芒果炭疽菌研究进展
2020-06-16 08:27:48李其利卜俊燕唐利华黄穗萍郭堂勋莫贱友
微生物学杂志 2020年1期
李其利, 卜俊燕, 唐利华, 黄穗萍, 郭堂勋, 莫贱友
(广西农业科学院植物保护研究所 广西作物病虫害生物学重点实验室,广西 南宁 530007)
芒果与香蕉、柑橘、苹果、葡萄并称世界五大水果,有着“热带果王”的美誉[1]。中国芒果主要分布在广西、海南、广东、云南、四川、福建、贵州和台湾8省区,2014年中国芒果种植面积17.32万公顷, 总产量143.77 万吨[2]。芒果炭疽病是芒果生长期和采后的重要病害之一,在世界芒果产区普遍发生[3],在芒果生长期,可以造成高于10%的损失。炭疽菌有明显的潜伏侵染现象,田间看似无病果实,常在后熟期和贮运期发病造成烂果,严重影响芒果的采后贮藏,造成的经济损失较为严重,贮运期损失一般为30%~50%,严重的高达100%[1]。本文从芒果炭疽病的症状、病原分类鉴定、生物学特性、侵染特性及致病机理等方面进行综述,为深入研究该病的发生流行规律、抗病材料的选育及该病的综合防治提供参考。
1 芒果炭疽病的症状
芒果炭疽病在叶片、花序、果实和枝梢上都可发生,且症状较为复杂。叶片感病初期出现不规则的褐色病斑,伴有黄晕。病斑可继续扩大至不规则形、圆形,呈黑褐色。多个病斑相互融合形成大病斑,甚至叶片枯死。嫩叶感病后病斑稍微突起,部分穿孔。嫩枝受害产生黑色斑点并上下扩展,严重的形成回枯症状,病斑以上部分枝叶坏死,病部出现许多小黑点。花序感病后出现黑褐色斑点,扩展形成圆形、条形斑。感病严重的花序变黑枯死,花蕾脱落,枯死[4]。未熟果实感病后,产生黑褐色小斑点。若果柄、果蒂感病,则果实很快脱落。成熟果实感病初期产生黑褐色圆形斑点,随后扩展至圆形或不规则形大病斑,中间凹陷。发病严重时病斑互相融合,果皮表面黑色。病部果肉初期变硬,后期变软腐。潮湿天气下,病部常产生淡红色分生孢子堆。此外,在台农一号、白象牙等部分品种果实上发现的红点病,最终被证实为不同症状的芒果炭疽病[5-6]。
2 芒果炭疽病病原学
2.1 芒果炭疽菌的分类鉴定
前人报道的芒果炭疽病的病原菌主要有两种,一种是无性阶段炭疽菌属胶孢炭疽菌Colletotrichumgloeosporioides(Penz.) Sace,另一种是尖孢炭疽菌ColletotrichumacutatumJ.H. Simmonds。胶孢炭疽菌属于半知菌亚门,腔孢纲,黑盘孢目,炭疽菌属(Colletotrichum),有性阶段为子囊菌门小丛壳菌Glomerellacingulata(Stenem.)Spauld. et.Schrenk,后者为小丛壳属G.acutatum。两者为害症状基本一致[7-8],其中胶孢炭疽菌C.gloeosporioides为主要病原菌[9]。
炭疽菌分类鉴定主要采用形态学鉴定和多基因分子鉴定方法相结合,形态学鉴定是最传统也最重要的区分手段。传统分类学指标有菌落培养特征,分生孢子、附着孢的形状、大小,以及刚毛形态特征,分生孢子梗及产孢细胞的形态和大小,菌核、厚垣孢子的有无和形态特征[10]。随着分子生物学技术的快速发展,Damm等[11]运用核糖体转录间隔区序列(ITS)、肌动蛋白基因(Actin gene-ACT)、几丁质合成酶A基因(chitin synthase A gene-CHS I)、3-磷酸甘油醛脱氢酶基因(glyeeraldehydes-3-phosphate dehydrogenase gene-GAPDH)、β-微管蛋白基因(β-tubulin-TUB2)和histone3(HIS3)基因进行联合建树分析,可将形态相似炭疽菌区分开来,并发现了4 个新种,Crouch等[12]将ITS、Apn2、Mat1/Apn1和Sod2基因位点进行联合分析,对形态上极难区分识别的复合群C.gramincola区分开,并引入6个新种。形态特征结合多基因系统学鉴定炭疽菌属真菌已成为发展趋势(表1)。目前用于鉴定炭疽菌的基因位点中,不同基因位点可有效区分的种类不同,其中Apmat基因位点能较好区分炭疽菌属真菌的大部分种,特别对于胶孢炭疽复合群,其单基因系统发育分析鉴定结果与ITS、ACT、CHS、CAL、GAPDH、TUB2基因位点的联合分析结果一致,可有效用于胶孢炭疽复合群的种类鉴定[13]。
根据现在的分类情况,胶胞炭疽菌是一个复合群,可分为22个种[24],而尖孢炭疽菌也是一个复合群,可分为31个种[25]。目前世界上已报道的芒果炭疽病的病原菌有12种(表2)。Lima等[23]从巴西东北部的芒果上分离出炭疽菌,联合 ACT、TUB2、CAL、GAPDH、GS、ITS六个基因分析,得到芒果炭疽菌的5个种类C.asianum、C.fructicola、C.tropicale、C.dianesei、C.karstii,其中C.dianesei是一个新种。Vieira等[26]从巴西伯南布哥州芒果中分离出炭疽菌,联合ACT、TUB2、CAL、CHS-1、GAPDH、ITS基因得到C.asianum、C.cliviae、C.dianesei、C.fructicola、C.karstii、C.tropicale、C.siamense和一个新种C.endomangiferae。Ismail等[27]联合TUB2和HIS3基因从意大利芒果中鉴定出C.karstii、C.kahawae。Calderón等[28]联合ACT、CHS-1、GAPDH、ITS、ApMat五个基因位点,得到C.asianum、C.siamense、C.theobromicola(syn.C.fragariae)。 国内报道的芒果炭疽菌种类较少,对采自攀枝花市仁和区的芒果叶斑病进行分离和纯化,采用传统形态鉴定法及ITS序列分析法对其鉴定,确定主要为胶孢炭疽菌[29];一种芒果果实上形成红点症状的病害“芒果红点病”,最初被认为是一种新病害,后来经过分离鉴定证实其为胶孢炭疽菌引起的炭疽病[5-6];广西的芒果炭疽病主要由3种炭疽菌引起,分别为亚洲炭疽菌(C.asianum)、暹罗炭疽菌(C.siamense)和果生炭疽菌(C.fructicola),其中亚洲炭疽菌为优势种群[30]。海南台农芒果的采后炭疽病病原菌主要也为这3种炭疽菌,但其优势种和各种所占比例稍有不同[29]。最近有人在广西发现C.scovillei也可引起芒果炭疽病[30]。我国芒果炭疽病的病原还未获得详细而系统的研究,一般都是对部分地区、部分分离来源(果实/叶片)的芒果炭疽病病原进行鉴定。国外已报道芒果炭疽病可由多种炭疽菌引起,我国各芒果产区生态环境差别大、品种多样且产期不一致,病原种类也可能存在显著差异。因此,有必要对我国以上八省(或自治区)不同品种、不同地理来源以及不同发病部位的炭疽病进行系统鉴定,为芒果炭疽病的抗病育种、发生流行规律和田间防治提供参考。
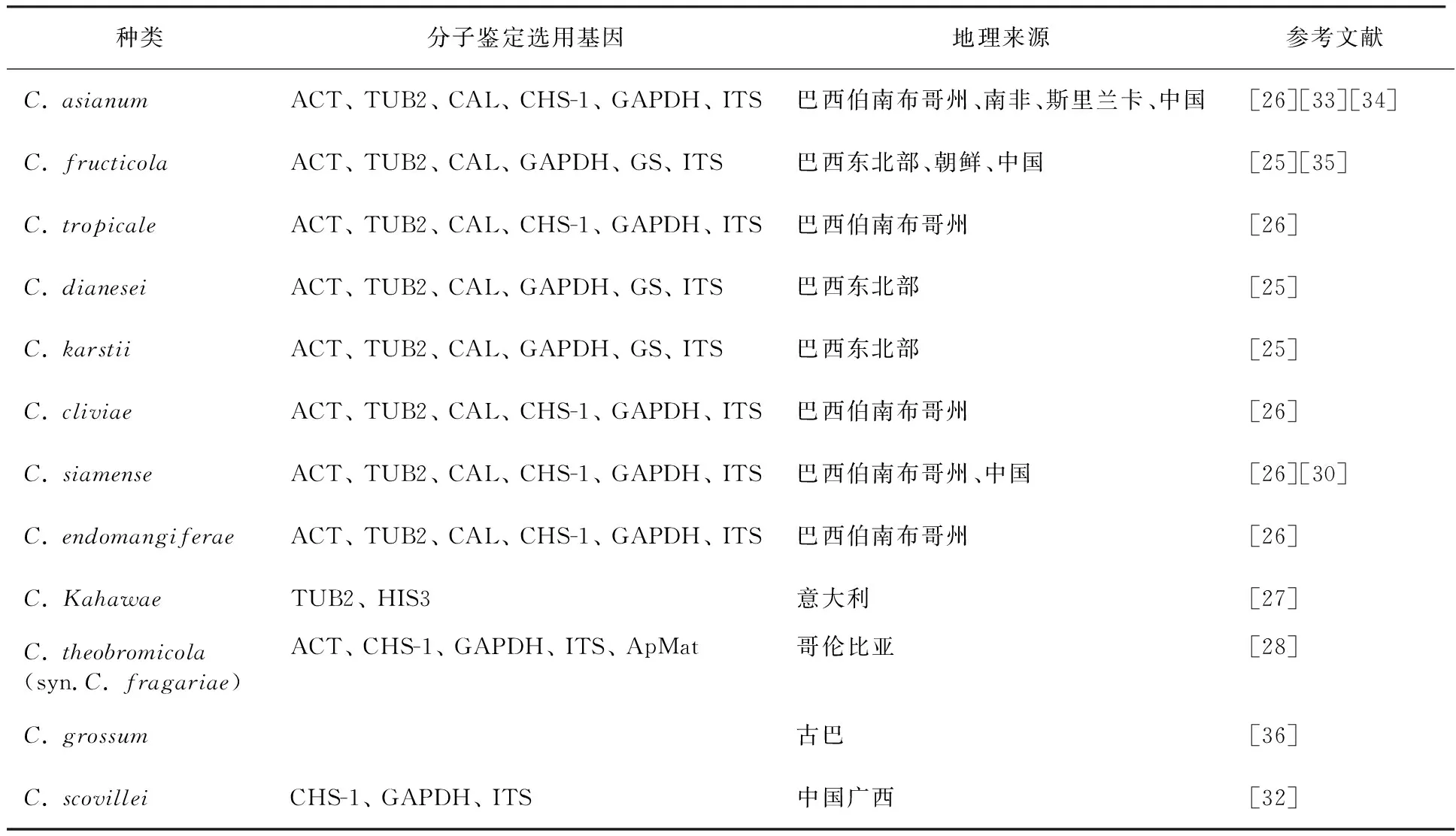
表2 国内外已报道的芒果炭疽菌种类
2.2 生物学特性
芒果炭疽菌在培养特征、菌丝生长、孢子萌发等生物学特性方面表现出种内多样性[28,37-38]。芒果胶孢炭疽菌营养生长和孢子形成的适温范围分别为20~31 ℃、25~31 ℃。pH 5.0~8.0菌丝生长较好;适宜产孢的pH值为3.5~4.5。萤光促进菌丝生长作用最强,红光和蓝光抑制菌丝生长[37]。附着胞形成的最适温度为30 ℃,相对湿度在100%+水膜的条件下分生孢子萌发较好。分生孢子萌发的最适pH值为3.0~7.0,附着胞形成的最适pH值为3.0~6.0。胡美姣等[8]将尖孢炭疽菌和胶孢炭疽菌两种芒果炭疽菌的生物特性进行比较,指出尖孢炭疽菌菌丝生长、孢子萌发及产孢的最适温度分别为26、24和28 ℃。胶孢炭疽菌丝生长、孢子萌发及产孢的最适温度分别为28、32和28 ℃,中性及近中性条件最有利于两种炭疽菌的孢子萌发和菌丝生长,pH值为3.0时有利于尖孢炭疽产孢,胶孢炭疽菌在pH值为7.0时产孢较多。含葡萄糖、蔗糖、果糖的培养基适宜于胶孢炭疽菌的菌丝生长,含淀粉的培养基利于其产生分生孢子;蔗糖、果糖有利于尖孢炭疽的菌丝生长,蔗糖和麦芽糖利于其产生分生孢子。胡美姣等[8]比较了芒果胶孢炭疽和尖孢炭疽菌两种之间菌丝生长、产孢、孢子萌发及对药剂敏感性等方面的差异,但并未比较不同温度对其病斑扩展的影响(表3),而Lima等[38]比较了温度对C.asianum、C.dianesei、C.fructicola、C.karstii和C.tropicale五种芒果炭疽菌在芒果果实上病斑扩展的影响,结果显示不同种芒果炭疽病菌最适发病温度在25~30 ℃,病斑随着温度的增加而减小。其中C.asianum和C.dianesei的最适温度分别是29.3 ℃和29.4 ℃,C.fructicola最适温度是27.1 ℃,C.karstii和C.tropicale最适温度分别是26.4、25.8 ℃。由于各芒果产区管理水平不一致,普遍存在打保险药、滥用农药的现象,因此导致芒果炭疽菌对多菌灵、苯菌灵、甲基硫菌灵、咪鲜胺等药剂的敏感性存在显著差异[39-42]。通过简单重复序列间区(Inter simple sequence repeat,ISSR)和随机扩增多态性DNA分析(Random Amplification Polymorphic DNA,RAPD)扩增多态性技术分析芒果炭疽菌遗传多样性[43-45],发现遗传多样性与芒果炭疽菌抗药性有一定相关性[43]。监测和分析芒果炭疽菌对杀菌剂的敏感性,可防止或减缓芒果炭疽菌对杀菌剂抗药性的产生,从而有效减少化学农药的用量。了解各地芒果炭疽菌的致病力强弱,有助于抗病材料的选育和抗病品种在田间的合理布局。然而,前人的研究多集中在芒果炭疽菌形态、致病力及遗传多样性等方面,很少有关于致病力变异机制的报道。
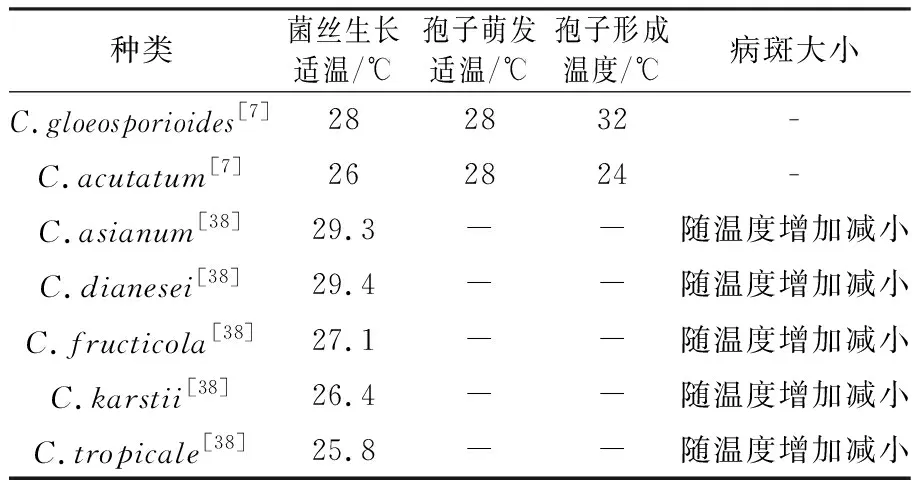
表3 不同种芒果炭疽菌生物学特性差异
注: “-”表示未测定
2.3 侵染特性
芒果炭疽病菌可在芒果枝条和烂叶等带病组织上存活两年以上[46],分生孢子盘生于芒果表皮下,成熟后冲破表皮释放分生孢子。芒果炭疽菌的分生孢子和菌丝体是该病的主要初侵染源[47],在条件适宜时形成大量的分生孢子,分生孢子进行侵染,以菌丝体在带病枝梢、僵果及枯枝上越冬[3,48]。病菌还可在病残体或果园的其他寄主上越冬,待来年气温回升,分生孢子可通过风雨、昆虫等方式在田间传播,孢子萌发后可产生附着胞,侵染钉穿透角质层并穿透表皮细胞壁侵入寄主体内,虽然炭疽菌在10 ℃环境下可引起芒果叶片产生病斑[3],但其最适温度范围在20~30 ℃。张贺等[41]研究发现,分生孢子在12~37 ℃范围内可萌发,形成附着胞的适宜温度在24~28 ℃。接种后6 h,附着胞开始形成,9~12 h是形成的高峰期;分生孢子在相对湿度90%~100%范围均可萌发且形成附着胞。相对湿度在88.4%以上最适宜病害发生,低于86%则发病率呈下降趋势[49]。附着胞在形成后10~12 h后开始黑化并逐渐变为黑褐色,这一黑褐色斑点可能是侵染钉的所在位置,或者是附着胞和植物表面组织之间的连接点[50],但Diéguez-Uribeondo等[51]认为这个点与穿透孔是随着侵染钉的形成以及病原菌在寄主细胞内的定殖生长而产生。该病菌主要侵染嫩梢、嫩叶、花序和果实,病菌以侵染丝的形式在果实表皮潜伏,侵染初期无明显症状,果实成熟或贮运期间才逐渐表现症状[52]。分生孢子产生的附着胞附着力和抗性较强,对干燥、高温、紫外线以及寄主组织表皮微生物的拮抗作用等逆境条件都具有较强的抗逆性,有利于其在寄主上长期生存。当病菌潜伏侵染到达一定程度后,是否发病主要取决于寄主植物的生理状况[53]。因此,温暖高湿、连续降雨,则病害迅速发展造成流行。目前世界上报道的芒果炭疽菌种类越来越多,不同芒果炭疽菌的侵染特性是否存在差异,需要进一步研究。
2.4 致病机理
芒果胶胞炭疽菌侵染植物组织的过程中,产生成分较为复杂的植物毒素[54-55],参与到芒果胶胞炭疽菌侵染植物组织的过程。该毒素在病原菌侵染过程中,可改变寄主植物组织细胞膜的通透性,有利于病原菌的侵入。此外,胶胞炭疽菌还会分泌果胶裂解酶、天冬氨酰蛋白酶、谷氨酸脱氢酶等一系列与致病相关的酶类,产生氨使环境pH值升高,并激发与致病相关的基因[56]。目前,已发现不同来源的芒果炭疽菌菌株对不同品种的芒果果实和叶子致病力存在显著差异[57],但其致病力变异的机理还需进一步研究。韦运谢等[58-59]对芒果胶孢炭疽菌漆酶基因Lac1和Lac2的序列特征进行探究,采用同源克隆、 SEFA-PCR 和 RT-PCR 技术对Lac1基因进行扩增,并对所获得的Lac1基因进行了序列特征和预测蛋白保守结构域的分析,也采用同源克隆和RT-PCR法获得胶孢炭疽菌漆酶基因Lac2。Lac1和Lac2基因具备真菌漆酶基因家族的序列特征,推测Lac1和Lac2基因可能与芒果炭疽菌的致病性有关。进一步研究表明漆酶基因Lac1,不仅影响致病性,而且影响病原菌的菌丝生长、分生孢子、附着胞的形成、色素沉着、黑色素生物合成、胞外水解酶的分泌及外源营养的利用等[60]。随后,采用相同的方法克隆了芒果炭疽病菌小柱孢酮脱水酶基因SCD1[61],内切葡聚糖酶基因CgEg1B等基因[62]、谷氨酸转运蛋白基因Cgglt1[63],分析其序列特征、推测了其蛋白的保守结构域,并构建了敲除载体,但其基因的功能还需进一步验证。芒果胶孢炭疽中与致病相关的基因还有很多,如何快速挖掘与致病相关基因,并研究其功能是需要进一步探讨的问题。
3 展 望
炭疽菌属真菌分类与鉴定常用的方法即传统形态学分类、分子生物学分类、次生代谢特异性等。每种分类方法都各有裨益,需采用多相分类,将形态学鉴定、多基因分子鉴定、次生代谢产物分析等方法相结合鉴定炭疽菌属真菌,从而提高分类的准确率。对于胶孢炭疽复合群(C.gloeosporioidesspecies complex),由于Apmat单基因鉴定结果与多基因联合分析鉴定结果一致,在通过ITS初步确定为胶孢炭疽菌后,可优先考虑使用Apmat进行分子鉴定。目前世界上报道的芒果炭疽病的病原菌有12种:C.asianum、C.fructicola、C.siamense、C.tropicale、C.dianesei、C.karstii、C.scovillei、C.grossum、C.cliviae、C.endomangiferae、C.kahawae、C.theobromicola(syn.C.fragariae),中国已经报道的芒果炭疽病病原有4种:C.asianum、C.fructicola、C.siamense、C.scoville。然而,前人一般都是对我国部分地区的芒果炭疽病的病原菌进行分类鉴定。因此,全面了解我国芒果炭疽病的病原种类及优势种群,明确其致病力分化情况,可为深入研究该病的发生流行规律、抗病材料的选育和抗病品种在田间的合理布局提供参考;对我国芒果炭疽病菌的抗药性进行系统分析,可防止或减缓芒果炭疽菌对杀菌剂抗药性的产生,从而有效减少化学农药的用量,也是研究芒果炭疽病绿色防控新策略、新方法和新药剂的基础。中国不同地区芒果炭疽病病原菌是否只有已报道的4种,其优势病原菌是否一致,这些不同种炭疽菌在生物学特性、侵染特性等方面是否存在差异?芒果炭疽病菌有潜伏侵染特性,如何快速检测芒果炭疽病菌不同种类,并研究其带菌情况与病害发生的关系,可为该病防治提供参考。中国不同地理来源的芒果炭疽菌致病力存在较大差异,其致病力变异与地理来源是否存在关联,致病力分化及扩散的分子机制如何?都是需要进一步研究的问题。此外,国内外关于芒果炭疽菌致病机理的报道较少,今后需要进一步加强其致病机理研究,为芒果炭疽病的绿色防控提供参考。
猜你喜欢
今日农业(2022年15期)2022-09-20 06:54:16
中国森林病虫(2021年6期)2021-12-20 08:45:42
现代畜牧科技(2021年4期)2021-07-21 06:13:34
小学生必读(中年级版)(2021年12期)2021-03-03 07:43:44
小学生必读(中年级版)(2021年11期)2021-02-22 05:19:06
宁夏医学杂志(2020年4期)2021-01-21 08:25:16
中国森林病虫(2018年4期)2018-09-19 12:13:08
衡阳师范学院学报(2016年3期)2016-07-10 07:16:27
塔里木大学学报(2015年1期)2015-04-25 02:38:44
营销界(2015年23期)2015-02-28 22:06:16