
自噬在绿脓杆菌感染时对中性粒细胞粘附的作用
2020-06-10 12:05袁喆晨汪林芳苏芬芬凌晓晶陈时豪曹佳泽朴正浩
杭州师范大学学报(自然科学版) 2020年3期
袁喆晨,汪林芳,苏芬芬,凌晓晶,陈时豪,曹佳泽,方 倩,朴正浩
(杭州师范大学医学院,浙江 杭州 311121)
0 引言
绿脓杆菌(Pseudomonasaeruginosa)是一种革兰氏阴性杆菌,广泛存在于自然界,是引起肺炎的主要病原菌[1].因绿脓杆菌具有对多种抗菌素的耐药性,绿脓杆菌性肺炎治疗较困难,病死率较高[2-3].先天免疫系统是控制绿脓杆菌感染的主要防御机制,但是机体不能及时清除病原菌是导致宿主预后不良的重要因素[3-4].
中性粒细胞是先天免疫系统中抵抗细菌感染的主要防御细胞,在趋化因子的诱导下经旋转、粘附、爬行和转移等一系列级联反应过程向炎症部位募集,对侵入的病原体发挥吞噬杀伤和清除作用[5-6].所以,有效地促进中性粒细胞的募集将有利于及时清除病原菌.绿脓杆菌的感染会引起中性粒细胞借助β2整合素(integrin)募集到炎症部位[4,7].整合素是细胞的跨膜分子,主要由β2和α亚基组成,其中LFA1(αLβ2;CD11a/CD18)和Mac1(αMβ2;CD11b/CD18)为中性粒细胞的主要分子[5-6,8].中性粒细胞的Mac1通过β2整合素与内皮细胞的表面分子相互作用,完成爬行和粘附的过程[5,9].
自噬(autophagy)是对胞质蛋白和细胞器进行降解的一种过程,对稳定细胞内环境具有十分重要的作用[10-11].在肿瘤细胞中自噬能够激活整合素,增加细胞与细胞外基质的粘附能力,并促进细胞的转移[12-13].在中性粒细胞中,自噬可以促进细胞脱颗粒[14]、杀菌和形成细胞外诱捕网(neutrophil extracellular trap, NET)[15],还可以促进中性粒细胞的分化及中性粒细胞成熟标志CD11b/CD18的表达[16].但自噬是否能调节中性粒细胞的募集还不清楚.
本研究通过分离小鼠骨髓中性粒细胞进行了体外的粘附实验,以了解自噬在绿脓杆菌感染时对中性粒细胞粘附的作用,进而为探索自噬在中性粒细胞募集中的作用提供理论依据.
1 材料与方法
1.1 试剂
3-甲基腺苷(3-methyladenine,3-MA)(HY-19312)购于MedChem Express;胎牛血清(11012-8611)购于四季青;红细胞裂解液(C3702)购于Beyotime Biotechnology;PecrollTM(10245267)购于Bioleaf Biotech;4%多聚甲醛固定液(E672002)购于BBI Life Sciences;BSA封闭液(C500036-0250)和30%的聚丙烯乙酰胺(B546018-0500)购于Sangon Biotech;RPMI1640(MA0215)和BCA蛋白浓度测定试剂盒(MA0082)购于Meilun Biotech;β-actin(#3700)和LC3A/B(#4108)抗体购于Cell Signaling Technology;CD11b-PE(12-0112)和CD18-FITC(11-0181)流式细胞仪荧光抗体购于eBioscience;ImmobilonTMWestern Chemiluminescent HRP Substrate(WBKLS0100)购于MILLIPORE.
1.2 小鼠骨髓中性粒细胞的提取
提取C57BL/6野生型小鼠大腿骨,用注射器抽取培养液反复灌洗骨髓,离心后收集骨髓细胞.用红细胞裂解液裂解其中的红细胞,用1×PBS终止反应,1 500 r/min离心5 min收集细胞.细胞用0.5 mL 1×PBS 混悬,小心置于已制备好的Percoll细胞分离液中,1 000 r/min的转速4 ℃离心30 min,并去除减速自然停止.离心结束后可见分层,取第三层细胞,用无菌的1×PBS洗涤两次,通过FACS检测中性粒细胞的百分比(CD11b/Gr1).
Percoll密度梯度分离液的制备:取100%的Percoll(按9∶1的比例混合Percoll原液和10×PBS),用1×PBS稀释成80%、65%、50%的3种Percoll液体,分别以体积4、3、3 mL依次缓慢加入一支15 mL离心管中,制作浓度梯度.
1.3 免疫印迹实验
按常规方法收集各组处理后的细胞,每组细胞加100 uL RIPA细胞裂解液(加磷酸酶抑制剂和1 mmol/L PMSF),在冰上裂解30 min后,收集裂解液在12 000 r/min条件下离心30 min,收集上清的蛋白质,用BCA蛋白浓度测定试剂盒测定蛋白浓度.取等量的各组蛋白质,加1×的上样缓冲液,于100 ℃条件下煮蛋白5 min,获得最后的蛋白样品.在12%的SDS-PAGE胶中每孔加入40 μg蛋白样品,利用伯乐电泳系统进行电泳,设置电压为100 V,电泳90 min.电泳完成后,在100 V的电压条件下转膜60 min.膜用BSA封闭液封闭60 min,然后4 ℃孵育一抗过夜.次日,用1×TBST在摇床上洗膜(每次5 min,共3次).在室温孵育二抗2 h后,用1×TBST缓冲液洗膜,最后用辣根过氧化物酶底物反应后,在暗室显影和定影.
1.4 细胞粘附实验
提取的骨髓中性粒细胞接种在涂有纤维粘连蛋白的培养板中,用绿脓杆菌(MOI 25)感染不同时间后,去除上清液中的未贴壁细胞,并用1×PBS洗一次培养板.每孔加入适量的1×PBS刮贴壁的细胞,收集细胞并计数为粘附细胞的数量.
1.5 流式细胞术
提取骨髓中性粒细胞,用绿脓杆菌(MOI 25)感染不同时间后收集全部细胞(悬浮和贴壁的细胞),用1×PBS清洗细胞.加入CD11b-PE和CD18-TIFC荧光抗体2 μL(抗体稀释100倍)4 ℃避光孵育细胞15 min.再用1×PBS清洗一次细胞后,加入200 μL 4%多聚甲醛固定液固定细胞,用流式细胞仪观察中性粒细胞整合素CD11b和CD18的荧光强度.
2 结果
2.1 绿脓杆菌感染引起骨髓中性粒细胞的粘附,增强整合素CD11b/CD18的表达

提取小鼠骨髓中性粒细胞,分成4组,每组细胞数量为5.1×105个.接种在涂有纤维粘连蛋白的培养板上,再用绿脓杆菌PA14菌株(MOI 25)感染细胞10、30、60 min后,去除上清液中未贴壁的细胞,清洗一次培养板后刮取贴壁的细胞并计数.0组为没有感染的实验组,每组3个复孔,实验重复3次.
图1 绿脓杆菌感染引起骨髓中性粒细胞的粘附
Fig.1Pseudomonasaeruginosapromotes the adhesion of neutrophils
中性粒细胞是先天免疫系统中抵抗细菌感染的主要防御细胞,募集到感染部位发挥抗感染的作用,而粘附是中性粒细胞募集级联反应过程中的一个必经阶段[5-6].在绿脓杆菌感染时中性粒细胞依赖β2整合素募集到感染部位[4].本课题组分离了小鼠骨髓中性粒细胞,感染绿脓杆菌进行研究时发现,中性粒细胞的粘附细胞数量随着感染时间的延长而增多,在30 min时粘附的细胞数量达到顶峰(图1).整合素是中性粒细胞发挥功能所需的跨膜蛋白[5-6],绿脓杆菌感染后中性粒细胞表面整合素CD11b/CD18的表达也增加(图2).由此确定,绿脓杆菌感染引起骨髓中性粒细胞的粘附和整合素CD11b/CD18的表达增强.
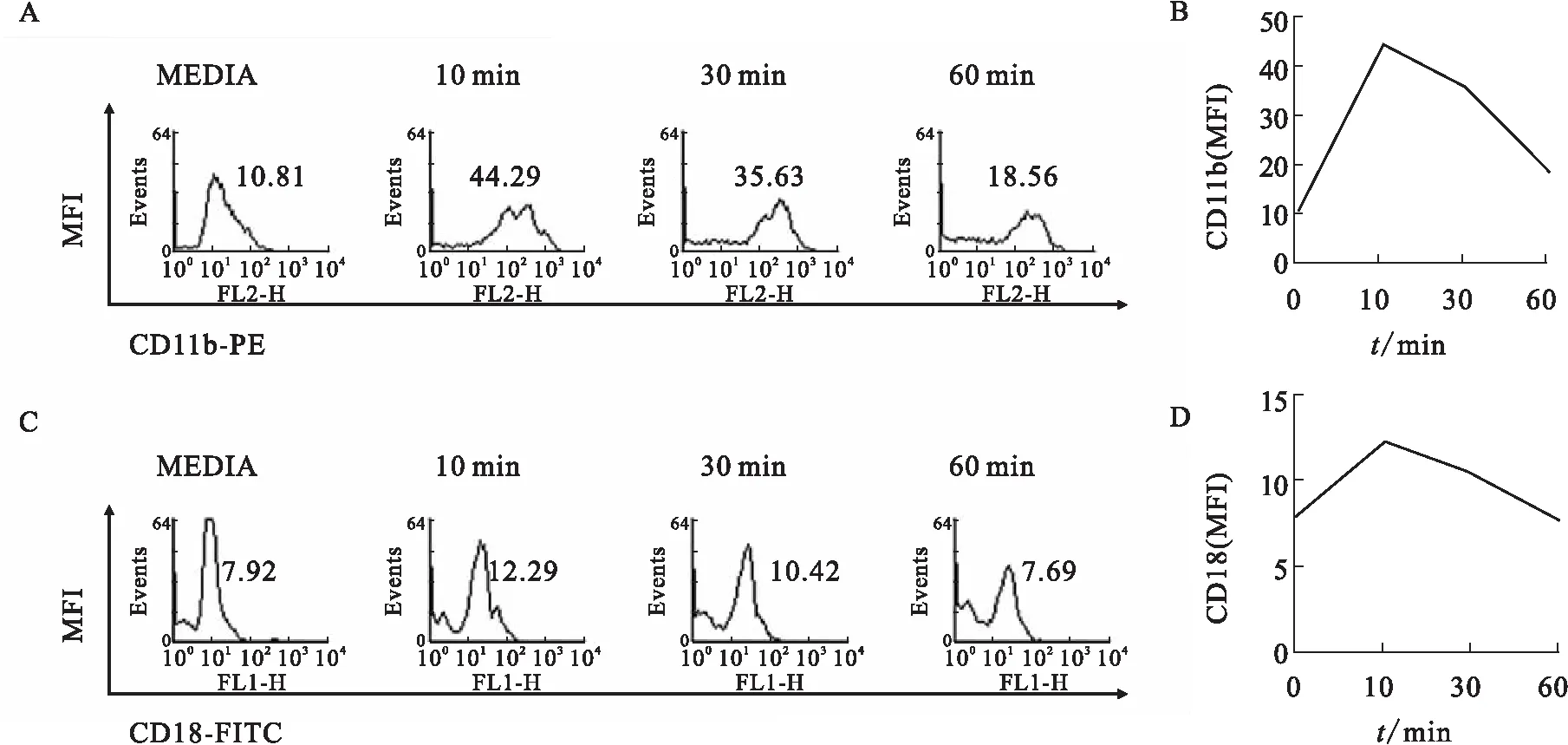
A、B:提取小鼠骨髓中性粒细胞,平均分成4组,每组细胞数量为5.5×105个.用绿脓杆菌(MOI 25)感染10、30、60 min,收集细胞后用流式细胞仪检测CD11b的荧光强度,并制作折线图分析趋势.实验重复3次.
C、D:提取小鼠骨髓中性粒细胞,平均分成4组,每组细胞数量为5.5×105个.用绿脓杆菌(MOI 25)感染10、30、60 min,收集细胞后用流式细胞仪检测CD18的荧光强度,并制作折线图分析趋势.实验重复3次.
图2 绿脓杆菌感染增强整合素CD11b/CD18的表达
Fig.2Pseudomonasaeruginosaenhances the expression of integrin
2.2 绿脓杆菌感染引起骨髓中性粒细胞自噬的发生
自噬调节中性粒细胞的功能,参与细菌感染的炎症反应[4,17].在肿瘤细胞中自噬能够激活整合素,增加细胞粘附并促进细胞的转移[12-13].为了研究中性粒细胞的粘附机制,本研究通过免疫印迹实验观察了自噬标记性分子LC3-I/II的表达,发现随着绿脓杆菌感染时间的增加,LC3-II蛋白的表达增加(图3A).3-MA是自噬的抑制剂[18],用3-MA预先处理骨髓中性粒细胞后感染绿脓杆菌发现,LC3-II的表达明显被抑制(图3B).可见,绿脓杆菌感染可引起中性粒细胞的自噬.

A:提取野生型小鼠的骨髓中性粒细胞,平均分成4组,每孔细胞数量为2.6×106个.用绿脓杆菌(MOI 25)感染10、30、60、120 min后获取蛋白,每孔蛋白量为40 μg左右.进行免疫印迹实验,观察LC3-I/II和Actin.
B:从小鼠的骨髓中分离骨髓中性粒细胞,每孔细胞数量为2.1×106个,3-MA抑制剂组预先处理1 h(浓度分别为10、50、100、200 μmol/l).清洗细胞后用绿脓杆菌(MOI 25)感染细胞60 min.获取蛋白,每孔蛋白量为40 μg左右,并进行免疫印迹实验观察LC3-I/II的表达.
图3 绿脓杆菌感染激活骨髓中性粒细胞中自噬的发生
Fig.3Pseudomonasaeruginosainduces autophagy in neutrophils from bone marrow
2.3 自噬调节绿脓杆菌感染时骨髓中性粒细胞的粘附和整合素的表达

提取小鼠的骨髓中性粒细胞,每孔细胞数量为2.9×106个.预先用3-MA(200 μmol/L)或DMSO处理后清洗一次细胞,接种在涂有纤维粘连蛋白的培养板上,再用绿脓杆菌(MOI 25)感染不同时间,观察粘附细胞的数量.0组为没有感染的组.
图4 自噬调节绿脓杆菌感染时骨髓中性粒细胞的粘附
Fig.4 Autophagy regulates the adhesion of neutrophils duringPseudomonasaeruginosainfection
为了确定自噬对中性粒细胞粘附的作用,用3-MA抑制剂预先处理骨髓中性粒细胞,结果发现自噬的抑制剂显著地抑制中性粒细胞的粘附(图4).CD11b是中性粒细胞执行功能的重要分子,在缺乏自噬的细胞中的表达显著地降低[16].本研究发现自噬的抑制剂3-MA抑制整合素CD11b的表达(图5A、B),但是CD18的表达几乎没有被抑制(图5C、D).因此,可以确定自噬在骨髓中性粒细胞的粘附和整合素的表达中起重要作用.
3 讨论
绿脓杆菌广泛分布于自然界,常在医院内引起感染,而且具有对多种药物的耐药性,所以治疗困难[1,19].绿脓杆菌感染引起中性粒细胞的募集[7],从骨髓经旋转、粘附、爬行和转移一系列过程募集到炎症部位[5-6].已有文献报道,在绿脓杆菌感染时中性粒细胞借助β2整合素募集到感染部位[4],整合素是中性粒细胞执行功能的重要跨膜蛋白[5-6,8].本研究发现,绿脓杆菌感染引起小鼠骨髓中性粒细胞的粘附,同时增强整合素CD11b和CD18的表达.
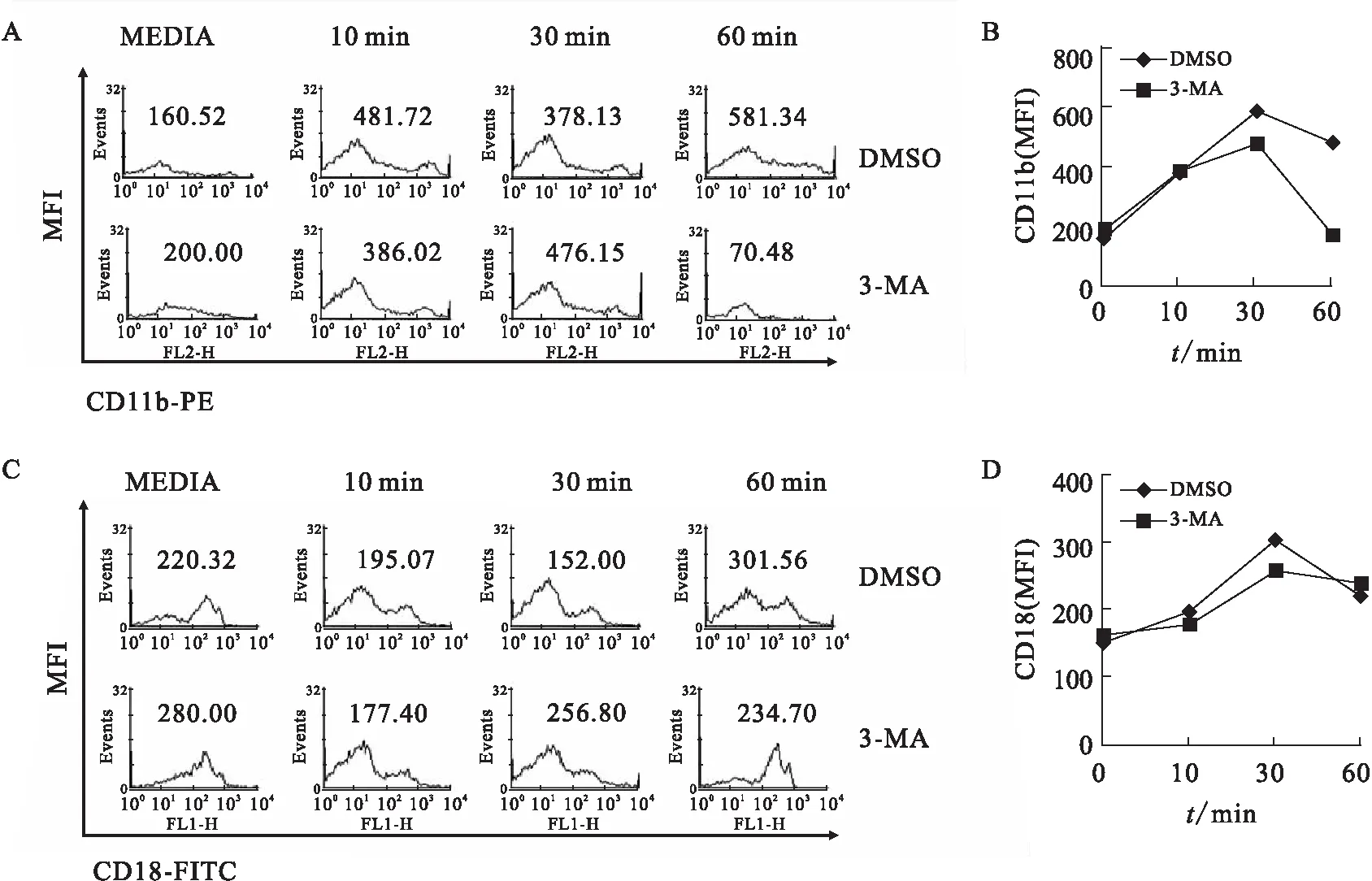
A、B:提取小鼠的骨髓中性粒细胞,每组细胞数量为7.2×105个.3-MA(200 μmol/L)或DMSO预处理后清洗一次细胞,感染绿脓杆菌(MOI 25).收集各组细胞后通过流式细胞术检测CD11b的荧光强度,并制作折线图观察变化趋势.实验重复3次.
C、D:提取小鼠的骨髓中性粒细胞,每组细胞数量为7.2×105个.3-MA(200 μmol/L)或DMSO预处理后清洗一次细胞,感染绿脓杆菌(MOI 25).收集各组细胞后通过流式细胞术检测CD18的荧光强度,并制作折线图观察变化趋势.实验重复3次.
图5 自噬调节绿脓杆菌感染时整合素的表达
Fig.5 Autophagy regulates the expression of integrin duringPseudomonasaeruginosainfection
自噬在肿瘤细胞中激活整合素,并促进细胞的转移[12-13].在中性粒细胞中自噬可以促进细胞脱颗粒、分化、杀菌等作用[14-16],但对中性粒细胞募集的影响还没有文献报道.已有研究发现,自噬可以促进中性粒细胞成熟标志CD11b/CD18的表达[16].CD11b/CD18是中性粒细胞迁移和激活必不可少的重要分子[5,20].本研究发现,绿脓杆菌感染引起骨髓中性粒细胞的自噬,而且自噬的抑制剂3-MA显著地抑制细胞CD11b的表达和中性粒细胞的粘附.
综上所述,绿脓杆菌感染可以激活骨髓中性粒细胞的自噬,而且自噬的抑制剂抑制细胞中整合素CD11b的表达和粘附.本研究确定自噬参与中性粒细胞的粘附,但是否促进中性粒细胞的募集尚不清楚,需进一步研究.
猜你喜欢
医学综述(2022年10期)2022-11-28
医学研究杂志(2021年10期)2021-11-26
皮肤病与性病(2021年3期)2021-07-30
口腔医学(2020年6期)2020-12-19
癌变·畸变·突变(2020年4期)2020-08-05
医学新知(2019年4期)2020-01-02
文苑(2018年18期)2018-11-08
家庭影院技术(2018年9期)2018-11-02
中国制笔(2017年2期)2017-07-18
诗林(2016年5期)2016-10-25
