
黔北桐梓地区奥陶系湄潭组下部腕足动物的组合特征
2020-06-08 22:44王冬梅杨宇宁黎斌钱庆环
贵州大学学报(自然科学版) 2020年3期
王冬梅 杨宇宁 黎斌 钱庆环


摘 要:黔北桐梓红花园剖面是华南扬子台地经典的奥陶系剖面之一,其中的湄潭组下部泥页岩段产出大量的腕足类化石。在前人研究的基础上,形态分析显示:红花园剖面湄潭组下部的腕足类化石共有27属,其中以正形贝类Sinorthis的丰度最高,为湄潭组下段腕足动物中的优势类型。此外,Paralenorthis,Desmorthis,Xinanorthis等也比较常见,同为早奥陶世扬子台地近岸浅水环境下Sinorthis动物群的共生腕足分子。通过与黔中乌当地区湄潭组下段的腕足动物组合进行多样性比较,结果显示:桐梓红花园剖面的腕足动物,无论是化石丰度还是形态分异度,都明显高于贵阳乌当地区。两地之间湄潭组沉积环境的差异和研究程度的不同可能是腕足动物化石面貌不一样的主要原因。红花园剖面湄潭组下部腕足动物的多样性迅速增加,为研究华南早奥陶世腕足动物演化史上首次辐射事件提供了新的证据,也为探讨该地质时期黔中古陆南、北缘近岸环境下腕足动物的地理区系划分提供了重要信息。
关键词:腕足动物;组合特征;湄潭组;早奥陶世;红花园剖面
中图分类号:Q915.816+.4
文献标识码: A
腕足动物是一类单体群居的海生底栖、身体构型呈两侧对称的无脊椎动物[1]。作为早古生代海洋中生物群落的重要组成部分[2],腕足动物可为研究奥陶纪时期海洋动物的生态系统和辐射演化提供信息与证据。遵义桐梓红花园剖面位于桐梓县以南红花园村的东侧(起点坐标为28.06°N,10685°E),是华南扬子台地典型的奥陶系剖面之一,也是贵州研究奥陶系最为详细的剖面。红花园剖面地层出露良好,地层序列较为齐全,奥陶系自下而上出露有桐梓组、红花园组、湄潭组、宝塔组和五峰组[3]。其中,湄潭组出露完整,厚达258 m,下部泥页岩段含有大量的腕足化石。早在1984年,张文堂等系统地描述了湄潭组的8个腕足动物属[4]。詹仁斌等于2001年重新测制了红花园剖面,并逐层采集化石,鉴定出湄潭组有48个腕足动物属[5],比较完整地揭示了红花园剖面湄潭组中腕足动物的基本组成面貌。此外,焦惠亮先后对红花园剖面湄潭组的部分腕足化石進行了摘录报道[6]。
结合前人的工作基础和本次野外化石采集的结果,本文鉴别出桐梓红花园剖面湄潭组下部的腕足动物共有27属之多,显示出很高的形态分异度。但是,奥陶纪时期地处黔中古陆南缘的贵阳乌当地区,其湄潭组下部腕足动物仅发现10属[7],分异度和丰度都远低于黔中古陆北缘的桐梓红花园地区。基于奥陶纪贵州这一独特的岩相古地理格架,对比桐梓红花园和贵阳乌当两个沉积区湄潭组下部腕足动物的组合特征,对于揭示早奥陶世黔中古陆南、北缘之间腕足动物的组合面貌,以及华南板块腕足动物演化史上首次辐射演化事件具有重要意义。此外,红花园剖面湄潭组下部腕足动物的多样性增加,反映了华南早、中奥陶世腕足动物的演化特征,证实正形贝目呈现出爆发式的增长模式。
1 地质背景
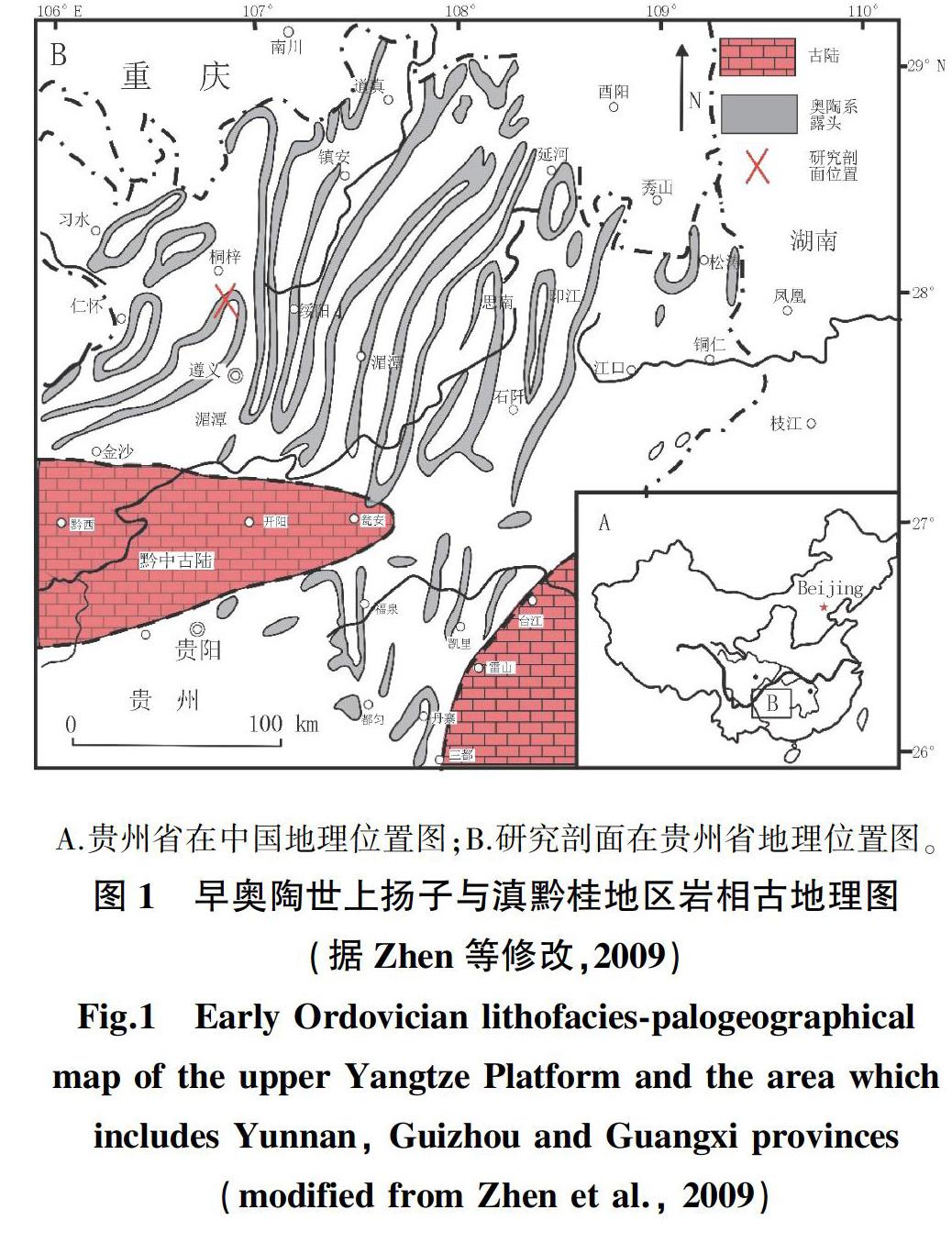
贵州中部地区在奥陶纪时期隆起上升为黔中古陆[8],围绕这一古陆的周边往往发育有比较齐全的奥陶纪近岸浅水相地层,桐梓红花园剖面则是黔中古陆北缘离海岸线不远的台地相地层序列(图1)。
湄潭组是红花园剖面中一套厚度较大的碎屑岩沉积,厚达258 m,主要为泥、页岩沉积,自下而上可分为3段:第一段厚110.78 m,以黄、灰绿及蓝灰色页岩为主,夹少量粉砂质粘土岩及砂质页岩;第二段厚24.64 m,以中厚层微至粗晶灰岩为主,夹少量砂质页岩及钙质粉砂岩;第三段厚122.87 m,以黄灰色砂质页岩为主,夹薄层砂岩、灰黑色页岩及灰色薄层泥质灰岩[9](图2)。本文研究的腕足化石采自湄潭组下部(相当于第一段),该岩性段以泥、页岩居多,含有丰富的腕足、笔石、三叶虫化石,与下伏红花园组呈整合接触,地层分界清楚。
前人研究结果显示,黔北和黔中地区奥陶系湄潭组下部的笔石生物地层相似,依据其笔石化石的产出序列,自下而上可分为Acrograptus filiformis带、Didymograptellus eobifidus带、Corymborgraptus deflexus带、Azygograptus suecicus带4个笔石带[10-11](图2)。本文所研究的桐梓红花园剖面湄潭组下部地层包括上述4个笔石带,且其顶界位Azygograptus suecicus笔石带的中部。因此,目标岩性段年代地层属于下奥陶统弗洛阶的地层[12]。
2 桐梓红花园剖面湄潭组下部的腕足动物化石
2.1 化石的保存方式和埋藏学特征
桐梓红花园剖面湄潭组下部采集的腕足大部分保存比较完整,破碎的化石较少,绝大多数腕足化石壳体的纹饰保存清晰。此外,化石多呈杂乱分布,没有定向性排列,而且大小不一,不具分选现象,其中有不少化石重叠在一起保存的现象,且以个体较小的标本居多(图3)。需要提及的是,湄潭组近底部有一个腕足化石的密集层,其中的腕足化石数量庞大,据不完全统计,化石密集层10 cm×10 cm范围内化石个体数达122 个,很多兼有腹壳和背壳聚集埋藏(图3)。
湄潭组下段的腕足动物化石的埋藏方式大致有以下3种(图4):(1)背、腹壳两相连接保存;(2)背壳或腹壳单独保存;(3)群体保存。其中,第三种埋藏方式可进一步分成两种保存状态:一是同属或不同属的个体大小不一,多个背壳、腹壳一起保存,但每个壳体多是独立的;二是大小不一的背壳、腹壳绞合在一起保存,而且常有较大的个体盖住较小个体的现象。据此,桐梓红花园剖面湄潭组下部的腕足化石呈现出快速原地埋藏或者近原地埋藏保存特征。
2.2 腕足动物化石的组合特征
结合前人资料[3]及本次研究工作,目前红花园剖面湄潭组下部的腕足動物共有27个属,分别为Nocturnellia,Euorthisina,Desmorthis,Nothorthis,Pseudomimella,Paralenorthis,Obolus,Sinorthis,Tritoechia,Anchigonite,Taphrorthis,Yangtzeella,Pseudoporambonites,Schedophyla,Palaeoglossa,Tarfaya?,Lingulella,Xinanorthis,Stichorophia,Ldiostrophia,Apatobolus,Nereidella,Leptella,Punctolira,Ferrobolus,Fenxiangella和Trondorthis。统计显示:这些腕足动物类群以中华正形贝Sinorthis数量最多,占所采标本总数的54.20%;其次为Paralenorthis,占所采标本总数的11.20%;此外,Desmorthis和Xinanorthis的数量也比较多,分别占所采标本总数的8.50%和7.10%(图5)。
分析可知,Sinorthis是湄潭组下段腕足动物中的优势种,Sinorthis(图6A—H)、Paralenorthis(图7C)、Desmorthis(图7I—J),Xinanorthis(图7E)为湄潭组下段的主要腕足分子, 一共占腕足动物总数的80%以上。这几个化石属都是扬子台地Sinorthis动物群的常见类型。这些腕足动物生活底质主要为泥质、粉砂质软泥, 处于相对浅水的陆棚区[7]。桐梓红花园地区在早奥陶世弗洛期(湄潭组沉积期)离黔中古陆北缘较近(图1),为典型的浅水陆棚环境,为Sinorthis动物群的适应辐射创造了先决条件。
在湄潭组下部4个笔石带中,Didymograptellus eobifidus带内的腕足动物化石属数量最多,为 23
属,而Azygograptus suecicus带中的腕足最少,只有9属。此外,Nocturnellia,Euorthisina,Nothorthis,Obolus,Sinorthis,Tarfaya?在上述4个笔石带中都有分布,具有较广的地层延限。不过,湄潭组自D.eobifidus 带往上,腕足动物的属种分异度明显降低。
3 研究区与黔中贵阳乌当地区湄潭组下段腕足动物的特征对比
3.1 腕足类组合特征的比较
桐梓红花园与贵阳乌当湄潭组下段腕足动物均属于扬子台地的Sinorthis动物群,两地占主导地位的腕足动物属种一致,主要组成分子均为Sinorthis,Paralenorthis,Xinanorthis,Lingulella,Leptella,但两地之间腕足动物的属种类型和多样性存在差异。比如,就生物多样性而言,桐梓红花园剖面湄潭组下段的腕足动物共有27个属,而贵阳乌当的大洼剖面与小谷农剖面湄潭组下段腕足动物共计10个属[7],可见两地的腕足动物多样性明显不同。
此外,不论桐梓红花园地区还是贵阳乌当地区,湄潭组下段自下而上的Acrograptus filiformis带、Didymograptellus eobifidus带、Corymborgraptus deflexus带和Azygograptus suecicus带中[10-11],红花园剖面各笔石带内产出的腕足动物,其属种分异度都远大于贵阳乌当地区的相应笔石带。不过,两个地区腕足动物分异度的变化趋势是一致的,都是在D. eobifidus带达到顶峰(表1)。
3.2 两地之间腕足动物多样性差异的原因
桐梓红花园地区与贵阳乌当地区湄潭组下部的腕足动物组成面貌差异明显,造成这种差异的原因可能有以下几种:
(1)两地所处的古地理位置处于不同的海洋环境,其沉积相不同。早奥陶世时期,桐梓红花园位于黔中古陆北缘,属浅海陆棚相,而黔中古陆南缘的贵阳乌当属滨岸—陆棚过渡相[12]。相比之下,浅海陆棚环境以海水循环良好,海底地势平坦,阳光及氧气充足为特点,有利于底栖动物的大量繁殖。这可能是红花园地区湄潭组下部的腕足动物明显高于贵阳乌当地区的主要原因。
(2)分异演化的不同步性。差异演化是奥陶纪生物大幅射的基本特征之一[13]。不同古地理位置、不同地层剖面上的腕足动物的分化具有不同步性[14]。这可能也是桐梓红花园与乌当地区之间湄潭组腕足动物的组成面貌存在明显差异的原因之一。
(3)研究程度不同也是一个重要因素。红花园剖面是贵州研究奥陶系最为详细的剖面。长期以来,很多学者对剖面进行了非常系统的研究,无论是化石采集力度还是研究程度都远远超过贵阳乌当地区。这也是目前导致两地腕足动物的组合特征差异显著的潜在原因。
4 演化意义
奥陶纪时期,华南扬子板块具有比较齐全的奥陶系地层序列和得天独厚的生物化石记录。综合研究显示,华南早、中奥陶世腕足动物的演化具有以下几个特征:(1)腕足动物多样性自奥陶纪特马豆克阶下部开始增加,在Acrograptus filiformis带成种速度加快,到Didymograptellus eobifidus带达到第一次峰值。红花园剖面的腕足动物化石记录很好地验证了这一辐射趋势。(2)桐梓红花园与贵阳乌当之间湄潭组下部腕足动物组合特征的差异性表明,腕足动物的形态分异可因古地理位置的不同而存在不同步性。(3)不同环境下的腕足动物群强烈分化[14]。
就属一级分类单元而言,桐梓红花园剖面湄潭组下段的腕足动物在D. eobifidus带到达峰值,有23属之多,之后分异度逐渐降低。乌当地区的分异度变化趋势与其相同(图8)。
红花园剖面湄潭组的腕足动物群组成(Sinorthis动物群)、化石保存以及地层沉积特征都显示出近岸浅水的特点。红花园剖面同乌当地区湄潭组下段腕足动物的分异度变化印证了华南早、中奥陶纪腕足动物的演化特征,记录了正形贝目的爆发式增长。桐梓红花园地区与贵阳乌当地区湄潭组下部的腕足动物组成面貌的差异表明,早奥陶世不同环境下腕足动物的形态分异具有不同步性。比较Nereidella、Leptella与Euorthisina等的延限位置与其他地区的差异,可以为研究华南奥陶纪早期的腕足动物的辐射控制因素提供证据[15-16]。
5 结语
(1)桐梓红花园剖面湄潭组下段的腕足動物共计有27个属,其中Sinorthis的化石数量最多,为湄潭组下段腕足动物中的优势种群,Paralenorthis,Desmorthis,Xinanorthis等也比较常见,同为早奥陶世扬子台地近岸浅水环境下Sinorthis动物群的共生腕足分子。
(2)桐梓红花园湄潭组下段的腕足化石有3种埋藏方式:背壳或腹壳单独保存、聚集保存以及背、腹壳两相连接保存。据此推测为快速原地埋藏或者近原地埋藏。
(3)桐梓红花园地区与贵阳乌当地区的湄潭组下段腕足动物均属于扬子台地的Sinorthis
动物群,两地占主导地位的腕足动物类型一致,但属种类型及多样性存在差异。这种现象可能是两地古地理位置不同导致岩性及沉积环境的差异造成。两地之间湄潭组中腕足动物的分
异度变化趋势相同,均在Didymograptellus eobifidus带中最高,往上分异度降低。
致谢:研究过程中得到贵州大学博士生陈争鹏的大力帮助,在此致以谢忱。
参考文献:
[1]童金南,殷鸿福. 古生物学[M]. 北京:高等教育出版社, 2007:110.
[2]张志飞. 早寒武世澄江化石库腕足动物研究[D]. 西安:西北大学,2006.
[3]李军,王怿,RAINER B. 贵州桐梓奥陶系十字铺组疑源类[J]. 微体古生物学报, 2000,17(1):30-38.
[4]许汉奎. 滇西西部中奥陶统施甸组的腕足类[J]. 古生物学报,2016,55(3):291-313.
[5]ZHAN R B,JIN J S. Ordovician-Early Silurian(Llandovery)stratigraphy and palaeontology of the Upper Yangtze Platform, South China[M]. Beijing:Science Press, 2007:39-18.
[6]焦惠亮. 奥陶系[M]//贵州省地质调查院中国区域地质志·贵州志. 北京:地质出版社,2017: 179-219.
[7]孙海静,杨宇宁,彭进, 等. 贵阳乌当早奥陶世湄潭组下段腕足动物组合特征[J]. 贵州大学学报(自然科学版),2011,28(1):23-29.
[8]沈志达,梅冥相,曾羽. 贵州太康运动的地层学效应——兼论“黔中古陆”的形成[J]. 贵州地质,1990,7(2):91-98.
[9]聂瑞贞. 黔中隆起及其周缘奥陶系层序地层格架及古地理演化[D]. 北京:中国地质大学(北京), 2006.
[10]杨宇宁,赵明宇,孙海静,等. 贵阳乌当下奥陶统湄潭组下段笔石带的划分[J]. 古生物学报,2011,50(1):77-91.
[11]张元动,刘晓,詹仁斌. 贵州遵义高桥奥陶系湄潭组下部的笔石[J]. 古生物学报,2007,46(2):145-166.
[12]汪啸风, 陈孝红, 王传尚, 等. 中国奥陶系和下志留统下部年代地层单位的划分[J]. 地层学杂志, 2004, 28(1):1-17.
[13]梅冥相. 论“黔中古陆”[J]. 贵州地质, 1994,11(3):199-206.
[14]詹仁斌, 靳吉锁, 刘建波. 奥陶纪生物大辐射研究: 回顾与展望[J]. 科学通报, 2013, 58(33):3357-3371.
[15]詹仁斌,戎嘉余,程金辉,等. 华南早、中奥陶世腕足动物多样性初探[J]. 中国科学(D辑:地球科学), 2004,34(10):896-907.
[16]ZHEN Y Y, PERIVAL I G, LIU J B, et al. Conodont fauna and biostratigraphy of the Honghuayuan formation (Early Ordovician) of Guizhou,South China[J]. Alcheringa: An Australasian Journal of Palaeontology, 2009,33(3):257-295.
(责任编辑:曾 晶)
