缺失,片段长度为655 bp。3 种鰧科鱼类的4 种碱基平均含量为:A 为22.38%,T 为28.36%,G 为18.42%,C 为30.84%,平均A+T 含量(50.74%)高于G+C 含量(49.26%)(表4)。20 个序列中共测得8 个单倍型,其中日本鰧2 个单倍型,土佐鰧2 个单倍型及青鰧4 个单倍型。在655 个位点中,保守位点476 个,变异位点179 个,简约信息位点174 个,单一突变位点5 个。日本鰧、土佐鰧和青鰧的单倍型多样性分别为1.000±0.500 0、0.400 0±0.237 3 和0.423 1±0.164 5,核苷酸多样性分别为0.128 2±0.129 0、0.0018±0.001 6 和0.000 7±0.000 74。

表4 3 种鰧科鱼类CO I 基因序列特征Tab.4 Characters of CO I gene fragments of the three Uranoscopidae species
2.3 种内与种间遗传距离
采用MEGA 6.0 软件,基于Kimura 2-parameter 算法计算12S rRNA 及CO I 的净遗传距离(表5、6)。在12S rRNA 基因中,种内遗传距离均为0.002;种间遗传距离范围为0.147~0.216,其中日本鰧与土佐鰧的遗传距离最小(0.147),其次是土佐鰧与青鰧(0.209),日本鰧与青鰧遗传距离最大,为0.216。在CO I 基因中,3 种鰧科鱼类种内遗传距离范围为0.001~0.005;种间遗传距离范围为0.192~0.261,种间遗传距离最大的是土佐鰧与青鰧(0.261),日本鰧与土佐鰧遗传距离最小。

表5 基于12S rRNA 基因序列的种内(对角线)和种间(下三角)遗传距离Tab.5 Intraspecific (on diagonal) and interspecific (below diagonal) genetic distance based on 12S rRNA gene fragments

表6 基于CO I 基因序列的种内(对角线)和种间(下三角)遗传距离Tab.6 Intraspecific (on diagonal) and interspecific (below diagonal) genetic distance based on CO I gene fragments
2.4 系统发育分析
以黄鳍刺虾虎鱼作为外群,从GenBank 上下载3 种鰧科鱼类的12S rRNA 序列,日本鰧、土佐鰧和青鰧的引用序列号分别为AB974631、LC037149 和LC 506657;从GenBank 上下载3 种鰧科鱼类的CO I 序列,日本鰧和土佐鰧的引用序列号分别为AP017446 和KX641475,青鰧的引用序列号为KU892957 和JQ738420。采用邻接法(NJ)构建系统进化树(图4、5)。结果显示,基于12S rRNA 与CO I 基因构建的系统树结构基本一致,其中日本鰧和土佐鰧之间的亲缘关系较近,与青鰧的亲缘关系较远。
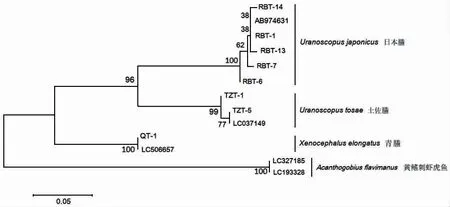
图5 基于12S rRNA 基因构建的单倍型NJ 树Fig.5 Phylogenetic tree of the three Uranoscopidae species based on 12S rRNA gene fragments
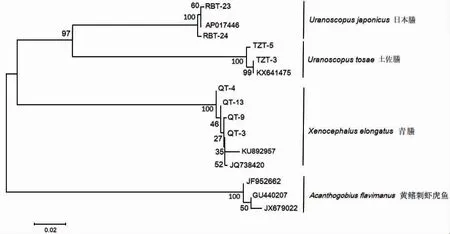
图6 基于COⅠ基因片段构建的单倍型NJ 树Fig.6 Phylogenetic tree of the three Uranoscopidae species based on CO I gene fragments
3 讨论
《鱼类分类学》中记载了瞻星鱼科Uranoscopoidae 有瞻星鱼属Uranoscopus和青鰧属Gnathagnus[2]。在《中国鱼类系统检索》中记载了我国鰧科有4 属6 种[4]。在《东海鱼类志》中,朱元鼎等[17]记录了我国鰧科有4 属5 种,4 属有鰧属Uranoscopus、项鳞鰧属Zalescopus、鱼鰧属Ichthyscopus和青鰧属Gnathagnus,5 种有日本鰧Uranoscopus japonicus(Houttuyn,1782)、少鳞鰧Uranoscopus oligolepis(Bleeker,1878)、项鳞鰧Uranoscopus tosae(Jordan &Hubbs,1925)、鱼鰧Ichthyscopus lebeck (Bloch &Schneider,1801)和青鰧Gnathagnus elongatus (Temminck &Schlegel,1843),其中将鰧属鱼类中具背鳍Ⅳ-13~14、臀鳍13~14、胸鳍17~18、腹鳍Ⅰ-5、尾鳍16~18、体背侧全部有白色网状斑纹、鳃孔后方具2 个尖肱棘和后棘特别尖长等特征的鱼种鉴定为日本鰧;将项鳞鰧属鱼类中具背鳍Ⅳ-13、臀鳍13、胸鳍19、腹鳍Ⅰ-5、尾鳍17、前鳃盖骨下缘有6 个短棘和下鳃盖骨下端有一短小棘的鱼种鉴定为项鳞鰧;将青鰧属鱼类中具背鳍12~13、臀鳍16~18、胸鳍18~20、腹鳍Ⅰ-5、尾鳍11~13、体背部灰青绿色且具不规则小斑、背鳍淡黄色和前鳃盖骨后缘无小突起等特征的鱼种鉴定为青鰧。在《福建鱼类志》中记载了我国鰧科有3 属5 种,其中鰧属Uranoscopus有日本鰧、双斑鰧Uranoscopus bicinctus(Temminck &Schlegel,1843)和少鳞鰧;鱼鰧属Ichthyscopus有鱼鰧;青鰧属Gnathagnus有青鰧[5]。在《浙江海洋鱼类志》一书中,赵盛龙等[1]760将中文名项鳞鰧改名为土佐鰧,日本鰧、土佐鰧和青鰧的形态学研究结果与《东海鱼类志》[17]的研究结果一致。山田[18]将具2 背鳍、鳃盖后方胸鳍上方肱棘明显的鰧科鱼类划分为鰧属,将鱼体背部具淡色斑纹、前鳃盖骨下缘具4 个尖棘和头部顶端达到两眼后缘连线的鱼种鉴定为日本鰧;将鰧属鱼类中鱼体背部无特殊斑纹的鱼种鉴定为土佐鰧;青鰧体背侧斑点呈褐色,胸鳍基底上没有羽状皮质小突起,背鳍上仅有软条。
本研究中日本鰧的臀鳍鳍条数与《浙江海洋鱼类志》[1]757、《福建鱼类志》[5]和《东海鱼类志》[17]所描述的不一致;青鰧的胸鳍鳍条数与《浙江海洋鱼类志》[1]762和《东海鱼类志》[17]中的研究结果不一致,但与《福建鱼类志》[5]中关于青鰧的胸鳍鳍条数的描述相一致。其他关于日本鰧、土佐鰧和青鰧的形态特征结果与本研究中的形态特征结果基本一致。本研究形态特征描述结果可为鰧科鱼类形态分析、物种鉴定等工作提供补充资料。
脊椎动物的线粒体基因组中含13 个蛋白质编码基因、2 个核糖体RNA 基因、22 个转运RNA 基因以及非编码区基因[19]。线粒体基因组含有多个标志性的遗传标记位点,可根据线粒体不同部位的进化速率不同,并结合研究的情况,选择合适的遗传标记位点进行分析。以往研究者们利用线粒体CO I 基因序列对鲻科Mugilidae[20]鱼类、鳕科Gadidae[21]鱼类、金线鱼属Nemipterus[19]鱼类、银鲳Pampus argenteus[10]等开展了研究,在CO I 基因序列的碱基组成中,结果表明平均A+T 的含量高于G+C 的含量,本研究中3 种鰧科鱼类的A+T 的平均含量(50.74%)略高于G+C 的平均含量(49.26%),这与以上鱼类的CO I 基因序列研究结果一致。MEYER,et al[22]认为,理想DNA 条形码检测到的同属内种间遗传差异应该明显大于种内遗传差异,并在二者之间形成一个明显的间隔区,称作barcoding gap,在CO I 基因片段上的种内差异低于0.01,种间遗传差异大于0.02。本研究中种内遗传距离范围在0.001~0.005 之间,日本鰧与土佐鰧遗传距离最小(0.192),其次是日本鰧与青鰧(0.241),土佐鰧与青鰧遗传距离最大(0.261)。研究结果与MEYER,et al 提出的标准相符,表明3 种鰧科鱼类种类的有效性,同时验证了日本鰧与土佐鰧遗传关系更近,青鰧与其两者遗传关系较远。
HAJIBABAEI,et al[23]于2006 年将200 bp 左右的序列应用于蛾类,并且成功利用所指定的200 bp 长度的序列有效鉴定该物种。由此,HAJIBABAEI,et al[23]提出了DNA 微型条形码的概念。DNA 微型条形码技术作为一个新兴的技术,在对物种分类识别上有着其自身快捷、简单的优点。其中,MIYA,et al[14]基于大量的线粒体基因组全序列设计了180 bp 左右的高通量条形码(metabarcoding),主要用于海洋鱼类多样性监测分析。MIYA,et al 使用12S rRNA 引物对冲绳水族馆及其附近海域的水样进行鱼类多样性验证,累计准确鉴定出70 个科、152 个属,共232 种鱼类。然而,微型条形码技术又由于其发展不成熟在一定程度上受到部分学者质疑,对于微型DNA 条码的优缺点已有文章综述[24-25]。本次实验结果表明微型DNA 条形码在鰧科鱼类鉴定中具有很高的准确性。本研究使用12S rRNA 引物对3 种鰧科鱼类进行分析,结果表明种内遗传距离均为0.002,种间遗传距离范围为0.147~0.216,其中日本鰧与土佐鰧的遗传距离最小(0.147),其次是土佐鰧与青鰧(0.209),日本鰧与青鰧遗传距离最大,为0.216。本研究结果与CO I 基因序列研究结果一致,说明Metabarcoding 适用于鰧科鱼类的系统发育关系研究。综上所述,线粒体12S rRNA、CO I 基因片段可以作为DNA 条形码用于鰧科鱼类鉴定、鰧科鱼类种间系统发育关系的研究。系统发育分析结果显示日本鰧与土佐鰧亲缘关系较近,与青鰧亲缘关系较远。分子系统发育结果与基于形态学的种属划分相吻合[1]。
由于鱼类的生长环境和发育阶段的不同影响鱼类的形态特征,仅依据形态特征鉴定物种可能会存在偏差,本研究将形态学方法与DNA 条形码技术相结合,对3 种鰧科鱼类进行分析和比较,为深入展开鰧科鱼类的分类和系统发育研究奠定了基础。
猜你喜欢
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
少年文艺·开心阅读作文(2021年8期)2021-09-05
儿童时代·幸福宝宝(2020年9期)2020-09-08
中国现代中药(2019年5期)2019-07-03
学苑创造·B版(2017年10期)2017-12-21
探索科学(2017年5期)2017-06-21
电子技术与软件工程(2016年20期)2016-12-21
河北渔业(2015年10期)2015-10-15
学生天地·初中(2014年1期)2014-02-17
