
山东鹊山省级自然保护区胡桃楸的居群结构及遗传多样性分析
2020-06-12 10:58胡德昌解孝满王仲礼
植物资源与环境学报 2020年3期
张 浩,胡德昌,①,解孝满,刘 丹,张 萍,王仲礼
(1.鲁东大学生命科学学院,山东 烟台 264025;2.山东省林木种质资源中心,山东 济南 250014)
胡桃楸(JuglansmandshuricaMaxim.)为落叶乔木,具有耐寒和抗风等优良特性,经济价值较高。由于栖息地受到不同程度破坏,生境退化或丧失,其居群数量急剧减少,存在濒危风险,目前在中国珍稀濒危植物信息系统(ISCREP)(http:∥www.iplant.cn/rep/)中该种已被列为吉林、河北和北京的地方保护野生植物。通过近5年的调查,作者发现在胶东半岛胡桃楸野生居群数量较少,呈现片断化分布特点,人为采挖和生境变化等问题导致胡桃楸野生居群减少甚至消失的风险,因而亟需加大其野生资源的保护力度。
已有研究者对新疆、河北及东北等地的胡桃属种类开展了居群特征和遗传多样性等研究[1-3],[4]19-33。王东升等[5]基于AFLP标记对山东分布的5个胡桃属群体进行了遗传多样性分析,发现这5个群体的遗传关系与其地理分布不完全一致。作者的实地调查结果显示:山东地区生长的胡桃楸个体的叶片和果型等部分生物学性状存在差异,但这些性状的遗传变异程度及其遗传关系尚不清楚,其个体变异及居群规模减小的原因有待分析和探讨。
作者在前期调查的基础上对山东鹊山省级自然保护区不同坡位样方进行居群生态学调查,并通过径级结构、静态生命表、期望寿命曲线、存活曲线和死亡率曲线探讨胡桃楸的居群结构和特征,并采用SRAP分子标记探讨胡桃楸个体间的遗传关系,以期为胡桃楸的居群结构变化、遗传变异和致危因子研究以及胡桃楸保护策略的制定奠定基础。
1 研究区概况和研究方法
1.1 研究区概况
鹊山省级自然保护区在山东半岛中北部(东经121°16′39″~121°24′21″、北纬37°07′47″~37°14′57″),总面积约1 485.2 hm2;属长白山系崂山山脉,主峰北垛海拔653.8 m,为典型的胶东低山丘陵地貌。该研究区属暖温带季风型大陆性气候,四季分明、季风显著,雨热同期、空气湿润,温差较小、光照充足;年平均气温11.6 ℃,年均降水量736.6 mm,年均蒸发量1 693.2 mm,空气相对湿度71%。土壤以棕壤为主。
该保护区林地面积1 358.5 hm2,研究区内植被既有暖温带阔叶林特征,又有温带针阔混交林特征,主要为赤松(PinusdensifloraSieb.et Zucc.)纯林(640.4 hm2)和赤松-麻栎(QuercusacutissimaCarruth.)混交林(475.5 hm2)。研究区内植物垂直特征明显,海拔300 m以下多为农田、果林、灌丛和落叶阔叶林,海拔300 m以上则多为针阔混交林和温带针叶林。
据不完全统计,该保护区共有维管植物115科435属786种,包括国家重点保护野生植物野大豆(GlycinesojaSieb.et Zucc.)和中华结缕草(ZoysiasinicaHance),山东珍稀植物长冬草〔Clematishexapetalavar.tchefouensis(Debeaux)S.Y.Hu〕、宽蕊地榆(SanguisorbaapplanataYu et Li)、滨海前胡〔Peucedanumwawrae(Wolff)Su〕、泰山韭(AlliumtaishanenseJ.M.Xu)、坚桦(BetulachinensisMaxim.)和迎红杜鹃(RhododendronmucronulatumTurcz.)等[6]。
1.2 研究方法
1.2.1 样地设置和样品采集 于2016年5月至9月,采用样方法在该保护区内设置东、西2条样线,并在东样线划分2个样方,在西样线划分4个样方,共6个样方;样方间距均大于500 m,样方面积20 m×20 m。根据海拔,将6个样方分为山体上部、中部和下部3组不同坡位的样地。其中,山体下部样地仅西山采石场(P1)1个样方,山体中部样地包含高顶后(P2)、黑石夼西沟东大梁(P3)和黑石里沟(P4)3个样方,山体上部样地包含黄泥洼(P5)和西山上部(P6)2个样方,基本涵盖了该保护区内胡桃楸的所有片状分布点。6个胡桃楸样方的基本信息见表1。
表1 山东鹊山省级自然保护区6个胡桃楸样方的基本信息
Table 1 Basic informations of six quadrats ofJuglansmandshuricaMaxim.in Queshan Provincial Nature Reserve of Shandong
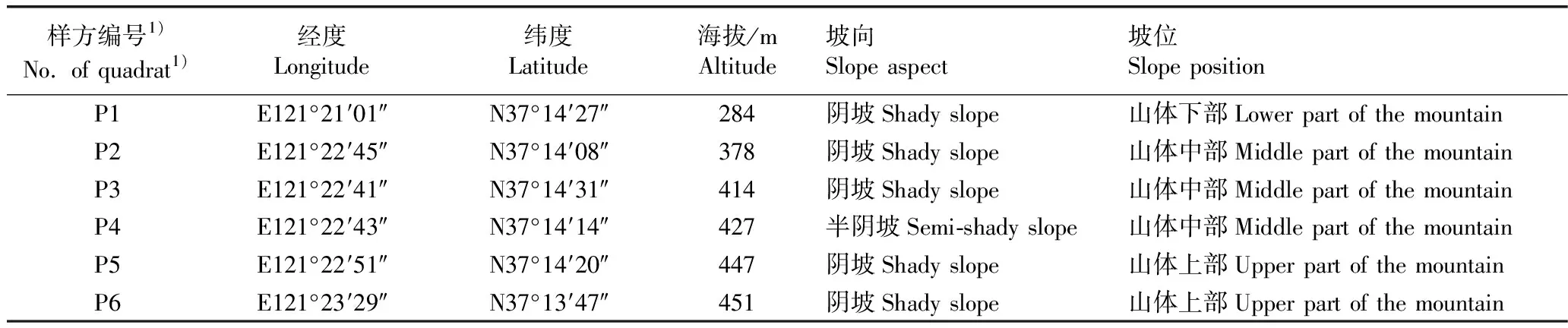
样方编号1)No. of quadrat1)经度Longitude纬度Latitude海拔/mAltitude坡向Slope aspect坡位Slope positionP1E121°21′01″N37°14′27″284阴坡Shady slope山体下部Lower part of the mountainP2E121°22′45″N37°14′08″378阴坡Shady slope山体中部Middle part of the mountainP3E121°22′41″N37°14′31″414阴坡Shady slope山体中部Middle part of the mountainP4E121°22′43″N37°14′14″427半阴坡Semi-shady slope山体中部Middle part of the mountainP5E121°22′51″N37°14′20″447阴坡Shady slope山体上部Upper part of the mountainP6E121°23′29″N37°13′47″451阴坡Shady slope山体上部Upper part of the mountain
1)P1:西山采石场Quarry of Xishan Hill;P2:高顶后Gaodinghou;P3:黑石夼西沟东大梁Dongdaliang in Xigou of Heishikuang;P4:黑石里沟Heishiligou;P5:黄泥洼Huangniwa;P6:西山上部Upper part of Xishan Hill.
于2016年春季在该保护区的西山采石场(E121°21′01″、N37°14′27″)、椴树洼(E121°22′26″、N37°13′40″)、狼夼(E121°23′18″、N37°13′52″)、高顶后(E121°22′45″、N37°14′08″)、黑石里沟(E121°22′42″、N37°14′14″)、虎龙头(E121°22′49″、N37°15′02″)和宋山(E121°22′51″、N37°14′20″)采集9份胡桃楸样本,前6个采样点各采集样本1份,最后1个采样点采集3份样本,依次编号1~9;另在西山采石场采集1份栽培胡桃(J.regiaLinn.)样本,作为外类群,编号10。采样点的确定综合考虑了胡桃楸的分布状况以及地理位置、株龄和生长状况等因子,且为了避免克隆单株,样株间隔大于30 m。在各样株的中部枝条上分别采集完整无斑的叶片3~5枚,硅胶干燥,供试。
1.2.2 居群生态学调查和居群径级结构分析 记录2条样线的生境条件、坡度、坡向、坡位和土壤类型,以及植被的种类、密度和盖度。对6个样方内的所有胡桃楸进行每木调查,记录个体数和生长状况等基本信息。对于成年树,记录其胸径、株高和冠幅,胸径用围尺(精度1 mm)测量,株高用测高仪(精度10 cm)测量,冠幅用直尺(精度1 mm)测量;对于幼苗和幼树,记录基径和株高,基径用游标卡尺(精度0.1 mm)测量,株高用测高仪测量。
参照文献[7],根据胸径(DBH)将胡桃楸划分为7个径级:Ⅰ级(DBH<2 cm)、Ⅱ级(2 cm≤DBH<5 cm)、Ⅲ级(5 cm≤DBH<10 cm)、Ⅳ级(10 cm≤DBH<15 cm)、Ⅴ级(15 cm≤DBH<20 cm)、Ⅵ级(20 cm≤DBH<25 cm)和Ⅶ级(25 cm≤DBH<30 cm),其中,Ⅰ级和Ⅱ级为幼苗和幼树,Ⅲ级至Ⅶ级为成年树。以各样方胡桃楸的径级为横坐标、个体数为纵坐标绘制径级结构图。
1.2.3 静态生命表编制及曲线绘制 采用径级代替龄级的方法编制不同坡位样地胡桃楸的静态生命表以及胡桃楸居群的整体静态生命表[7]。
在静态生命表编制过程中,部分样方中胡桃楸的死亡率呈现负值,因而,参考江洪[8]和张维等[1]的数据分析方法,采用匀滑处理进行校正。静态生命表编制完成后,以径级为横坐标,期望寿命(ex)、标准化存活数(lx)和死亡率(qx)为纵坐标,分别绘制各样地胡桃楸的期望寿命曲线、存活曲线和死亡率曲线。
1.2.4 SRAP分子标记分析 采用CTAB[9]法提取叶片DNA并进行质量检测,-20 ℃保存。根据文献[10]中的SRAP引物序列及扩增程序,采用BIO-RAD伯乐S1000梯度PCR仪(美国Bio-Rad公司)进行PCR扩增,从77对SRAP引物中,筛选出14对扩增结果稳定、谱带清晰、多态性丰富的引物(M1E3、M1E11、M1E6、M2E2、M2E5、M3E9、M4E2、M4E8、M5E10、M6E6、M6E7、M7E9、M7E10和M7E11)用于SRAP扩增,扩增产物经质量体积分数2%琼脂糖凝胶电泳检测,用Tanon 2500R凝胶成像系统(上海天能科技有限公司)观察并记录谱带。
1.3 数据处理和分析
采用EXCEL 2007软件处理数据。根据SRAP扩增条带“有”或“无”生成“1”、“0”原始矩阵,采用NTSYSpc2.10软件计算遗传相似系数,采用UPGMA法进行聚类分析并绘制树状图。
2 结果和分析
2.1 胡桃楸居群的径级结构分析
山东鹊山省级自然保护区6个胡桃楸样方的径级结构见图1;胡桃楸居群的径级结构见图2。
由图1可见:P2样方(高顶后)内个体数最少,占6个样方个体总数的8.1%;P3样方(黑石夼西沟东大梁)内个体数最多,占6个样方个体总数的32.6%。位于山体下部的P1样方(西山采石场)中,Ⅰ级(DBH<2 cm)至Ⅲ级(5 cm≤DBH<10 cm)的个体数为20,其中Ⅰ级和Ⅱ级(2 cm≤DBH<5 cm)的个体数占该样方个体总数的45.0%,Ⅳ级(10 cm≤DBH<15 cm)及以上径级个体缺失,该样方内胡桃楸整体表现为增长型。在山体中部及上部样方中,位于山体中部的P3样方缺失Ⅵ级(20 cm≤DBH<25 cm)和Ⅶ级(25 cm≤DBH<30 cm)个体,位于山体中部的P4样方(黑石里沟)和位于山体上部的P5样方(黄泥洼)缺失Ⅶ级个体,位于山体中部的P2样方和位于上体上部的P6样方(西山上部)内各径级个体均有分布;其中,各样方中Ⅰ级至Ⅳ级的个体数均高于Ⅴ级(15 cm≤DBH<20 cm)至Ⅶ级的个体数,这5个样方的胡桃楸整体表现为稳定型。
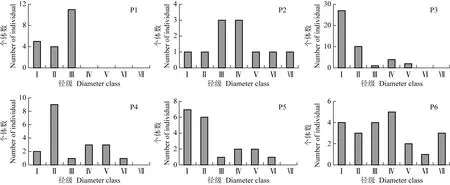
Ⅰ:DBH<2 cm;Ⅱ:2 cm≤DBH<5 cm;Ⅲ:5 cm≤DBH<10 cm;Ⅳ:10 cm≤DBH<15 cm;Ⅴ:15 cm≤DBH<20 cm;Ⅵ:20 cm≤DBH<25 cm;Ⅶ:25 cm≤DBH<30 cm.DBH:胸径Diameter at breast height.
P1:西山采石场(山体下部样地)Quarry of Xishan Hill(plot at lower part of the mountain);P2,P3,P4:分别为高顶后、黑石夼西沟东大梁和黑石里沟(山体中部样地)Gaodinghou,Dongdaliang in Xigou of Heishikuang and Heishiligou,respectively(plot at middle part of the mountain);P5,P6:分别为黄泥洼和西山上部(山体上部样地)Huangniwa and upper part of Xishan Hill,respectively(plot at upper part of the mountain).
图1 山东鹊山省级自然保护区6个胡桃楸样方的径级结构
Fig.1 Diameter class structure of six quadrats ofJuglansmandshuricaMaxim.in Queshan Provincial Nature Reserve of Shandong
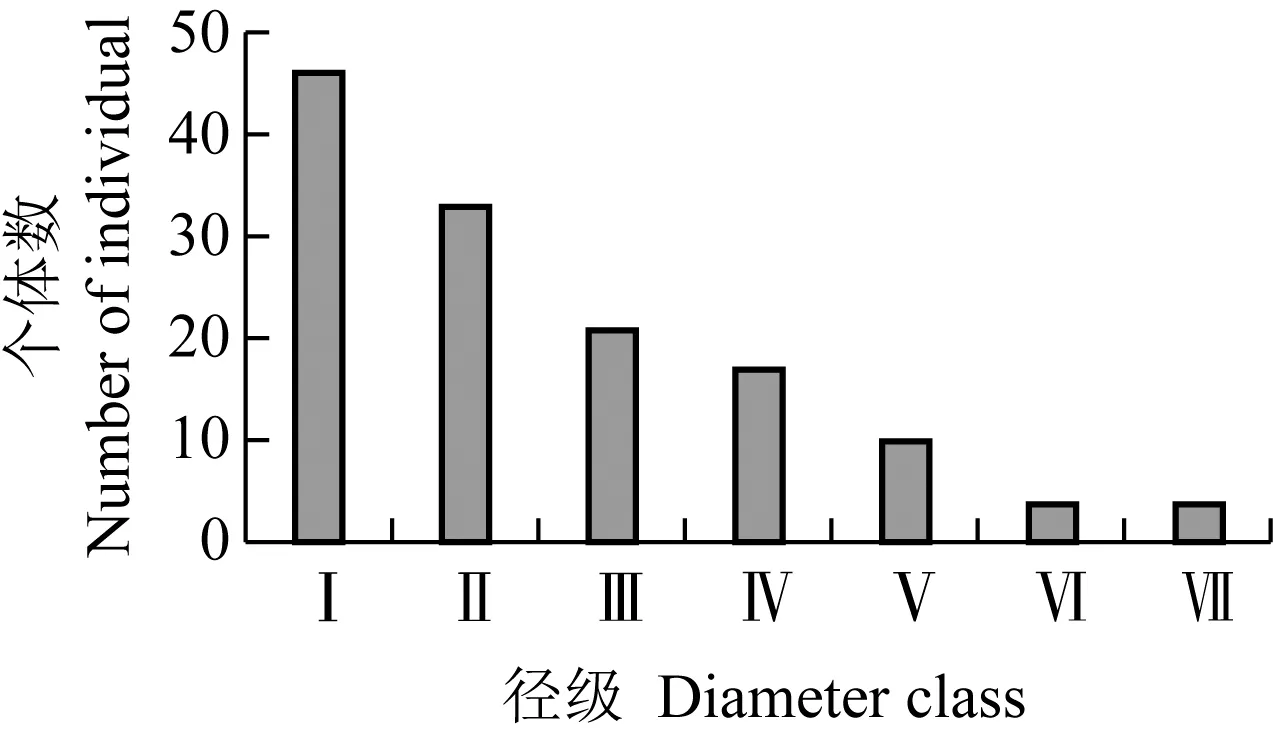
Ⅰ:DBH<2 cm;Ⅱ:2 cm≤DBH<5 cm;Ⅲ:5 cm≤DBH<10 cm;Ⅳ:10 cm≤DBH<15 cm;Ⅴ:15 cm≤DBH<20 cm;Ⅵ:20 cm≤DBH<25 cm;Ⅶ:25 cm≤DBH<30 cm.DBH:胸径Diameter at breast height.
图2 山东鹊山省级自然保护区胡桃楸居群的径级结构
Fig.2 Diameter class structure ofJuglansmandshuricaMaxim.population in Queshan Provincial Nature Reserve of Shandong
由图2可见:6个样方的胡桃楸个体总数为135株,Ⅰ级至Ⅶ级的个体数分别占个体总数的34.1%、24.4%、15.5%、12.6%、7.4%、3.0%和3.0%。其中,Ⅰ级和Ⅱ级的个体数占个体总数的58.5%,而Ⅲ级至Ⅶ级的个体数占个体总数的41.5%,其中Ⅵ级和Ⅶ级的个体数占个体总数的6.0%,表现出增长型居群的径级结构特征。
2.2 胡桃楸居群静态生命表及生命过程分析
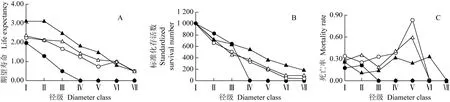
山东鹊山省级自然保护区胡桃楸居群和居群内不同样地的静态生命表见表2;胡桃楸的期望寿命曲线、存活曲线和死亡率曲线见图3。
由表2可见:山体上部、中部和下部样地内以及整个居群中不同径级胡桃楸个体存活数(ax)总体变化较平稳,从低径级到高径级依次减少,但山体中部样地的胡桃楸个体存活数波动较大,表明在大多数情况下,核桃楸个体可以通过环境的筛选进入下一阶段的生长发育,以完成居群的天然更新。山体上部、中部和下部样地内以及整个居群中不同径级胡桃楸个体的期望寿命(ex)总体均随径级增加而逐级递减,而死亡率(qx)总体随径级增加小幅波动,仅山体中部样地的V级(15 cm≤DBH<20 cm)胡桃楸个体的死亡率最高(0.835),导致整个居群该径级的胡桃楸个体死亡率也较高(0.599),表明胡桃楸在该阶段由中等大树向大树过渡,是其生活史中较为脆弱的阶段。
由图3-A可见:胡桃楸居群和居群内不同样地中胡桃楸个体的ex值总体上均随径级增加而逐渐降低,仅山体中部样地中Ⅴ级和Ⅵ级(20 cm≤DBH<25 cm)的ex值有小幅波动。小径级个体的ex值均高于大径级个体,山体中部和上部样地的胡桃楸个体的ex值明显高于山体下部样地,且胡桃楸整个居群的ex值总体上高于山体下部和中部样地,但低于山体上部样地。
由图3-B可见:胡桃楸整个居群的存活曲线呈直线型,根据Deevey[11]的划分标准,该胡桃楸居群的存活曲线基本符合Deevey Ⅱ型。山体上部、中部和下部样地的胡桃楸存活曲线分别属于直线型、直线型和凹型,其中,山体中部和上部样地的胡桃楸存活曲线均符合Deevey Ⅱ型,随径级增加,个体标准化存活数逐渐下降;而山体下部样地的胡桃楸存活曲线则接近Deevey Ⅲ型,前期干扰因子导致Ⅳ级(10 cm≤DBH<15 cm)及以上个体标准化存活数急剧减少。
表2 山东鹊山省级自然保护区胡桃楸居群和居群内不同样地的静态生命表1)
Table 2 Static life tables of population and different plots in population ofJuglansmandshuricaMaxim.in Queshan Provincial Nature Reserve of Shandong1)
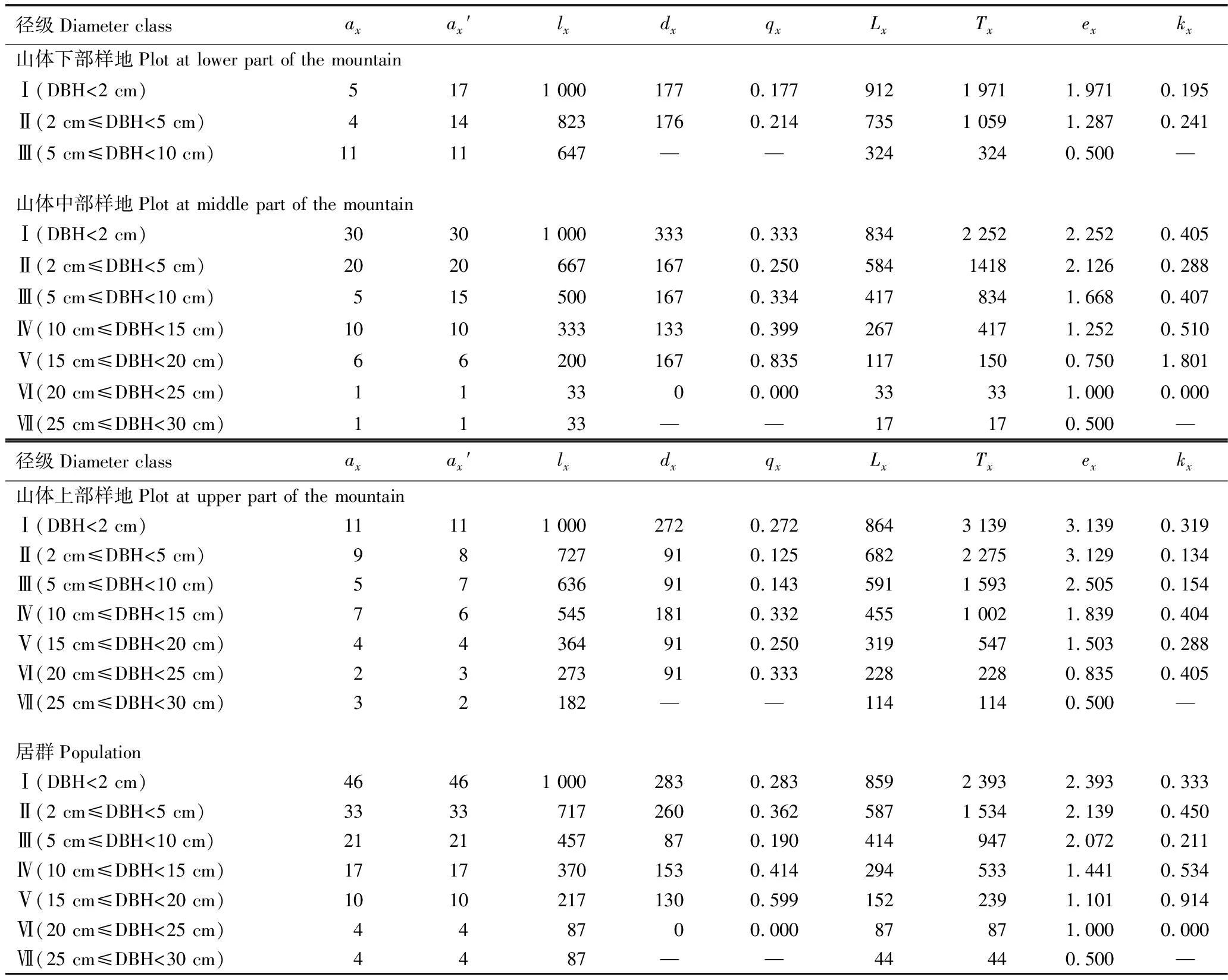
径级Diameter classaxax′lxdxqxLxTxexkx山体下部样地Plot at lower part of the mountainⅠ(DBH<2 cm)5171 0001770.1779121 9711.9710.195Ⅱ(2 cm≤DBH<5 cm)4148231760.2147351 0591.2870.241Ⅲ(5 cm≤DBH<10 cm)1111647——3243240.500—山体中部样地Plot at middle part of the mountainⅠ(DBH<2 cm)30301 0003330.3338342 2522.2520.405Ⅱ(2 cm≤DBH<5 cm)20206671670.25058414182.1260.288Ⅲ(5 cm≤DBH<10 cm)5155001670.3344178341.6680.407Ⅳ(10 cm≤DBH<15 cm)10103331330.3992674171.2520.510Ⅴ(15 cm≤DBH<20 cm)662001670.8351171500.7501.801Ⅵ(20 cm≤DBH<25 cm)113300.00033331.0000.000Ⅶ(25 cm≤DBH<30 cm)1133——17170.500—径级Diameter classaxax′lxdxqxLxTxexkx山体上部样地Plot at upper part of the mountainⅠ(DBH<2 cm)11111 0002720.2728643 1393.1390.319Ⅱ(2 cm≤DBH<5 cm)98727910.1256822 2753.1290.134Ⅲ(5 cm≤DBH<10 cm)57636910.1435911 5932.5050.154Ⅳ(10 cm≤DBH<15 cm)765451810.3324551 0021.8390.404Ⅴ(15 cm≤DBH<20 cm)44364910.2503195471.5030.288Ⅵ(20 cm≤DBH<25 cm)23273910.3332282280.8350.405Ⅶ(25 cm≤DBH<30 cm)32182——1141140.500—居群PopulationⅠ(DBH<2 cm)46461 0002830.2838592 3932.3930.333Ⅱ(2 cm≤DBH<5 cm)33337172600.3625871 5342.1390.450Ⅲ(5 cm≤DBH<10 cm)2121457870.1904149472.0720.211Ⅳ(10 cm≤DBH<15 cm)17173701530.4142945331.4410.534Ⅴ(15 cm≤DBH<20 cm)10102171300.5991522391.1010.914Ⅵ(20 cm≤DBH<25 cm)448700.00087871.0000.000Ⅶ(25 cm≤DBH<30 cm)4487——44440.500—
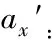
Ⅰ:DBH<2 cm;Ⅱ:2 cm≤DBH<5 cm;Ⅲ:5 cm≤DBH<10 cm;Ⅳ:10 cm≤DBH<15 cm;Ⅴ:15 cm≤DBH<20 cm;Ⅵ:20 cm≤DBH<25 cm;Ⅶ:25 cm≤DBH<30 cm.DBH:胸径Diameter at breast height.
图3 山东鹊山省级自然保护区胡桃楸的居群和居群内不同样地的期望寿命曲线(A)、存活曲线(B)和死亡率曲线(C)
Fig.3 Life expectancy curve(A),survival curve(B)and mortality rate curve(C)of population and different plots in the population ofJuglansmandshuricaMaxim.in Queshan Provincial Nature Reserve of Shandong
由图3-C可见:在山体中部样地以及整个居群中,胡桃楸个体死亡率随径级增加呈逐渐升高然后降低的趋势,并均在Ⅴ级达到峰值,死亡率分别为0.835和0.599,而山体上部样地的胡桃楸个体死亡率随径级增加小幅波动。总体上看,山体中部样地的胡桃楸个体死亡率相对偏高。从Ⅰ级(DBH<2 cm)和Ⅱ级(2 cm≤DBH<5 cm)以及从Ⅴ级和Ⅵ级,胡桃楸个体的生长易受人为干扰、环境筛选或种内竞争等因子影响,生存压力较大;Ⅲ级(5 cm≤DBH<10 cm)的个体死亡率较低,说明此径级的胡桃楸存活能力较强。
2.3 胡桃楸SRAP分子标记和遗传关系分析
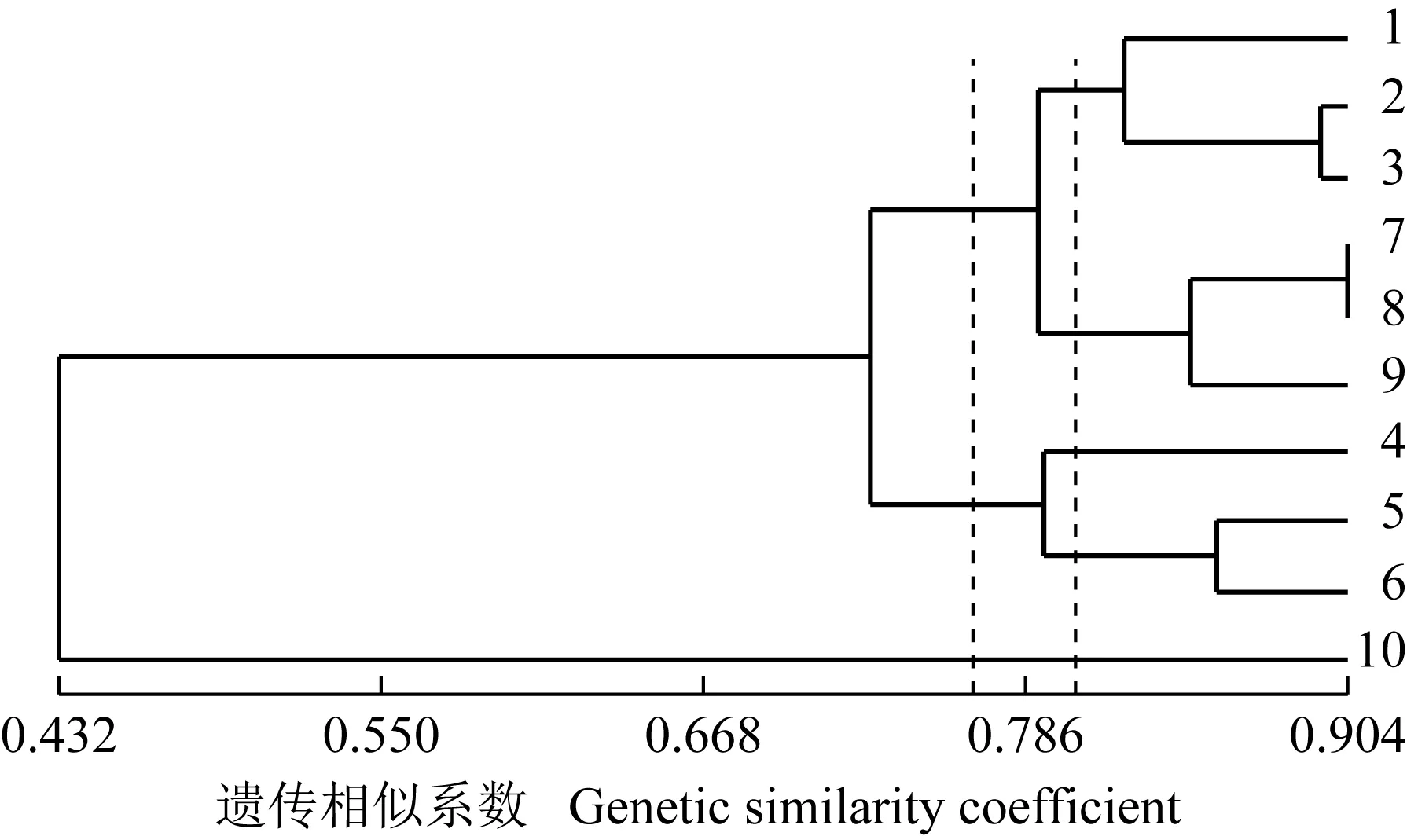
采用14对SRAP引物,对采自山东鹊山省级自然保护区的9份胡桃楸样本及1份外类群样本(胡桃栽培样本)进行遗传分析,其中4对引物M2E5、M3E9、M6E7和M1E3的SRAP扩增图谱见图4。依据SRAP分子标记分析结果计算供试样本的遗传相似系数,结果见表3;依据表3的结果,采用UPGMA法进行聚类分析,结果见图5。
2.3.1 遗传多样性和遗传相似性 SRAP分子标记分析结果表明:10份样本中共扩增104个位点,其中遗传多态性位点占49.0%。9份胡桃楸样本的多态性位点数差异较大,其中,采自狼夼的3号样本多态性位点最多(41),采自黑石里沟的5号样本多态性位点最少(11)。在引物M2E5扩增图谱的280 bp位点(图4-A),采自狼夼的3号样本和采自宋山的8号样本差异明显;在引物M6E7扩增图谱的350和400 bp位点(图4-C),不同样本差异明显。
由表3可见:9份胡桃楸样本间遗传相似系数均较高,平均值为0.786;其中,采自宋山的7号和8号样本间的遗传相似系数最大,达到0.904;采自狼夼的3号与采自黑石里沟的5号样本间以及采自高顶后的4号与采自宋山的7号样本间的遗传相似系数最小,均为0.654,表明胡桃楸居群内个体间的遗传变异较为丰富。而9份胡桃楸样本与栽培胡桃(外类群)间的遗传相似系数为0.394~0.510,表明胡桃楸与胡桃的种间遗传差异较大。
2.3.2 遗传关系 聚类分析结果(图5)显示:在遗传相似系数0.762处,供试的10份样本分为3组,其中,栽培胡桃单独成为第1组,9份胡桃楸样本聚为第2组和第3组。第2组包括4号、5号和6号样本;第3组由其余6份样本聚成。在遗传相似系数0.804处,还可将第3组分为2个亚组,Ⅰ亚组包括采自西山采石场的1号样本、采自椴树洼的2号样本和采自狼夼的3号样本,Ⅱ亚组则由采自宋山的7号、8号和9号样本聚成。
在I亚组中,2号和3号样本间的遗传关系较近,二者分布于相邻的沟谷中;而1号样本与其他样本间的遗传关系较远,且采自该保护区最西侧(西山采石场),地理距离相对较远。在Ⅱ亚组中,3份样本间的遗传相似系数为0.827~0.904,均采自同一区域,地理分布相对集中。说明胡桃楸各样本间的遗传关系与其地理分布有一定的关联。
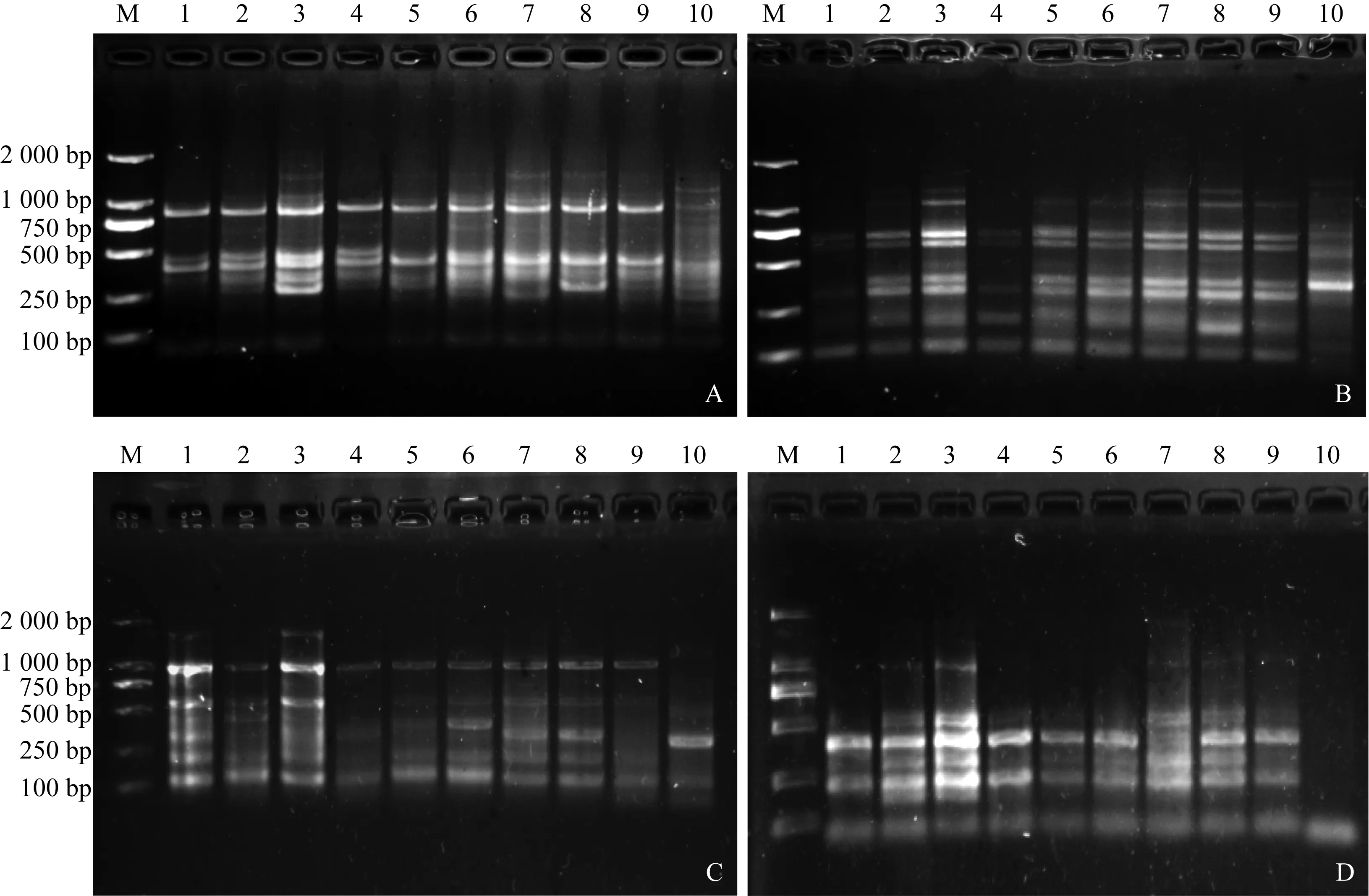
A,B,C,D:分别为引物M2E5、M3E9、M6E7和M1E3的扩增图谱Being amplification patterns of primers M2E5,M3E9,M6E7 and M1E3,respectively.
M:DNA marker.1-10:供试样本 Samples tested:1.采自西山采石场Collected from Quarry of Xishan Hill;2.采自椴树洼Collected from Duanshuwa;3.采自狼夼Collected from Langkuang;4.采自高顶后Collected from Gaodinghou;5.采自黑石里沟Collected from Heishiligou;6.采自虎龙头Collected from Hulongtou;7,8,9.采自宋山Collected from Songshan;10.栽培胡桃(外类群)CultivatedJ.regiaLinn.(outgroup).
图4 山东鹊山省级自然保护区胡桃楸不同样本的SRAP扩增图谱
Fig.4 SRAP amplification patterns of different samples ofJuglansmandshuricaMaxim.in Queshan Provincial Nature Reserve of Shandong
表3 山东鹊山省级自然保护区胡桃楸不同样本间的遗传相似系数1)
Table 3 Genetic similarity coefficient among different samples ofJuglansmandshuricaMaxim.in Queshan Provincial Nature Reserve of Shandong1)
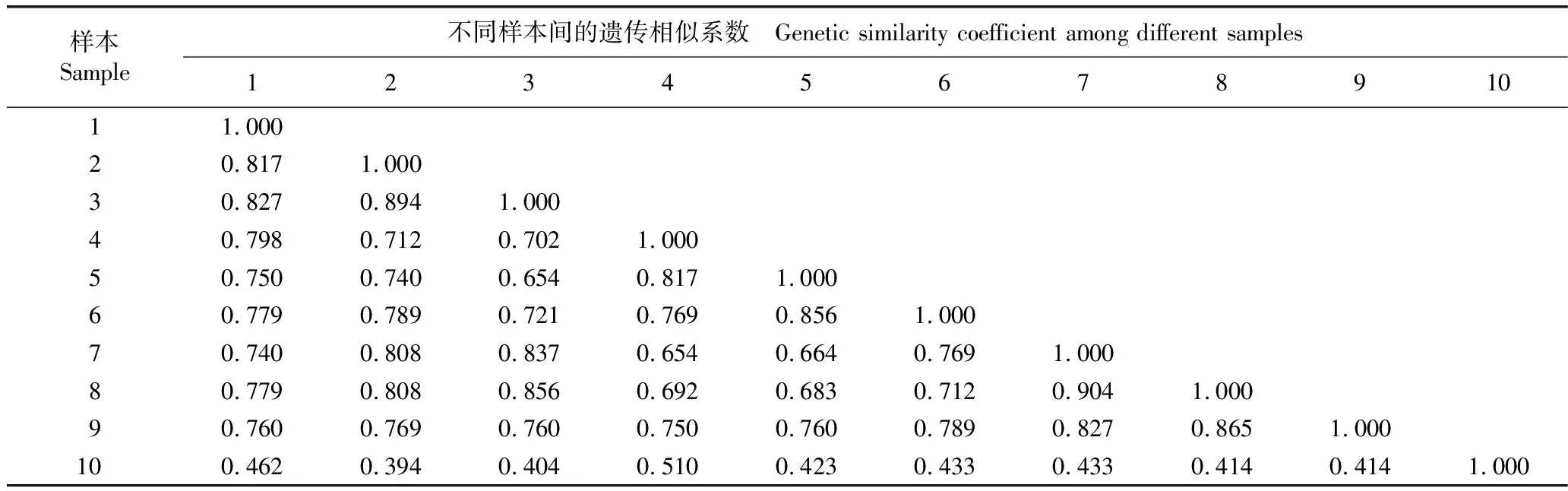
样本Sample不同样本间的遗传相似系数 Genetic similarity coefficient among different samples1234567891011.00020.8171.00030.8270.8941.00040.7980.7120.7021.00050.7500.7400.6540.8171.00060.7790.7890.7210.7690.8561.00070.7400.8080.8370.6540.6640.7691.00080.7790.8080.8560.6920.6830.7120.9041.00090.7600.7690.7600.7500.7600.7890.8270.8651.000100.4620.3940.4040.5100.4230.4330.4330.4140.4141.000
1)1-10:供试样品 Samples tested:1.采自西山采石场Collected from Quarry of Xishan Hill;2.采自椴树洼Collected from Duanshuwa;3.采自狼夼Collected from Langkuang;4.采自高顶后Collected from Gaodinghou;5.采自黑石里沟Collected from Heishiligou;6.采自虎龙头Collected from Hulongtou;7,8,9.采自宋山Collected from Songshan;10.栽培胡桃(外类群)CultivatedJ.regiaLinn.(outgroup).
1-10:供试样品 Samples tested:1.采自西山采石场Collected from Quarry of Xishan Hill;2.采自椴树洼Collected from Duanshuwa;3.采自狼夼Collected from Langkuang;4.采自高顶后Collected from Gaodinghou;5.采自黑石里沟Collected from Heishiligou;6.采自虎龙头Collected from Hulongtou;7,8,9.采自宋山Collected from Songshan;10.栽培胡桃(外类群)CultivatedJ.regiaLinn.(outgroup).
图5 山东鹊山省级自然保护区胡桃楸不同样本的UPGMA聚类图
Fig.5 UPGMA clustering dendrogram of different samples ofJuglansmandshuricaMaxim.in Queshan Provincial Nature Reserve of Shandong
3 讨论和结论
野生濒危植物渐少的主要原因是天然植被屡遭破坏、生境恶化[12-14]。调查结果显示:山东鹊山省级自然保护区内的胡桃楸居群呈现片状分布,其中,山体下部样地中Ⅳ级(10 cm≤DBH<15 cm)及以上的个体缺失,而中部和上部样地的径级结构相对完整,表明该居群的实生繁殖受生存环境影响较大。调查区域内Ⅲ级(5 cm≤DBH<10 cm)及以上径级的胡桃楸成年树共56株,最大胸径为27.2 cm;胡桃楸幼苗和幼树数量较多(共79株,平均每个样方13株),但各样方间幼苗和幼树的数量差异较大,有部分样方内的幼苗和幼树相对较少,如P1样方(西山采石场)仅5株、P2样方(高顶后)仅1株、P4样方(黑石里沟)仅2株。造成这种现象的原因主要有2个方面:一方面是胡桃楸结实率较低,且果实易被采食,可萌发的种子数量较少;另一方面则是生境条件的影响,胡桃楸多生长于沟底乱石间,土壤贫瘠,只有少量落在裸岩间潮湿土壤中的成熟种子能顺利萌发,且只有少量幼苗能存活。
在鹊山省级自然保护区山体下部样地中,Ⅳ级及以上的胡桃楸个体缺失,且Ⅱ级(2 cm≤DBH<5 cm)的个体死亡率高于Ⅰ级(DBH<2 cm),说明至少在5年前山体下部样地的胡桃楸幼苗可能因环境影响和人为干扰而难以存活;山体中部样地受人为干扰较少、生境较为稳定,前期各径级胡桃楸个体死亡率相对平稳,而Ⅴ级(15 cm≤DBH<20 cm)的个体死亡率最高;山体上部样地胡桃楸个体死亡率总体较低,与其受人为干扰较少有关。从Ⅲ级开始,胡桃楸个体数明显减少,表明该胡桃楸居群在发育初期自然繁殖能力相对正常,幼苗和幼树较为丰富,然后以高死亡率为代价从幼龄植株发育为成年植株,进而完成整个生活史;而胡桃楸成年个体的适应能力较强,其生长主要受种内竞争和自疏作用的影响。
由于群落处于不同的演替阶段,分布于不同地区、不同坡位的植物可能呈现不同的分布格局[15-16]。在鹊山省级自然保护区不同坡位的样地中,山体中部和上部样地胡桃楸个体的生存状况明显优于山体下部样地,其存活曲线符合Deevey Ⅱ型,而山体下部样地胡桃楸存活曲线则接近Deevey Ⅲ型。闫娜等[3]的研究结果表明:雾灵山国家级自然保护区胡桃楸居群中幼树和中龄树所占比例较大,存活曲线接近Deevey Ⅲ型。造成这种差异的原因可能有2个:一是研究区域的自然环境不同;二是人为干扰程度以及种间对生存资源的竞争压力差异明显。
在群落内部,不同植物居群为争夺有限的生存资源而相互竞争,导致个体死亡率明显上升。在鹊山省级自然保护区,当胡桃楸的幼苗或幼树生长发育形成中龄树后,群落的结构、郁闭度和盖度发生变化,受光照和营养条件以及伴生树种的影响,Ⅲ级至Ⅴ级胡桃楸个体死亡率呈现上升趋势。总体上看,研究区域内胡桃楸个体期望寿命(ex)波动较小,其中,低径级的ex值相对较高,高径级的ex值相对较低。这是由于低径级的胡桃楸个体生存空间比较充裕,且对养分和水分的需求量较小,对资源和生存空间的竞争不强;而高径级的胡桃楸个体从环境中获得的生存资源均不能满足其生长需求,种间和种内竞争较强,导致自疏作用和他疏作用增强,因此,高径级的胡桃楸个体期望寿命下降。
王东娜[4]25-27的研究结果表明:以ISSR分子标记为基础,分布于东北地区的6个胡桃楸居群的遗传多样性均较高,且不同居群的亲缘关系与其地理距离基本相一致。本研究中,通过SRAP分子标记分析,显示鹊山省级自然保护区的胡桃楸居群内存在丰富的遗传变异,且居群内各样本间的遗传关系与其地理分布有一定的关联。考虑到生态环境的同质性,研究区域内胡桃楸个体间叶片和果形等表型性状的差异可能与其遗传特性相关。
虽然鹊山省级自然保护区的胡桃楸居群属于增长型,但该居群易受人为干扰、生境条件、繁殖特性以及种内竞争等因子的影响,其中,人为干扰是关键的致危因子。由于该保护区内胡桃楸居群的遗传多样性较高,因而,应制定相应的保护策略以维持现有的遗传多样性水平。根据居群的生境特点,建议采用人工扰动的方式使种子能落入适宜的土壤环境中萌发形成幼苗,维持该居群的天然更新。另外,建议强化退耕还林、幼林抚育等措施,提高幼林成活率,扩大成林覆盖面积,从而保持群落结构稳定。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
植物研究(2022年5期)2022-10-12
河北科技师范学院学报(2022年2期)2022-08-26
江西农业学报(2022年5期)2022-08-10
林业调查规划(2022年3期)2022-06-13
浙江中医药大学学报(2021年6期)2021-07-12
园艺与种苗(2021年3期)2021-05-26
种子(2019年6期)2019-07-13
电影(2018年11期)2018-11-24
美食(2017年5期)2017-05-11
