
冷冻温度对非发酵面团蛋白质结构及面团特性的影响
2020-06-11 13:52:24杨静洁张影全郭波莉顾熟琴
中国粮油学报 2020年5期
杨静洁 张 波 张影全 郭波莉 顾熟琴
(北京农学院食品科学与工程学院1,北京 102206) (中国农业科学院农产品加工研究所;农业农村部农产品加工综合性重点实验室2,北京 100193)
顾熟琴,女,副教授,1962年出生,农产品加工及安全控制
冷冻非发酵面团一般用于速冻汤包、蒸饺、烧麦等中国传统主食的生产[1],由于其方便、快捷的特点,已经逐渐成为中国传统面制品实现主食工业化生产的主要产品形式,受到越来越多消费者的青睐。
冷冻过程中面团品质的劣变降低了产品食用品质,缩短了产品的货架期[2]。目前,很多学者研究了冷冻对面团加工品质及产品食用品质的影响[3, 4],证明冷冻面团品质劣变与面团中蛋白质品质劣变具有很大的关系[5]。冷冻温度决定冷冻速率,影响冷冻面团质量。Yi等[6]发现适当的冷冻温度不仅可以对面包面团的弹性、持气性具有积极作用,并且可以降低面包外壳硬度、维持面包体积。Akbraian等[7]研究冷冻甜面团发现,冷冻温度越高,面包内聚性、弹性下降越明显;Olivera[8]等研究冷冻意大利面发现,冷冻温度越低,产品的质构、感官特性与新鲜意大利面条越接近;黄忠明等[9]研究非发酵面团发现,降低冷冻温度,可以降低面团失水率,提高面团拉伸强度;李杰平等[10]发现随着冷冻温度降低,汤包皮面团的弹性模量、黏性模量上升,硬度下降,熟汤包皮的质构特性得到改善。尽管有众多学者研究了冷冻温度对面团品质及其加工制品品质的影响,但是对于不同冷冻温度下,非发酵面团中蛋白质结构变化的系统性研究还比较少,其与面团质构特性之间的关系尚不明确。
本研究以不同的冷冻温度冻结面团,分析面团中蛋白质组分、谷蛋白大聚体(GMP)含量、游离巯基含量、二级结构,以及面团质构特性变化,探讨蛋白质结构与面团质构特性的关系,为传统中式非发酵冷冻面团产品的冷冻工艺优化提供借鉴。
1 材料与方法
1.1 材料与试剂
小偃22号小麦品种:清理样品后,根据籽粒硬度(硬质小麦)与含水量(11.3%)确定润麦加水量。润麦24 h,采用MLU 202实验磨制粉,出粉率约为70%。
实验用小麦粉质量性状见表1。

表1 实验用小麦粉质量性状
1.2 实验方法
1.2.1 面团制作
为了保证面团搅拌程度的一致性,本实验采用827504型粉质仪和面。按照14%含水率校正后的小麦粉质量称取样品,按照粉质仪吸水率加入蒸馏水,当面团稠度达到500 BU时取出面团;将面团分割成60 g的小面团,利用860033/002型拉伸仪的搓圆设备整形后,用保鲜膜包裹,置于-20、-30、-40 ℃超低温冰箱中进行冷冻。当面团中心温度降至-18 ℃时,将面团转移至-18 ℃冰箱中冻藏24h后,取出待用。取搅拌稠度达到500 BU且不进行冷冻处理的面团为对照组,将面团分割、揉圆;一部分用液氮进行快速冷冻后,再冻干,用于蛋白质结构分析;另一部分直接用于质构分析。
1.2.2 冷冻曲线的绘制
将温度计置于面团中心,每隔30 min记录一次温度,面团在三种冷冻温度条件下冷冻至中心温度达到-18 ℃过程中记录的温度变化曲线,即为冷冻曲线。
1.2.3 样品处理方法
冷冻面团于25 ℃、相对湿度85%的恒温恒湿箱中解冻,至面团中心温度达25 ℃后,部分样品用于质构实验。另取部分解冻面团样品于液氮中快速冷冻后,用ALPHA 1-2LD PLUS 真空冷冻干燥机进行真空冷冻干燥72 h,绝对真空度为3.7×10-5MPa,冷冻阱温度-50 ℃。对冻干样品采用MM400研磨仪粉碎,过60目筛,样品粉末于4 ℃冷藏,用于蛋白质结构分析。
1.2.4 游离巯基(-SH)含量测定
参照Anderson等[11]的方法进行测定。
1.2.5 谷蛋白大聚体(GMP)含量测定
取250 mg冻干粉加入23 mL质量分数1.5%的SDS-0.05 mol/L磷酸盐缓冲液(pH=6.9),漩涡振荡至完全分散无明显结块,在30 ℃下以1 000 r/min振荡5 min后,25 ℃, 11 600 g离心30 min,弃上清液,采用杜马斯燃烧定氮仪测定沉淀中蛋白质含量,即为GMP近似含量。每个样品做3次重复,取平均值。
1.2.6 蛋白质组分分析
利用体积排阻高效液相色谱(SE-HPLC)法进行分析。
蛋白质提取:参照Rakszegi等[12]、Wang等[13]和刘锐等[14]的方法,略修改。提取分为两步:1)将70 mg冻干样品加入7 mL质量分数为0.5%~0.1 mol/L的SDS磷酸钠缓冲液(pH=6.9),漩涡振荡混匀后,用ComfortTM恒温混匀仪在30 ℃以750 r/min恒温振荡混匀30 min,室温下13 000 g离心20 min。取上清液,这部分为可溶性蛋白质。2)向沉淀中再加入7 mL提取液,将沉淀物捣碎后恒温振荡10 min,混匀后,使用Y mnl-150Y超声波细胞破碎仪进行超声粉碎(超声波细胞破碎仪每隔2 s工作2 s,总时长1 min);随后以13 000 g离心15 min;取上清液,这部分为不溶性蛋白质。所有提取液在上样前均过0.45 μm滤膜,每份测定2次。
色谱系统:Agilent 1200高效液相色谱仪。
色谱条件:色谱柱为TOSOH TSK-gel G4000 SWxl;流动相为0.1% SDS-0.02 mol/L 磷酸盐缓冲液(pH=6.9),流速 0.7 mL/min,进样量 20 μL。采用1200二级管阵列检测器,检测波长210 nm。蛋白质标准品:甲状腺球蛋白(670 000)、γ-球蛋白(150 000)、清蛋白(44 300)、核糖核酸酶 A(13 700)。
参照Wang等[13]和刘锐等[14]方法,将冷冻面团样品中可溶性蛋白、不可溶性蛋白的排阻色谱分为6个峰,然后进行蛋白组分计算与分析。
1.2.7 蛋白质二级结构分析
图谱扫描:称取冻干样品2 mg,加入0.2 g干燥过的优级纯溴化钾作为稀释剂,与样品一起在玛瑙研钵中充分研磨后压片,10 kg压力保持30 s,然后将制得的均匀透明薄片放入Tensor 27 傅里叶变换红外光谱仪(FTIR)中测定。扫描范围400~4 000 cm-1,分辨率4 cm-1,信号扫描累加64次。每种处理的样品重复测定3次。
图谱处理:采用仪器自带POUS 8.1和Peak Fit 4.12 对图谱进行分析处理。首先对所得原始吸收图谱进行背景扣除水分、CO2气氛补偿,然后对位于酰胺I带(1 600~1 700 cm-1)的吸收峰进行基线校正、傅里叶自退去卷积、二阶求导分析,使得叠加的各子峰得到分辨。根据二级结构对应峰面积占总峰面积的比例定量计算出二级结构含量。参照宋国胜等[15]方法,确定各个波长区间所对应的二级结构归属:β-折叠,1 615~1 637 cm-1、1 682~ 1 700 cm-1;无规则卷曲,1 637~1 645 cm-1;α-螺旋,1 646~1 664 cm-1;β-转角,1 664~1 681 cm-1。
1.2.8 面团质构特性的测定
将解冻后面团用TA-XT2i型质构仪测定面团的质构特性。具体步骤为:将解冻后面团制成直径约6 cm球体放在载物台上,调整质构仪为TPA模式;实验采用探头:P/50铝制圆柱形探头;测定参数设置为:测前速度3 mm/s,测试速度1 mm/s,测后速度3 mm/s,压缩比例70%,2次压缩时间间隔10 s,触发力10 g。每个样品做6次重复,取平均值。本实验中选择硬度、弹性、黏附性、内聚性、回复性作为质构评价指标。
1.3 统计分析
采用Excel 2010、Origin8.0进行数据整理与图表绘制,所有数据采用平均值±标准差表示。采用SPSS 19.0中One-way ANOVA程序进行方差分析,采用Duncan法进行多重比较。
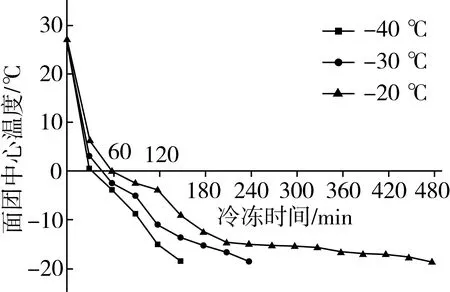
图1 不同温度条件下的面团冻结曲线
2 结果与分析
2.1 不同温度条件下面团冻结曲线
面团经-40、-30、20 ℃冷冻至中心温度为-18 ℃分别需要150、240、480 min(图1),对应的冷冻速率分别为18.2、11.4、5.7 ℃/h。因此,冷冻温度越低,冷冻速率就越高。从图3看出,面团的冻结经过为三个阶段,即表面降温(预冷)阶段、最大冰晶生成(相变)阶段和持续降温阶段[16]。不同冷冻温度下面团冻结曲线在第一阶段基本接近,没有较大差别;但在第二阶段,面团在-40、-30、-20 ℃下冷冻,通过最大冰晶生成带用的时间分别为30、40、60 min。通过最大冰结晶带的时间决定了面团中形成冰晶的数量、尺寸大小及分布[9]。冷冻速率快,通过最大冰结晶带用时短,晶核生长的时间越短,就会形成体积较小的冰晶,对面团内部结构的破坏程度越小;反之,则会形成体积较大的冰晶。
2.2 不同温度条件下面团蛋白质结构差异
2.2.1 游离巯基(-SH)含量
由图2可知,冷冻对面团中游离巯基含量具有显著影响。与对照组相比,经-40、-30、-20 ℃冷冻的面团中游离巯基含量显著上升了0.22、0.49、1.63 μmol/g。在-20 ℃下冷冻的面团中游离巯基含量显著高于-30 ℃和-40 ℃冷冻的面团;而在-30 ℃与-40 ℃下冷冻的面团之间游离巯基含量无显著差异。
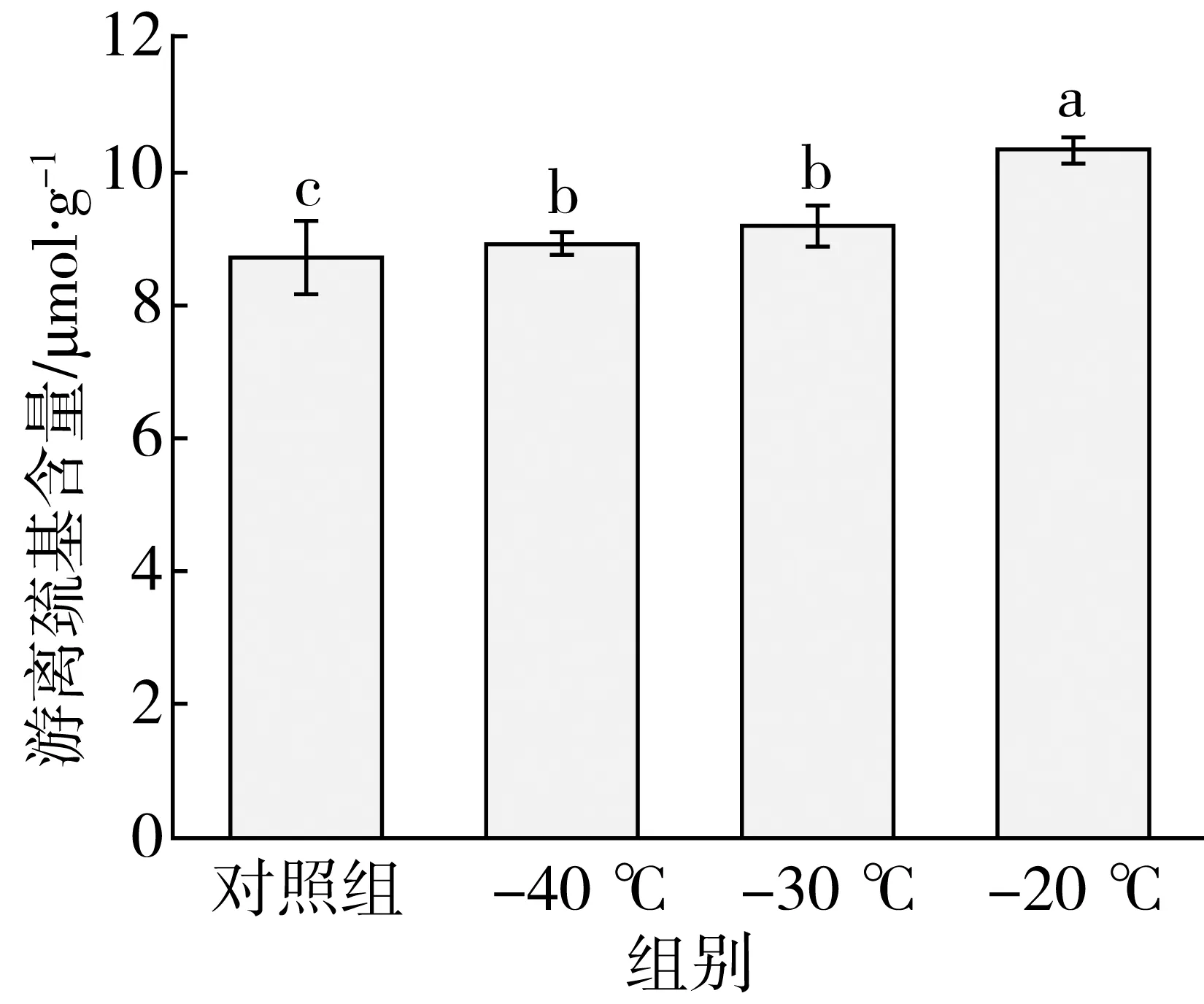
注:不同小写字母表示P<0.05水平上差异性显著,下同。图2 冷冻温度对面团中游离巯基含量的影响
2.2.2 蛋白质二级结构
由表2可知,对照组面团中蛋白质的二级结构包括α螺旋、β-折叠、β-转角、无规则卷曲四种结构,与Zhang[1]、Meziani[17]的结果基本一致。冷冻对面团中蛋白二级结构具有显著影响。与对照面团相比,所有冷冻组面团中α-螺旋、β-转角含量均有不同程度的下降,而β-折叠、无规则卷曲结构含量上升。但对照组面团中α-螺旋、β折叠结构含量与-30 ℃、-40 ℃冷冻的面团基本无显著性差异,与-20 ℃冷冻面团差异性显著。在-20 ℃冷冻的面团中α-螺旋、β-折叠、无规则卷曲结构与-30 ℃、-40 ℃冷冻的面团差异性显著,而-30 ℃与-40 ℃冷冻的面团在这三种二级结构上无显著性差异;三种冷冻温度下冷冻的面团中β-转角结构无显著性差异。

表2 冷冻温度对面团中蛋白二级结构相对含量的影响
注:同一列不同小写字母表示P<0.05水平上差异性显著,下同。
2.2.3 谷蛋白大聚体(GMP)含量
冷冻温度对面团中谷蛋白大聚体含量具有显著影响。与对照组面团相比,冷冻后面团中的GMP含量显著下降,-40、-30、20 ℃冷冻的面团中GMP含量分别下降了0.19、0.43、0.52%(图3)。在-40 ℃冷冻的面团中GMP含量显著高于与-20、-30 ℃冷冻的面团,而-20 ℃冷冻与-30 ℃冷冻的面团中GMP含量无显著性差异。Graveland等[18]指出谷蛋白大聚体主要是通过二硫键交联而成,不同冷冻温度下GMP含量的变化与图2游离巯基含量变化相吻合,说明冷冻过程会使GMP以二硫键断裂的方式发生解聚,并且冷冻温度越高,冷冻速率越慢,GMP解聚程度越大。
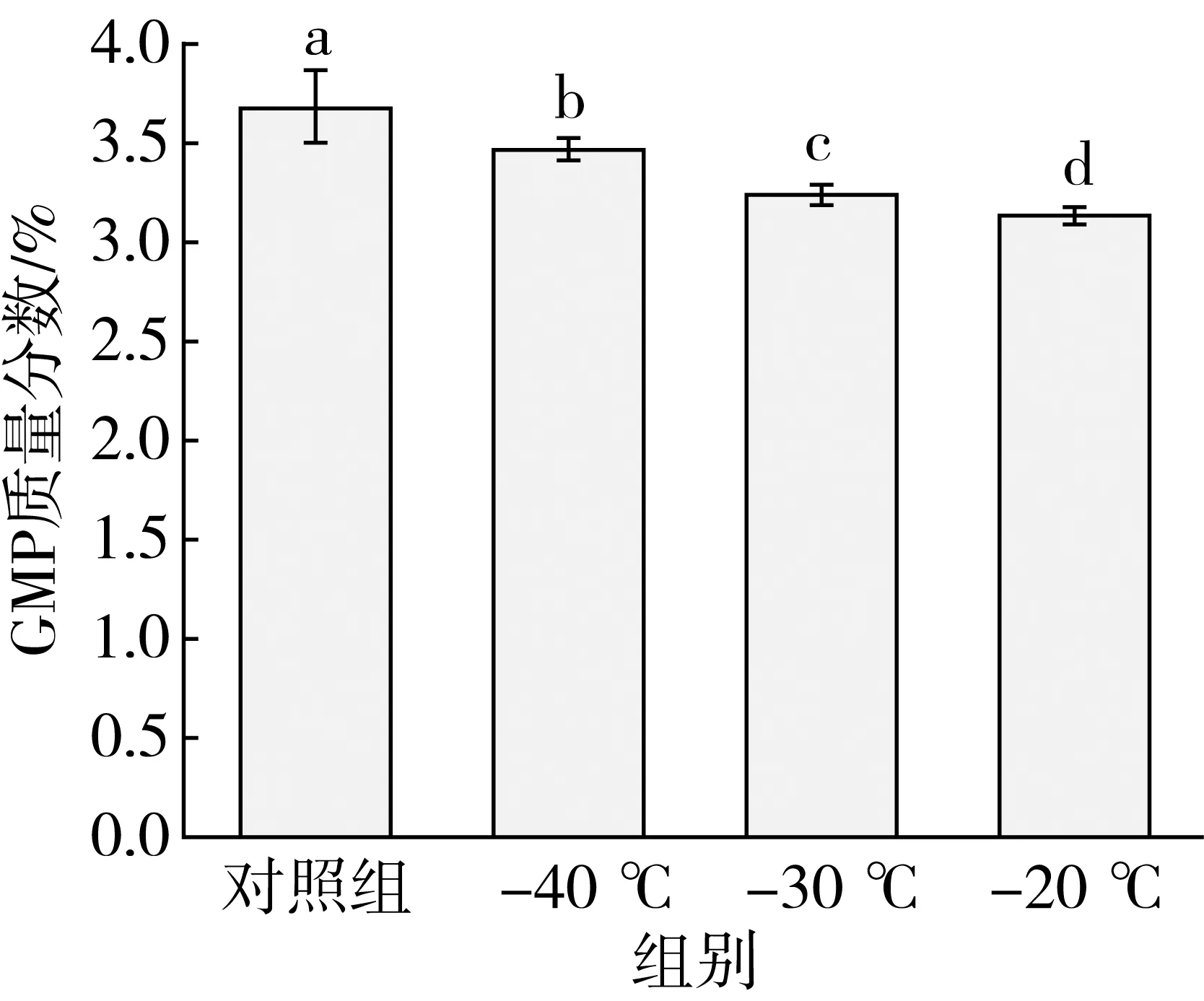
图3 冷冻温度对面团GMP含量影响
2.2.4 蛋白质组分
与对照组面团相比,面团冷冻后蛋白质组分发生显著变化,总体表现为LPP显著降低,LMP和SMP显著升高(表3)。这说明冷冻处理后的面团所含聚合体蛋白比例减少,单体蛋白比例增加,即蛋白质聚合程度降低。不同冷冻温度对面团组分变化影响不同。在-40 ℃下冷冻的面团中LPP、LMP、SMP含量与-20 ℃、-30 ℃冷冻的面团差异性均显著;而-30 ℃冷冻的面团与-20 ℃冷冻的面团仅在LMP含量上差异性显著,在LPP、SMP含量上无显著性差异。

表3 冷冻温度对面团蛋白组分的影响
2.3 冷冻温度对面团质构特性影响
冷冻对面团质构具有显著性影响。冷冻后面团的硬度、黏附性和回复性呈上升趋势,弹性和内聚性呈降低趋势。对照组面团与-40 ℃冷冻的面团之间质构指标基本无显著差异,与-20 ℃、-30 ℃冷冻的面团质构指标间差异性显著(表4)。
对于硬度、黏附性,-20 ℃冷冻与-40 ℃冷冻的面团之间差异性显著;对于弹性,-30 ℃冷冻的面团与-40 ℃冷冻的面团之间差异性显著,但-20 ℃冷冻面团与-30 ℃冷冻的面团之间差异性不显著;对于内聚性,三种冷冻温度下冷冻的面团间无显著差异;对于回复性,-20 ℃冷冻面团与-30、-40 ℃冷冻面团之间差异性显著,而-30 ℃冷冻的面团与-40 ℃冷冻的面团间无显著性差异。-20 ℃冷冻对面团质构特性影响最为显著。
2.4 面团蛋白结构与其质构特性的相关关系
相关分析结果表明,谷蛋白大聚体含量与面团硬度呈显著负相关,而与弹性、内聚性呈显著正相关;大分子聚合体蛋白与面团内聚性呈显著正相关,与面团回复性呈显著负相关;大分子单体蛋白与面团内聚性呈显著负相关、与面团回复性呈显著正相关;小分子单体蛋白与面团回复性呈显著负相关;β-转角与内聚性呈极显著相关;无规则卷曲与内聚性呈显著负相关,与回复性呈显著正相关。蛋白组分与面团质构特性之间相关性更显著(表5)。

表5 蛋白结构指标与面团TPA参数间的相关性分析
注:*代表P<0.05水平上差异性显著;**代表P<0.01水平上差异性显著。

表4 不同冷冻温度冷冻的面团解冻后TPA指标值
3 讨论
3.1 冷冻温度对蛋白质结构的影响
冷冻后面团中游离巯基含量升高,谷蛋白大聚体含量降低,聚合体蛋白发生解聚,蛋白二级结构发生改变。这是因为冷冻过程中,冰晶可能对蛋白质分子间的二硫键造成破坏[4],分子结构变得松散,蛋白质结构发生改变,导致聚合体蛋白发生解聚;Ribotta等[19]和Zhao等[20]等认为冷冻过程中GMP的解聚可能是因为冰晶的形成以及水分的迁移对二硫键、疏水键、离子键造成破坏,Bhatnagar等[21]认为除了冰晶的影响,冷冻带来的冷冻浓缩及低温效应也可能是导致GMP解聚的原因。冷冻后,对蛋白分子结构起到骨架作用的α-螺旋结构中的氢键等非共价键受到冰晶的机械破坏,蛋白质的亲水和疏水残基暴露在外界环境中,导致蛋白分子间、分子内出现新的交联现象,从而改变了蛋白质的二级结构[22]。
由面团冻结曲线可知,冷冻温度决定了冷冻速率,冷冻温度越低,冷冻速率越高。本研究中,随着冷冻温度的降低,面团中蛋白质解聚程度及二级结构变化程度降低,这是因为冷冻温度低时,冷冻速率较高,面团中的水分子没有足够的时间转移就降温至冻结点,因而会在面筋网络结构附近形成体积较小的冰晶体,其对面团内部面筋结构及大分子物质之间的非共价键破坏程度比较小;相反,冷冻温度高时,面团冷冻速率较慢,由于降至冻结点温度以下需要的时间很长,冰晶形成的速度小于水分子的扩散渗透速度,水分子可以进行不定向运动,形成较大的冰晶体,大的冰晶体对面团网络结构的机械损伤较大,更容易造成面筋纤维状结构变细或者断裂,加剧大分子蛋白的解聚以及蛋白分子构象的改变[9, 10]。因此,较高的冷冻速率能够抑制面团中大冰晶的形成,进而减少冰晶对维持蛋白骨架α-螺旋结构中氢键的破坏,对维持蛋白结构稳定具有积极作用。
李学红等[23]将麦谷蛋白、麦醇溶蛋白在-40 ℃冷冻并冻藏至28 d时,β-折叠结构含量均降低、α-螺旋结构含量均上升,并且麦谷蛋白中二者的变化幅度显著高于麦醇溶蛋白;王沛[5]将纯品面筋蛋白、谷蛋白、醇溶蛋白在-35 ℃冷冻后于-18 ℃进行冻藏,冻藏至30 d时三者α-螺旋结构分别下降了10.47%、13.97%、25.83%,反向平行β-折叠结构分别增加了3.24%、7.75%、12.11%。本研究中面团中蛋白二级结构变化程度低于前人研究结果,一方面可能是因为冻藏时间选择的不同,随着冻藏时间延长,冰晶的生长及重结晶对蛋白中非共价键的破坏加剧,蛋白二级结构发生更为剧烈的改变;另一方面可能是因为原材料的不同,面团是一个复杂的混合体系,其内部各种物质之间可能形成了更多的非共价键,从而使蛋白二级结构更加稳定,这还需要进一步的实验进行验证。本实验中,冷冻对面团中蛋白的四种二级结构均有显著性影响;Meziani等[17]研究甜面团则发现,冷冻速率仅对面团中α-螺旋、β-折叠两种结构具有显著影响。这可能是因为盐、糖、黄油等物质的添加对面团内部非共价键的形成具有积极促进作用,从而降低了冷冻对蛋白二级构象的消极影响。
3.2 冷冻温度对面团质构特性的影响
与对照组面团相比,冷冻导致非发酵面团硬度上升,弹性、内聚性下降。由本研究中蛋白质结构变化结果可知,冷冻会引起二硫键、氢键以及离子键发生变化,这导致面团内部聚合物交联情况发生变化,淀粉与蛋白质之间的作用力也发生改变[24],进而改变面团内部结构,最终导致面团质构发生变化。降低冷冻温度,提高冷冻速率可以降低非发酵面团质构变化程度,这与黄忠民等[9]、李杰平等[10]研究结果一致;Alain Le-Bail等[25]、Kondakci等[26]研究发现,过高或者过低的冷冻速率都会降低发酵面团质构特性,而适宜的冷冻速率可以减少冰晶对面团内部结构的破坏并且有利于酵母活性的提高。因此,冷冻速率对不同种类的冷冻面团影响不同。
较低的冷冻温度对维持非发酵面团中蛋白质结构特性具有积极作用,从而可以降低非发酵面团品质劣变程度;但过低的冷冻温度会影响发酵面团中酵母的活性从而降低发酵面团的品质。冷冻面团中其他组分的添加会导致面团加工特性发生变化,不同冷冻面团加工终产品对冷冻工艺的要求不同,因此今后还需要针对不同冷冻制品的面团进行研究,重点研究面团品质劣变程度与最终产品食用品质变化程度之间的关系。
4 结论
冷冻后非发酵面团中蛋白质结构及面团质构特性发生显著变化。随着冷冻温度的降低,冷冻速率的上升,面团中聚合体蛋白比例上升,单体蛋白含量下降,GMP含量增加,游离巯基含量降低,蛋白聚合程度增加;α-螺旋含量增加,β-折叠含量降低,无规则卷曲含量降低,β-转角无显著性变化。
随着冷冻温度的降低,面团的硬度、回复性、黏附性呈降低趋势,内聚性、弹性呈上升趋势;蛋白质结构与面团质构之间具有显著相关性。
降低冷冻温度,可以提高面团的冷冻速率,进而降低冷冻对非发酵面团中蛋白质结构及面团品质的破坏程度。
猜你喜欢
现代商业银行·财富生活(2023年12期)2024-01-20 00:02:24
食品安全导刊(2021年21期)2021-08-30 08:22:00
科普童话·学霸日记(2021年12期)2021-05-30 10:48:04
学生导报·东方少年(2019年11期)2019-06-11 11:03:24
现代营销(创富信息版)(2018年6期)2018-09-05 03:14:32
食品研究与开发(2016年20期)2016-11-05 02:19:01
琴童(2016年5期)2016-05-14 15:43:58
财经问题研究(2015年4期)2015-05-04 20:20:27
食品工业科技(2014年11期)2014-03-11 18:16:08
食品工业科技(2014年7期)2014-03-11 18:14:58
