
污泥中蛋白类物质厌氧转化影响因素及其促进策略研究进展
2020-06-09 10:03陈思思杨殿海庞维海董滨戴晓虎
化工进展 2020年5期
陈思思,杨殿海,庞维海,董滨,戴晓虎
(同济大学环境科学与工程学院,上海200092)
蛋白类物质(在污泥中难以分离纯净的蛋白质,一般用有机氮的含量乘以6.25的值来代表蛋白类物质的含量[1])是剩余污泥中的主要有机组分(占有机质的50%~60%)[2],是污泥厌氧消化产沼气的主要底物,其降解率的提升将对剩余污泥厌氧转化效率的提升起到决定性所用。与此同时,蛋白类物质是污泥中氮元素的最主要载体,对蛋白类物质降解转化的研究将促进污泥厌氧消化过程中氮循环的理论研究。此外,蛋白类物质更是剩余污泥中的主要亲水性物质,其在决定污泥脱水性能方面发挥着比多糖和腐殖质类物质更明显的作用[3-4]。最新研究表明污泥的表面亲水指数与污泥胞外蛋白质二级结构中的α螺旋的比例有着较强的正相关性,且污泥的亲水性能相比于胞外蛋白质亲水官能团的含量,更多地取决于亲水官能团的空间分布[4]。因此,明晰剩余污泥中蛋白类物质的厌氧转化机制和限制性因素,将有利于明晰我国污泥厌氧转化的限制机制,从而对其进行针对性强化,这不仅可以从源头上强化我国剩余污泥的厌氧转化效率,完善污泥厌氧消化过程中的氮循环理论,还可能从减少亲水性物质的角度改善其后续的脱水性能。
本文重点归纳了污泥中蛋白质的厌氧转化机制,并在此基础上总结了污泥中蛋白类物质厌氧转化的影响因素和对其定向促进的研究进展,以期为污泥中蛋白类物质的定向厌氧转化提供借鉴与启发。此外,系统性总结了污泥中蛋白类物质的前沿解析方法,旨在为污泥中蛋白类物质的研究提供全面支撑。
1 蛋白质厌氧转化机制
在厌氧消化过程中,蛋白质完全转化为甲烷与二氧化碳的过程可概括为4个过程:①胞外酶分解作用(蛋白质水解阶段);②大分子有机物发酵为有机酸(氨基酸发酵阶段);③发酵中间产物降解为乙酸(产酸阶段);④以乙酸或氢气/二氧化碳产甲烷(产甲烷阶段)。
1.1 蛋白质水解
蛋白质是由氨基酸以“脱水缩合”的方式组成的多肽链经过盘曲折叠形成的具有一定空间结构的物质,可由胞外酶(蛋白酶)水解成其组成的多肽和氨基酸。研究表明,蛋白质在厌氧反应器中的降解机制与在动物体内如瘤胃中的降解机制存在较大差异。在瘤胃中,需由糖类发酵细菌降解蛋白质,而氨基酸的发酵并不能为其提供足够的能量[1-5];在厌氧反应器中,由具有蛋白水解作用的细菌主导蛋白质的降解,这些过程能够为其代谢过程提供足够的能量[6]。同时也有大量研究表明消化污泥中主要的蛋白水解细菌为革兰氏阳性菌,主要为梭状芽胞杆菌属(genusClostridia),同时这些细菌也在氨基酸发酵中发挥着重要作用[6]。
根据水解程度,蛋白质水解可以分为完全水解(得到的水解产物为各种氨基酸的混合物)和部分水解(水解产物是各种大小不等的肽段和单个氨基酸),在这个过程中必须要对其二级结构和一级结构造成一定程度的改变。蛋白质二级结构指它的多肽链中有规则重复的构象,限于主链原子的局部空间排列,不包括与肽链其他区段的相互关系及侧链构象,二级结构主要有α-螺旋、β-折叠、β-转角和无规卷曲,是通过骨架上的羰基和酰胺基团之间形成的氢键维持的,氢键是稳定二级结构的主要作用力[7]。蛋白质二级结构将直接影响蛋白质的稳定性和絮凝特性,研究表明α-螺旋将诱导蛋白质的稳定性,促进活性污泥的聚集性和絮凝性;而反平行的β-折叠和无规卷曲结构的大量存在将会减弱活性污泥的稳定性与聚集性[8-10],蛋白质的稳定性与聚集性越强,酶或微生物等与其接触对其利用就越困难。蛋白质的一级结构是指蛋白质多肽链中氨基酸的排列顺序,以及在蛋白质分子中起到稳定肽键空间结构作用的二硫键的位置,这是由遗传信息决定的。对蛋白质一级结构的破坏、释放肽链中的氨基酸降有利于酶或微生物等对其利用进而进行氨基酸发酵过程[11]。
1.2 氨基酸发酵
各类氨基酸在分子量大小和结构上存在着很大差异,对其降解也根据类型和浓度的不同存在着多种代谢途径和相应的代谢产物。这些代谢产物包含各种不同的有机物(主要为短链和支链有机酸)、氨氮、二氧化碳以及少量氢气和含硫化合物。氨基酸发酵已经得到大量文献报道[12-17],通过分离微生物种群,在生化和生理方面对氨基酸发酵进行了大量的深入研究。
氨基酸降解主要通过两种途径:①成对的氨基酸可通过Stickland反应得到降解;②单一氨基酸也能通过另外一种过程得到降解(非耦合降解),该过程需要氢利用型细菌的存在。其中Stickland反应是氨基酸分解的常见过程,通常需要一个氨基酸作为电子供体(其产物由于少一个碳原子而短于原氨基酸),同时需要另一个氨基酸作为电子受体(其产物中碳原子数量与原氨基酸相同)。一些特定的氨基酸既能作为电子供体,也能作电子供体(如亮氨酸、苯丙氨酸、酪氨酸、色氨酸和精氨酸)。Stickland反应是氨基酸发酵最简单的方式,且每转化1mol 的氨基酸将会为细胞提供约0.5mol 腺嘌呤核苷三磷酸(ATP)[12]。与非耦合氨基酸分解相比,通过Stickland反应的分解过程更加迅速[13]。
各类氨基酸厌氧发酵细菌如表1 所示。依据Mead[17]和McInerney[6]的研究,这些细菌依据他们参与的Stickland反应以及利用氨基酸的种类主要被分为5类。表1中也依据Hippe[14]的研究总结了每种细菌所产生的酶以及这些细菌的特点。第一类细菌为主导Stickland反应的微生物,在发酵过程中,他们都能够利用脯氨酸产生中间产物包括δaminovalerate、α-aminobutyrate 或γ-aminobutyrate。参与Stickland反应的常见氨基酸包括脯氨酸、丝氨酸、精氨酸、精氨酸、甘草、亮氨酸、异亮氨酸、缬氨酸、丝氨酸、赖氨酸、丙氨酸、半胱氨酸、蛋氨酸、三氨酸、苯丙氨酸、酪氨酸和色氨酸。
氨基酸发酵细菌中不能进行Stickland反应的微生物如表1中二、三、四、五类所示,其中主要包括严格的产孢菌类(梭状芽胞杆菌种Clostridialspecies) 以及不产孢子的专性厌氧微生物如Peptostreptococcus(Micrococcus)等。其他也能够降解氨基酸的微生物如Campylobacterspp. 、Acidaminococcusfermentans、Acidaminobacter hydrogenoformans、Megasphaera elsdenii、Eubacterium acidaminophilum以及一些硫酸盐还原菌[18]。第二类细菌都能利用甘氨酸,有些种也能够利用精氨酸、组氨酸、赖氨酸。第三类细菌都能利用组氨酸、丝氨酸和谷氨酸,有些种类也能够利用精氨酸、天冬氨酸盐、苏氨酸、酪氨酸和色氨酸。第四类细菌只包含利用丝氨酸和苏氨酸的C.putrefaciens。第五类细菌只包含能利用丙氨酸,丝氨酸,苏氨酸和半胱氨酸的C. propionicum。以上四类细菌都不能产生在Stickland 反应中所发现的δ-aminovalerate。在混合环境、存在混合氨基酸的条件下,非耦合的氨基酸降解只能在作电子受体的氨基酸缺乏时才能够发生,对于酪蛋白、白蛋白和明胶等蛋白质,经非耦合过程降解的氨基酸只占全部氨基酸降解量10%以下[19]。
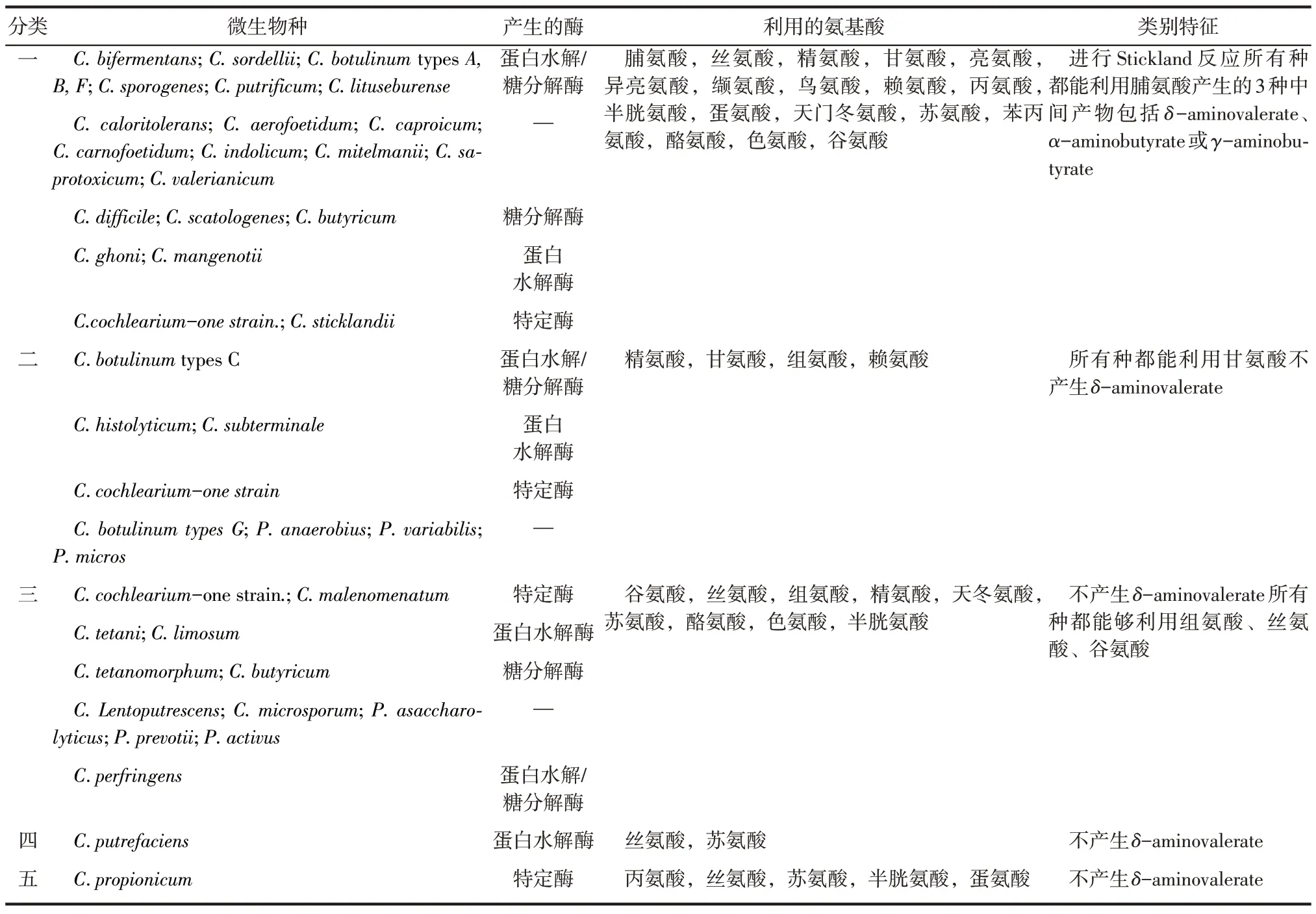
表1 降解氨基酸的厌氧细菌汇总
2 污泥中蛋白类物质厌氧转化的影响因素
Tian 等[20-21]根据X 射线光电子能谱分析图谱分析将污泥中含氮物质分为蛋白质、无机氮、吡咯和吡啶等四类。在复杂的污泥厌氧消化系统中,蛋白类物质的降解率已有部分文献报道。一些研究主要报道了厌氧消化后污泥中蛋白类物质与多糖的降解率,Pinnekamp[22]发现污泥经厌氧消化后,蛋白类物质和多糖的降解率分别为39% 和52%;在Bougrier 等[23]的研究中蛋白类物质和多糖的降解率分别为35%和50%;Chen 等[2]发现脱水污泥经SRT为20 天的中温厌氧消化后,蛋白类物质和多糖的降解率分别为46.2%和49.4%。可见污泥中的蛋白质的厌氧消化降解率普遍较低,由于蛋白类物质为污泥中最主要的有机成分,因此其较低的降解率将主要限制污泥的厌氧消化效率。
在系统运行过程中,蛋白质的降解主要受到基质中其他有机物尤其是多糖的影响、代谢产物氨氮的影响以及其来源的影响,相关研究归纳见表2。Breure 等[24]研究了高浓度糖类(最高可达5g/L)对明胶的水解和酸性发酵的影响(最高可达5g/L),在明胶培养基达到稳定状态后添加相对高浓度的葡萄糖或乳糖作为二次基质,发现在基质中碳水化合物的浓度升高时蛋白质的降解会逐渐减慢,明胶的降解率在其单独发酵和有碳水化合存在的条件下分别为90%和77%;Tommaso 等[25]研究了碳水化合物对牛血清蛋白厌氧消化效率的影响,结果发现尽管添加碳水化合物后系统运行更加稳定,但其中蛋白质的降解率下降了8.8%。Yang 等[26]研究了其中蛋白类物质与多糖的厌氧消化动力学,发现在污泥中多糖比蛋白类物质先降解,且其效率高于蛋白质,厌氧消化前3天由于多糖的快速降解抑制了蛋白酶的形成,使得蛋白质的降解处于停滞期,停滞期后蛋白质的降解与多糖一样都遵循一级动力学,且分为快速降解和慢速降解两个阶段。鉴于多糖对蛋白质降解的抑制作用,在基质中蛋白质和多糖同时存在的情况下,有研究从碳氮比的角度对蛋白质与多糖在基质中的最佳配比做了研究,发现蛋白质与多糖的比例为8∶2时能够达到最佳的共消化效果[27]。氨氮是蛋白质代谢的重要终产物,其含量过高时会对系统内的细菌与古菌都产生毒害作用,从而降低系统的水解酸化以及产甲烷效率[28-29],在Li 等[28]的研究中也证明了在氨氮浓度升高的情况下,与氨基酸代谢相关的功能基因受到了抑制;在Chen 等[30]的研究中对氨抑制的缓解做出了研究,发现系统经驯化恢复之后其蛋白质的降解会得到恢复。研究还发现,在Wagner 等[31]的研究中,酵母提取物和络蛋白氨基酸的降解产气率远高于牛奶中蛋白,特定的蛋白来源中,精氨酸有着最高的降解产气率,且其降解基本不受碳氮比的影响。由此可见由系统运行、物料配比以及代谢产物等因素带来的对蛋白类物质厌氧转化的限制作用,通过系统的优化和调控,都已被报道可得到解决或缓解。
此外,污泥中蛋白质的降解还会受到污泥中无机物如微细砂(二氧化硅)[32]和金属离子的影响[33]。Dai 等[32]研究表明微细砂会与蛋白质形成蛋白-微细砂结合体,导致有机质的酶结合位点减少、表面位点密度下降、表观活化能的上升,导致污泥中蛋白质降解率降低;蛋白质中的羧基是金属离子与有机物络合的重要桥连基,金属离子与蛋白质的络合将限制其的水解及其后续降解[34-35]。
3 污泥中蛋白类物质厌氧转化的促进研究
在完善厌氧消化系统运行、调控其内部因素的基础上,大量学者开发了一系列预处理措施如酸碱预处理[36]、超声预处理[37]、生物预处理[38]、热水解预处理[39]等改善有机组分的溶出性能以提高其中蛋白质的降解率,强化污泥厌氧消化效率,相关研究总结见表2。

表2 污泥中蛋白类物质厌氧转化的影响因素及促进研究总结
研究表明破坏蛋白质折叠构型是水解的必要步骤,打破蛋白质的氢键网络与折叠构型也将有助于蛋白质水解及利用效率的提高[43]。Devlin 等[36]发现经酸预处理后蛋白质与多糖的降解率分别提高了7%和9%;Appels 等[44]研究发现低温热水解(60~90℃)未能明显促进这两类有机物的降解;Feng等[40]研究发现添加零价铁后污泥中蛋白类物质与多糖的降解率分别提高了15%和34.1%;Shao 等[41]发现好氧消化相比于厌氧消化更能够实现蛋白类物质的有效降解;Xiao等[42]对比研究了酸、碱、紫外照射预处理改变蛋白质构象对提高蛋白质废水发酵产氢气的效果,结果表明使用紫外线照射能有效达到破坏蛋白质氢键及其构型的效果,对其水解起到了促进作用;Chen 等[2]发现经高温热水解(160℃)后脱水污泥中蛋白类物质的降解率提高了28.9%。
可见针对蛋白类物质的赋存状态与结构强化其厌氧转化的策略目前处于多种方法进行尝试的阶段,在以上强化措施中,效果尚可且较容易工程推广的技术为高温热水解技术,这一技术已被研究表明能够增强蛋白质的溶解,且其对污泥厌氧消化性能的强化作用得到了大量的工程验证[45]。然而由于污泥泥质的较大差异和强化机制研究的缺乏,其促进效率仍参差不齐[39,46-47],高温热水解在突破我国剩余污泥厌氧消化瓶颈上是否具有关键性作用也亟待研究。
4 污泥中蛋白类物质的解析方法
4.1 氨基酸的测定方法
在众多的氨基酸测定方法中,沼液中总氨基酸量的测定主要通过茚三酮比色法[48],而沼液中的氨基酸组分的测定,主要以气相色谱法[49]和高效液相色谱法[50]为主。由于大多数氨基酸无紫外吸收和荧光发射特征,为提高分析检测灵敏度和分离选择特性,通常需将氨基酸进行柱前或柱后衍生化,并选用阳离子交换或反相液相色谱法对其进行分离并经紫外或荧光检测(OPA/FMOC-Cl/RT-HPLC)来实现各组分的测定[51],能使氨基酸产生荧光的衍生剂有邻苯二甲醛(OPA)、9-氯甲酸芴甲酯(FMOCCl)、丹酰氯(DANSYL-Cl)等。沼液中溶解游离氨基酸的含量较低,并且沼液中复杂的其他成分以及所包含的许多细小固体悬浮物会对衍生化过程产生干扰,如高浓度氨氮和挥发性胺容易与某些待测氨基酸形成荧光副产物,干扰氨基酸的分析与测定。孟庆国等[49]采用气相色谱法测定了沼液中的18 种溶解游离氨基酸,然而,该研究中所采用的分析方法较为繁琐,且未涉及样本加标回收率、精密度等参数的进一步分析。氨基酸是一类包含至少一个羧基和一个氨基官能团的化合物(等电点2.8~10.8),它在不同的pH条件下,会以阳离子、阴离子和中性分子的形式存在。因此,李建华等[52]研究开发了pH调节(pH=10.2)旋蒸浓缩联合超滤离心(3K Millipore)的前处理新方法,并在此基础上进一步采用OPA-FMOC-Cl 作为柱前衍生化试剂,使用反相高效液相色谱对沼液中的溶解游离氨基酸进行分析,并通过外标法对所开发的沼液中溶解性氨基酸分析方法进行了评估。结果表明,与传统的SCX-SPE 小柱固相萃取方法相比,所使用的新预处理方法因其较高的回收率和精确度及操作的简便性而更适用于沼液中溶解游离氨基酸的测定,并且避免了SCX-SPE 法对氨基酸选择性吸附和对其他干扰物的非选择性吸附而导致的部分氨基酸损失过大的现象。
4.2 荧光光谱法
污泥液相中有机物初始状态及被微生物降解代谢过程中的络氨酸、蛋白质、类富里酸和类胡敏酸等在紫外光的激发下会产生特征荧光,显示出其组成和官能团结构的大量信息[53]。三维荧光光谱法(excitation-emission-matrices,EEM)因具有快速性、选择性与敏感性,被广泛应用于对蛋白质和腐殖酸等物质的检测中[54],然而原始谱中重叠的荧光团和荧光带经常影响对物质性质的解析与测定。平行因子分析法(parallel factor analysis,PARAFAC)可以将荧光信号分解为相对独立的荧光现象而加以鉴别,从而提高了准确性,将平行因子分析引入三维荧光光谱分析是对该技术的重要提升[55]。运用PARAFAC 分析后,溶解性有机物中的组分一般被分为两大类:类腐殖质物质和类蛋白质物质。依据各类有机物的最大激发和发射波长,溶解性有机物中能被鉴别出的物质有类络氨酸、类色氨酸、类蛋白质、不同种类的类腐殖质如富里酸等[56]。Li 等[57]采用EEM-PARAFAC 表征了脱水污泥中有机物在厌氧消化过程中的迁移转化,发现其上清液中的荧光类物质包括类络氨酸、类色氨酸以及类腐殖质类物质含量逐渐上升,且蛋白质的含量发生了明显的快速降低。
4.3 宏蛋白组学
由于污泥中的主要蛋白类物质成分来源于微生物细胞内或其分泌的胞外聚合物,且污泥的厌氧消化过程是由微生物主导,而蛋白质是微生物生命活动的主要承担者,因此从蛋白质信息出发分析微生物功能与代谢途径将是解析污泥中蛋白质降解途径的必要条件。宏蛋白组学(metaproteome,或称元蛋白质组学)从宏基因组学(将群落中所有微生物DNA 提取出来研究)的概念中提出,指环境混合微生物群落中所有生物的蛋白质组总和[58]。其功能与优点在环境生物学分析中主要体现在以下3个方面:①可较准确地反映活性污泥微生物脱氮除磷、去除有机物代谢活动,还可用于跟踪新的功能基因、代谢途径中或者在环境胁迫下微生物产生的特征性蛋白质[59];②可以整体把握活性污泥微生物的功能及其动态,是理解宏代谢组学(生态系统中所有微生物的代谢产物)的关键步骤[60];③宏蛋白质组学可以为研究活性污泥功能性微生物设计特异性探针,在线鉴定和定量分析特定群落微生物表达的蛋白质,推动活性污泥微生物种群多样性的研究发展[61]。Abram 等[62]首次将宏蛋白质组学用于厌氧消化(AD)系统中的颗粒污泥微生物研究,结果表明在低温(15℃)环境下,以葡萄糖为基质的合成废水厌氧消化过程中,检出的18 个蛋白质中有14个与糖酵解和产甲烷作用代谢功能密切相关,为低温下AD系统处理废水过程中生物降解转化途径的研究提供了理论依据。Hagen 等[63]采用宏蛋白组学解析了高温厌氧消化系统中的细菌与古菌分别的产酸与产甲烷代谢途径、参与中间产物代谢的主要蛋白质以及脂肪酸的代谢网络。因此,采用宏蛋白组学对于厌氧消化过程中有机物的代谢途径,尤其是对蛋白质及其重要的代谢功能的表征具有一定的可行性。
5 结语与展望
蛋白质的厌氧降解主要包括蛋白质水解与氨基酸代谢两个重要步骤,蛋白质的结构与氨基酸组分差异将决定其厌氧转化性能和持水能力。污泥胞外蛋白质二级结构中α-螺旋的比例与污泥的表面亲水指数呈现显著正相关关系。蛋白质来源、污泥中其他有机物如多糖、污泥中无机物如微细砂(二氧化硅)和金属离子、以及代谢产物如氨氮等都会对污泥中蛋白类物质的厌氧转化性能带来显著的影响。这些影响通过一定的预处理手段可以得到缓解,研究发现破坏蛋白质的构象、二级结构与氢键网络等将有效强化其厌氧转化性能,且在众多策略中,高温热水解技术发挥了有效的作用并得到了广泛应用。然而对污泥中蛋白类物质的研究仍然停留在定性阶段,鲜有准确的定量转化研究。通过对污泥中蛋白类物质的前沿解析方法包括氨基酸测定方法、荧光光谱法和宏蛋白组学的归纳与总结,可以在一定程度上推动污泥中蛋白类物质的研究,促进其定向转化研究,从而强化其厌氧转化性能以及消化产物的脱水性能。
猜你喜欢
军事文摘(2022年16期)2022-08-24
食品安全导刊(2021年21期)2021-08-30
粉末冶金技术(2021年1期)2021-03-29
法制博览(2020年27期)2020-11-30
科学(2020年6期)2020-02-06
中学化学(2019年3期)2019-07-08
科技与创新(2018年21期)2018-11-30
中学化学(2016年2期)2016-05-31
课程教育研究·下(2016年2期)2016-03-25
专用汽车(2015年2期)2015-03-01
