
杨树miR393对FBL基因家族成员调控作用的鉴定
2020-06-06 08:18:16楚立威贺学娇舒文波卢孟柱
中南林业科技大学学报 2020年5期
唐 芳,楚立威,贺学娇,舒文波,卢孟柱
(1.中国林业科学研究院 a.林业研究所;b.林木遗传育种国家重点实验室;c.国家林业和草原局林木培育重点实验室,北京 100091;2.南京林业大学 南方现代林业协同创新中心,江苏 南京 210037;3.华中农业大学 a.教育部园艺植物生物学重点实验室;b.园艺与林业科学学院,湖北 武汉 430070)
microRNAs(miRNAs)是一类长度为21 ~ 22 个核苷酸的负调控非编码小RNA,可以对其特异的靶基因进行转录及转录后负调控[1-2]。大部分miRNAs 是以独立的转录单位存在,由RNA 聚合酶II 转录成的原始转录本(pri-miRNA),可以形成一个发卡结构并被进一步剪切,释放出成熟的miRNAs。植物miRNAs 通过靶向mRNA 进行剪切或翻译抑制,在植物发育和环境胁迫反应中发挥重要的调控作用[3-4]。miRNA 是通过碱基互补配对原则选择靶基因的,因此一个靶基因家族中的成员因具有同源度高的结构域,可能会同时受到相同miRNA 的调控。
大多数miRNA 家族在植物进化过程中具有较高的保守性[5]。miR393 作为植物体中保守的miRNA 家族之一,在拟南芥、水稻、杨树等不同植物物种中均存在。尽管编码miR393 前体的序列在物种内、物种间各有不同,但其成熟体序列相同或只有1 ~2 个碱基的差别[6-8]。拟南芥miR393是由MIR393a 与MIR393b 2 个前体编码而来,但成熟体序列完全相同,它的靶基因是4 个F-box 家族的生长素受体基因(TIR/AFB)[6],miR393 可直接对TIR/AFB转录本进行剪切,起负调控作用[5-6,9]。 在水稻中,miR393 2 个前体编码的成熟体只有1 个碱基长度的差异,且只靶向剪切OsTIR1与OsAFB22 个基因[10-11]。在杨树中,通过小RNA与降解组高通量测序,发现miR393 由4 个前体编码,但成熟体序列完全相同,并且有4 个TIR/AFB同源家族基因可能受到miR393 的剪切[12]。
在不同植物中,受miR393 调控的靶基因都属于TIR1的同源基因。美国Estelle 实验室[13]和英国Leyser 实验室[14]均于2005年同时证实TIR1 是一种生长素受体蛋白,在拟南芥中其属于7 个相关F-box 蛋白基因中一个小的亚家族[15]。该蛋白是在拟南芥对抗生长素运输抑制剂遗传研究时被发现的,故名为“运输抑制应答因子1”(Transport Inhibitor Response Protein 1,TIR1)。有研究证明,拟南芥中存在6 个TIR1基因的同源基因,分别是AFB1、AFB2、AFB3、AFB4、AFB5和COL1,它们作为一个基因家族在拟南芥中起着包括生根、幼苗生长发育、种皮发育在内各不相同的作用[16-18]。 在杨树中也有与拟南芥TIR1家族直系同源的基因家族,即FBL基因家族[19]。该基因家族有8 个成员,其中FBL1对杨树根的发育起到十分重要的作 用[20],而对于其他家族成员的功能研究还尚未有报道。拟南芥的4 个F-box 家族成员TIR1、AFB1、AFB2、AFB3受到miR393 的剪切调控[6],而杨树有8 个同源的FBL基因,它们是否也受到miR393 的调控需要进一步验证。
为揭示杨树miR393 对FBL基因家族不同成员的调控作用,本研究首先从系统进化及基因结构等方面对杨树FBL家族成员进行分析,再通过miR393 与FBL基因家族成员的序列比对、miR393及FBL家族基因在杨树不同发育时期及不同组织中的表达模式和5′-RACE 检测,验证出FBL基因家族8 个成员中只有PtFBL1、PtFBL2、PtFBL3和PtFBL4是miR393 的靶基因。通过揭示miR393对FBL基因家族不同成员调控的差异,为进一步研究mi393-FBL 调控关系奠定了理论基础。
1 材料与方法
1.1 FBL 基因家族的鉴定及物种间的关系
从植物基因组数据库网站Phytozome(https://phytozome.jgi.doe.gov,V12.1)中下载拟南芥、水稻、杨树和小立碗藓FBL家族成员的核酸及氨基酸序列。利用ClustalX2 对其核酸序列和氨基酸序列进行比对[21],并使用MEGA5.0 中的邻近算法构建系统发育树[22],来分析拟南芥、水稻、杨树及小立碗藓FBL基因家族的进化关系。
1.2 杨树FBL 基因结构与motif 分析
利用Gene struture display server(GSDS,http://gsds.cbi.pku.edu.cn)对杨树FBL基因家族8个成员的外显子/内含子结构及miR393 剪切位点进行分析[23]。利用NCBI 上的Conserved Domain Database 软件(CDD,https://www.ncbi.nlm.nih.gov/cdd/)对杨树FBL家族8 个基因的保守结构域进行预测,通过IBS1.0[24]软件对预测结果与miR393 剪切位点进行绘图。
1.3 杨树不同时期及不同组织RNA 的提取及反转录
为了研究杨树miR393 和FBL家族不同基因在不同发育时期及不同组织的表达模式,分别选择84K 杨(Populus alba×Populus glandulosa)半个月组培苗的叶(L-0.5M)、茎(S-0.5M)和根(R-0.5M),1 个月组培苗的叶(L-1M)、茎(S-1M)和根(R-1M),3 个月土培苗的顶端分生组织(SAM-3M)、未展开叶(YL-3M)、第1 至3 片展开叶(ML-3M)、第1 个茎段(N1-3M)、第5 茎段(N5-3M)、第10 茎段(N10-3M)、茎基部(Base-3M)和根(R-3M)为材料,采用LC sciences 总RNA 纯化试剂盒(#TRK-1001,LC sciences)提取上述植物材料的总RNA(包含mRNA、rRNA、miRNA 和其他小RNA)。取1.5 μg 起始量的总RNA,首先通过Poly(A) Tailing Kit(#AM1350,ThermoFisher)将3′ 端 无PolyA的RNA 加上PolyA 尾,再取10 μL 加PolyA 尾的RNA(750 ng),使用SuperScript® III 第一链合成系统(#18080-051,ThermoFisher)进行反转第一链cDNA 的合成,反转录过程按说明书进行,通用反转录引物为5′-AACGAGACGACGACAGA CTTTTTTTTTTTTTTTV-3′。反转录得到的cDNA稀释20 倍后进行后续的定量qRT-PCR。
1.4 实时荧光定量PCR
使 用Primer3(http://frodo.wi.mit.edu/primer3/input.htm)对ptc-miR393 和FBL家族基因进行定量PCR 的引物设计,引物设计原则为20 ~25 bp 长度,退火温度58 ~60 ℃,PCR 产物长度为150 ~250 bp,引物序列见表1,其中PP2A-2为内参基因。定量PCR 反应体系为:10 μL KAPA SYBR FAST qPCR Master Mix(#K4601,KAPA)、2 μL 上述稀释20 倍的cDNA、定量上下游引物(10 μmol )各0.8 μL 和6.4 μL 去离子水,每个样本进行4 个技术重复。在LightCycler 480定量PCR 仪(Roche)上进行定量PCR 的运行,PCR 扩增程序为:95 ℃ 3 s;95 ℃ 10 s,60 ℃ 30 s, 72 ℃ 3 s,重复45 个循环;95 ℃ 3 s。融解曲线分析条件:95 ℃ 5 s(以每s 增加0.11 ℃的速率使65 ℃增至95 ℃)。

表1 杨树FBL 基因家族定量PCR 引物序列Table 1 Quantitative PCR primer sequences of poplar FBLgene family
1.5 miR393对杨树FBL家族基因剪切的5′-RACE验证
为了检测ptc-miR393 对杨树FBL家族基因的剪切情况,需要通过5′-rapid amplification of cDNA ends(5′-RACE)实验对杨树8 个FBL基因分别进行剪切位点的验证。首先,在ptc-miR393 对FBL1-8基因可能的剪切位点下游设计2 条基因特异的巢式引物(表2),之后分别与上游SMART II A 接头上的通用outer 引物(5′-ctaatacgactcactat agggcAAGCAGTGGTATCAACGCAGAGT-3′) 和inner引物(5′-ctaatacgactcactatagggc-3′)进行两轮巢式PCR 的扩增。本实验以84K 杨组培苗为材料,采用LC sciences 总RNA 纯化试剂盒(#TRK-1001,LC sciences)提取组培苗全株的总RNA,再通过SMART ™ RACE cDNA Amplification Kit(Clontech,TaKaRa)反转录出带SMART II A 接头的第一链cDNA。将cDNA 稀释10 倍后,利用Takara EX Taq(#RR902A,TaKaRa) 进行巢式PCR 的第一轮反应,取1 μL 第一轮产物作为模板进行巢式PCR 的第二轮反应。第二轮PCR产物通过琼脂糖凝胶电泳后,切取目标片段,经回收纯化后连接到PMD19-T 克隆载体(#6013,TaKaRa),转化DH5α 感受态大肠杆菌,涂板后37 ℃过夜培养,每个菌板挑15 个单克隆进行测序。
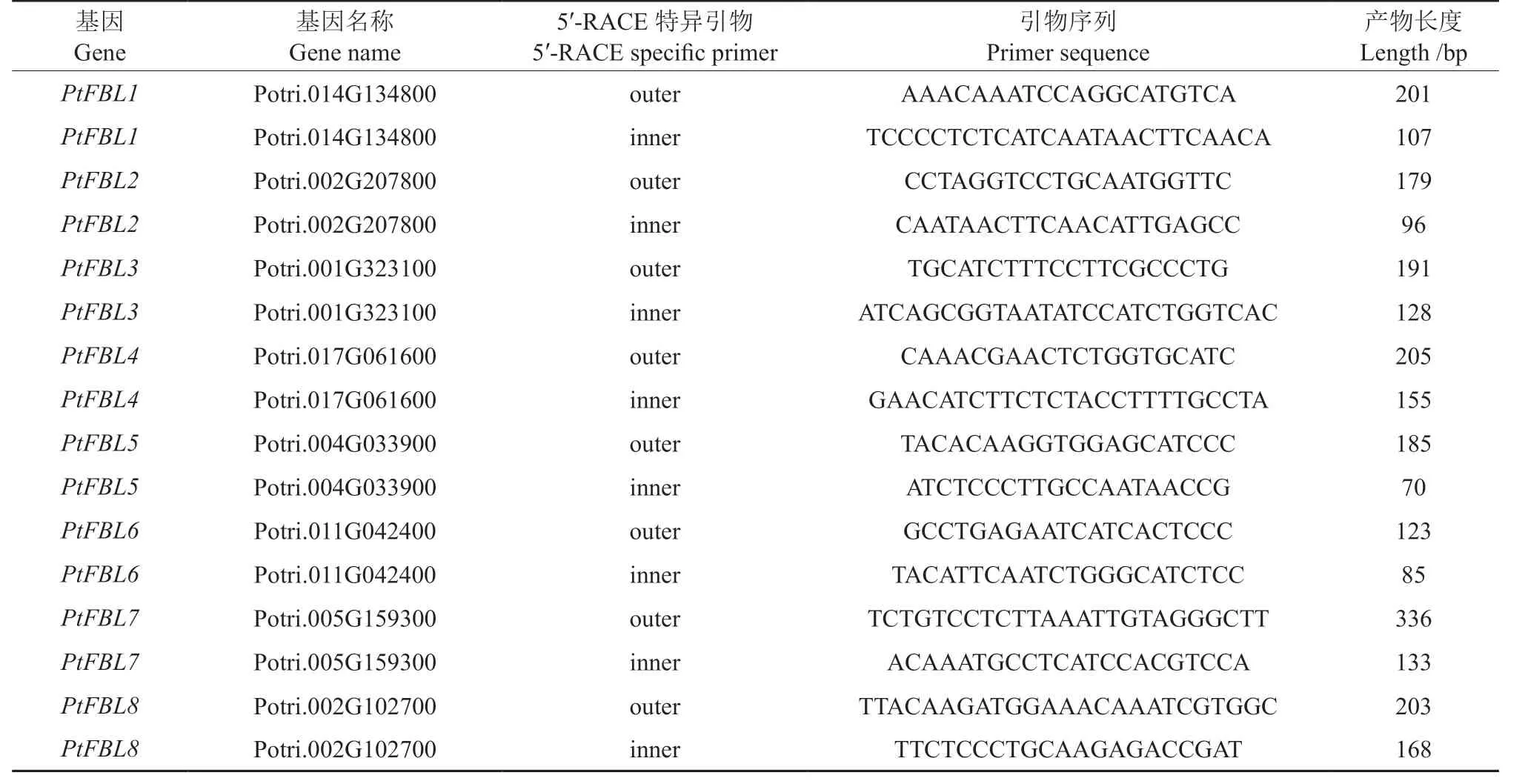
表2 杨树FBL 基因家族5′-RACE 特异引物序列Table 2 5′-RACE specific primer sequences of poplar FBLgene family
2 结果与分析
2.1 FBL 基因家族成员的系统进化关系
为分析杨树FBL家族基因与其他物种同源基因的系统进化关系,本研究分别选取双子叶植物、单子叶植物和蕨类植物代表物种拟南芥、水稻和小立碗藓3 个物种的FBL家族基因成员与杨树进行系统进化分析。如图1 所示,FBL基因家族在拟南芥、杨树、水稻和小立碗藓中分别有7个、8 个、8 个和12 个成员。4 个物种的FBL基因按照蛋白序列主要分为了两个大的亚群,杨树的8 个FBL基因主要聚在了第一大亚群,这个大亚群又分为4 个小亚群。PtFBL3和PtFBL4与拟南芥的ATFBL2和ATFBL3先聚在一起,再和水稻的LOC_Os04g32460 聚在一起,最后和LOC_Os11g31620 聚成一个小亚群。PtFBL1和PtFBL2和AtTIR1先聚类到一起,再与ATFBL1聚类,最后与水稻的LOC_Os05g05800 构成第二个小亚群;PtFBL5 和PtFBL6 与小立碗藓的Pp3c20_17150 和Pp3c23_5870 以 及Pp3c23_11300 和Pp3c24_9800聚类组成第三个小亚群。第四个小亚群是由杨树的PtFBL7和PtFBL8与拟南芥的ATFBL4和ATFBL5两两聚类,再一起与水稻的LOC_Os02g52230 和LOC_Os03g08850 聚类。在第二个大亚群中没有杨树的基因,也可以分为4 个亚群:LOC_Os01g63420、LOC_Os05g37690、LOC_Os03g15880 和ATCOL1 聚成一个小亚群;小立碗藓的Pp3c20_4050、Pp3c23_18960、Pp3c3_8580;Pp3c9_10120、Pp3c15_1200 和Pp3c17_7680, 以及Pp3c18_16630 和Pp3c22_6020 分别单独聚类在一起。从进化树也可以看出,杨树PtFBL1-PtFBL4具有较高的同源性,它们是在一个进化分支上,而PtFBL5、PtFBL6和PtFBL7、PtFBL8分别位于不同的分支且与PtFBL1—PtFBL4的同源关系较远。另外,小立碗藓的FBL同源基因具有相对独立的进化分支。

图1 杨树、拟南芥、水稻和小立碗藓FBL 基因家族的 系统进化树Fig.1 Phylogenetic trees of the FBL gene family in Poplar, Arabidopsis, rice and Physcomitrella
2.2 杨树FBL 基因家族的功能结构分析
利用Gene structure display server(GSDS)对杨树FBL基因家族8 个成员的外显子/内含子结构进行分析,并在结果中搜索潜在的ptc-miR393剪切位点。如图2 所示,除了PtFBL7包含4 个外显子和3 个内含子之外,其余7 个FBL基因均由3 个外显子和2 个内含子构成。其中,PtFBL5与PtFBL6与其他FBL基因相比,具有较长的内含子和3′UTR 区序列。此外,ptc-miR393 在PtFBL1、PtFBL2、PtFBL3与PtFBL4中都具有相似度高的互补配对序列且均位于第3 个外显子上。
利用Conserved domain database(CDD)软件对杨树FBL家族8 个基因的保守结构域进行预测,发现PtFBL1、PtFBL2、PtFBL3、PtFBL4、PtFBL5和PtFBL6都具有关键的F-box 结构域和2 个ANN1 super family 结 构 域, 而PtFBL7和PtFBL8则缺少了F-box 结构域和一个ANN1 super family 结构域(图3)。此外,PtFBL3、PtFBL7和PtFBL8在第二ANN1 super family 结构域下游都多了一个LRR_RI super family 结构域,这个结构域与ANN1 super family 结构域有重叠的部分,而ptc-miR393 的互补配对序列位 于 基 因 的F-box、ANN superfamily1、ANN superfamily2 结构域之后。
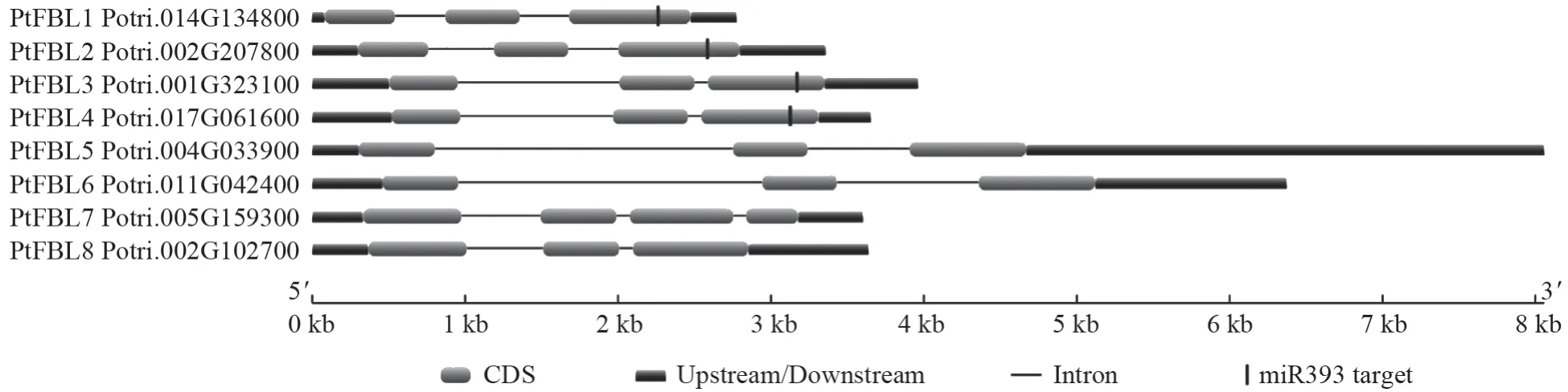
图2 杨树FBL 家族基因的外显子/内含子结构Fig.2 Exon/Intron structure of poplar FBLfamily genes
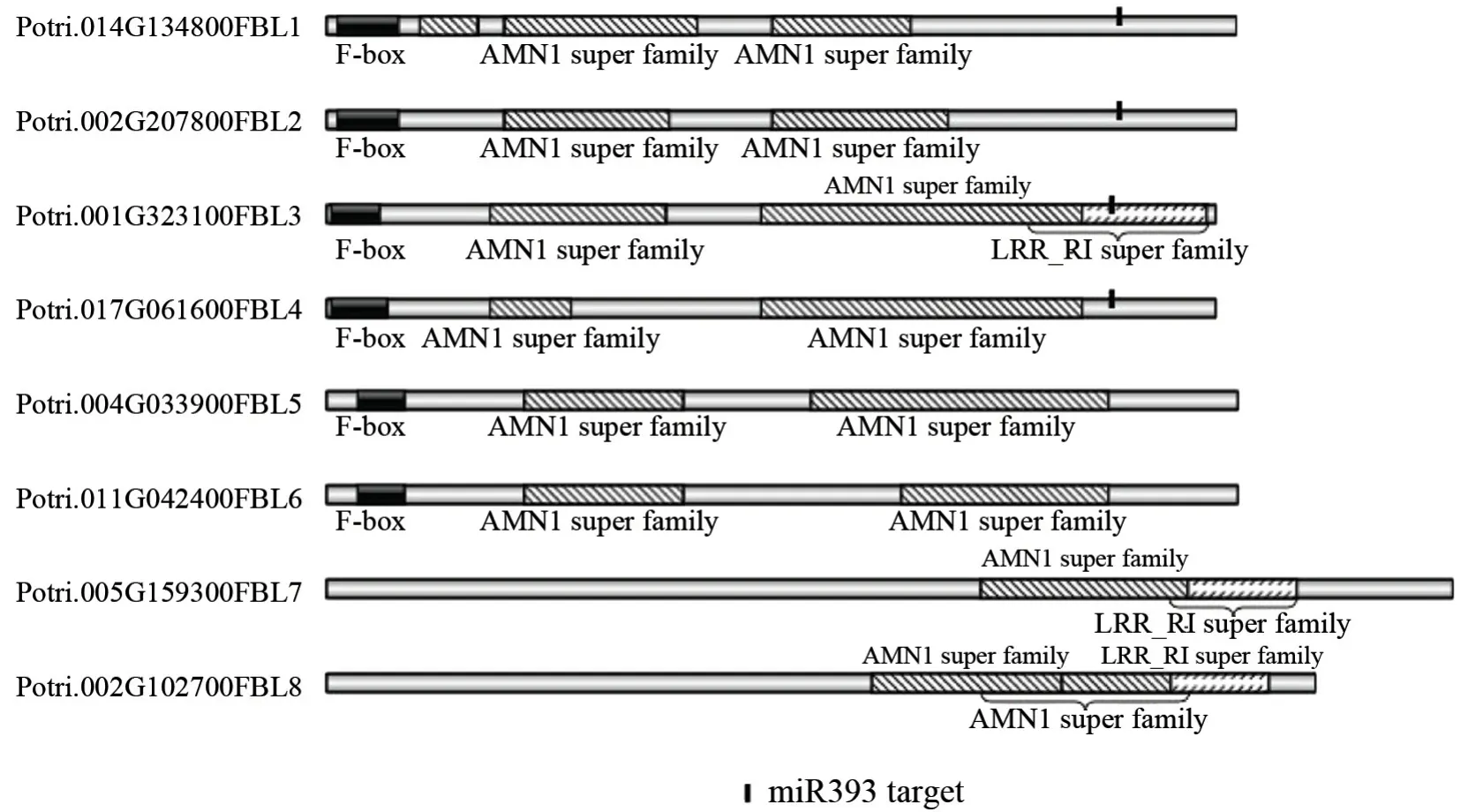
图3 杨树FBL 基因家族成员的结构域分析Fig.3 Domain analysis of poplar FBL gene family members
2.3 杨树FBL 基因家族受miR393 剪切的调控作用
为了检测ptc-miR393 对杨树FBL家族成员的靶向作用,利用ClustalX2 对杨树FBL基因家族8 个成员的CDS 序列和ptc-miR393 进行了序列相似性分析,结果显示,PtFBL1和PtFBL2、PtFBL3和PtFBL4、PtFBL5和PtFBL6及PtFBL7和PtFBL8两两之间的序列相似度分别为91.51%、93.94%、92.55%和90.20%,而这8 个基因之间的序列相似度仅有57.48%,说明杨树FBL基因之间存在一定的序列差异,也和它们的功能结构差异相对应。同时,ptc-miR393 与PtFBL3、PtFBL4的反向互补序列同源相似度最高,其次是与PtFBL1、PtFBL2,与PtFBL5、PtFBL6以及PtFBL7、PtFBL8相似度较差(图4a)。将ptc-miR393 成熟体序列与杨树FBL基因家族成员的核酸序列进行配对比较,发现PtFBL1、PtFBL2、PtFBL3和PtFBL4与ptc-miR393 的互补配对率较高,只有2 ~3 个碱基的差异,并且是在ptc-miR393 的3′末端区域。而PtFBL5、PtFBL6、PtFBL7和PtFBL8与ptcmiR393 有5 ~7 个碱基差异,且在ptc-miR393 的5′端前10 个碱基就有1 ~2 个碱基差(图4b)。根据ptc-miR393 与FBL基因的碱基配对差异,推测PtFBL1-PtFBL4受ptc-miR393 的剪切调控,而PtFBL5-PtFBL8不受ptc-miR393 的剪切。
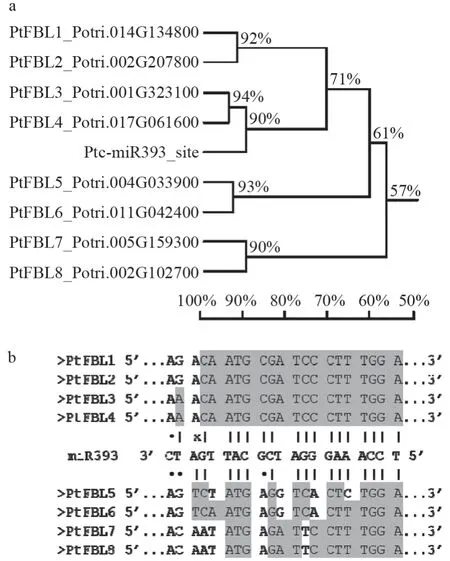
图4 杨树FBL1-8 基因与ptc-miR393 核酸序列的比对Fig.4 Nucleic acid sequence alignment of poplar FBL1-8 gene and ptc-miR393
2.4 杨树miR393 与FBL 家族基因的表达模式
为了进一步检验杨树miR393 对FBL基因家族成员的调控作用,对ptc-miR393 与FBL家族基因在杨树不同发育时期的不同组织中进行表达分析。如图5 所示,PtFBL3和PtFBL4的表达模式一致,与PtFBL2的表达趋势也比较相似,但与PtFBL1的表达模式存在较大差异。PtFBL1在杨树1 个月的组培苗的根、茎、叶中高表达,而PtFBL2、PtFBL3和PtFBL4是在组培苗的叶片和茎中高表达。PtFBL1—PtFBL4在杨树3 个月的土培苗中都表现出相似的表达趋势,特别在幼嫩的组织中高表达,比如在未展开叶片及顶端分生组织;同时随着茎间数的增多,PtFBL1—PtFBL4的表达量逐渐降低。相反,ptc-miR393 在杨树幼嫩的组织中表达量都比较低,特别是杨树组培苗的组织。在3 个月的土培苗中,ptc-miR393 的表达量从顶端分生组织到茎基部逐渐增多,与PtFBL1—PtFBL4呈相反的表达模式。PtFBL7、PtFBL8与PtFBL1—PtFBL4表达模式有所不同,它们在土培苗中不同的茎段和未展开叶中都高表达,在15 d组培苗的叶片和茎中表达量也较高。而PtFBL5和PtFBL6只在土培苗中的不同节间和根中高表达,在组培苗中的表达量都较低。从ptc-miR393 与PtFBL1—PtFBL8基因在杨树不同发育时期和组织中的表达模式可以看出,PtFBL1—PtFBL4的表达与ptc-miR393 呈相反的趋势,而PtFBL5—PtFBL8的表达不受ptc-miR393 的影响,说明PtFBL1—PtFBL4可能受ptc-miR393 直接调控。为了验证这一推测,分别对PtFBL1、PtFBL2、PtFBL3、PtFBL4、PtFBL5、PtFBL6、PtFBL7和PtFBL8进行了5′-RACE 的实验验证,结果表明,PtFBL1、PtFBL2、PtFBL3和PtFBL4经过两轮巢式PCR 产物的克隆测序,都能检测到有明确的ptc-miR393剪切位点(图6),而PtFBL5、PtFBL6、PtFBL7和PtFBL8经过二轮巢式PCR 后都未获得目标长度的片段,说明它们不受ptc-miR393 的剪切调控。
3 结论与讨论
3.1 结 论
miR393 的靶基因是FBL基因家族。杨树FBL基因家族共有8 个不同成员,两两成员之间的同源性较高。PtFBL1—PtFBL4之间具有较高的同源 性, 而PtFBL5、PtFBL6和PtFBL7、PtFBL8分别位于不同的分支且与PtFBL1—PtFBL4的同源关系较远,并且它们的结构域有一定的改变。ptc-miR393 与PtFBL1—PtFBL4的互补配对率较高,与PtFBL5—PtFBL8之间有5 ~7 个碱基的差异,通过组织表达分析和5′-RACE 检测,验证出PtFBL1—PtFBL4是ptc-miR393 的靶基因,而PtFBL5—PtFBL8不受ptc-miR393 的剪切调控。
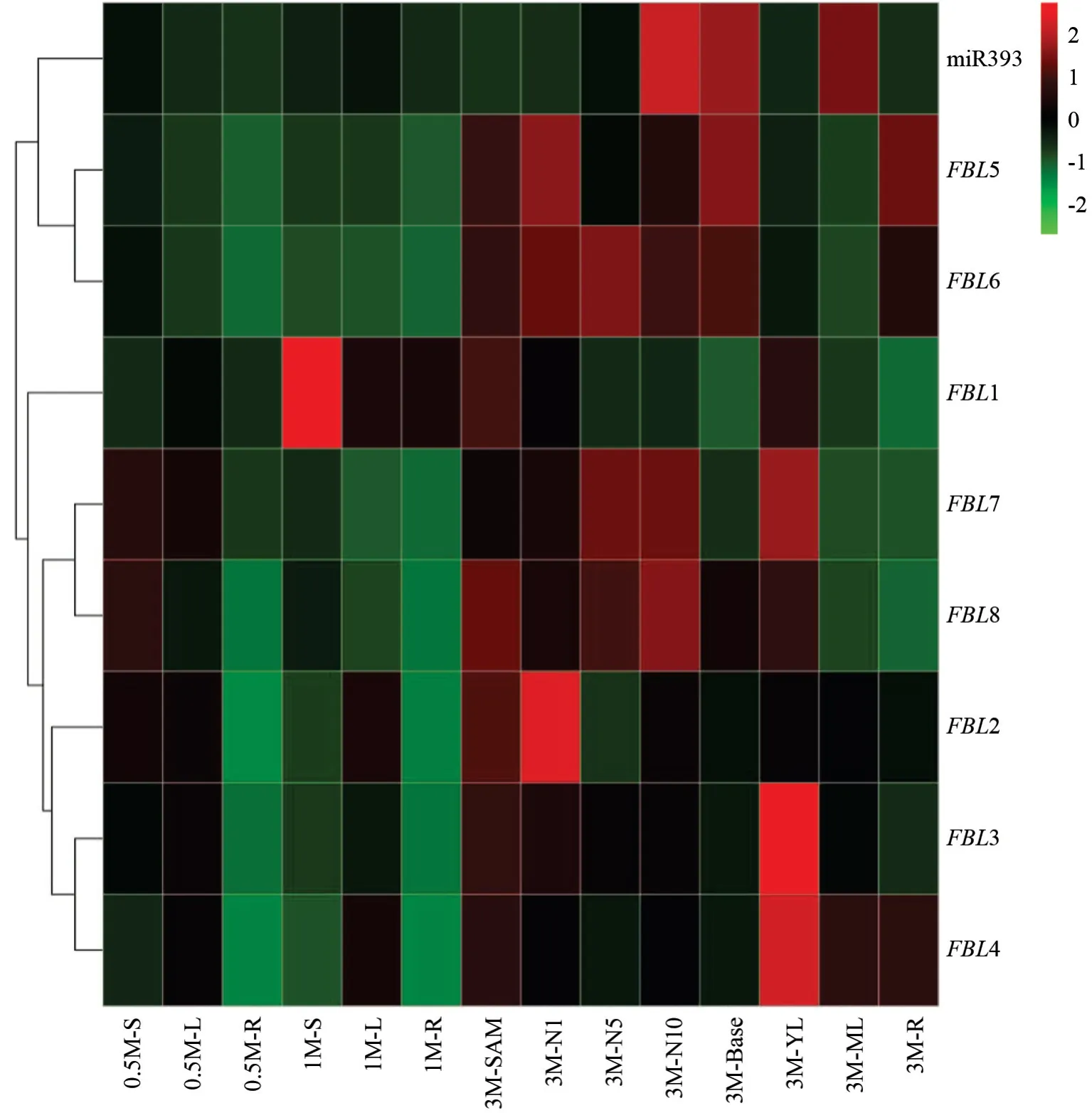
图5 杨树miR393 与FBL 家族基因在不同发育时期组织中的表达Fig.5 The expression of miR393 and FBLfamily genes in poplar tissues at different developmental stages

图6 杨树FBL1-8 基因的5′-RACE 验证结果Fig.6 Results of 5′-RACE validation of FBL1-8 genes
3.2 讨 论
杨 树ptc-miR393 与PtFBL1—PtFBL4的互补配对率较高,而与PtFBL5—PtFBL8之间有5 ~7个碱基差异,并且PtFBL5和PtFBL6在第11 个碱基处有错配。根据植物miRNA 与靶基因的配对原则,miRNA 与靶基因不得有超过4 个碱基的错配,并在5′端的第10 或11 位点不得有碱基错配;miRNA 与靶基因结合的种子序列(miRNA 5′端起的第1 ~12 个碱基)不得有相邻位点及超过2.5 个碱基的错配[25-26]。推测PtFBL1—PtFBL4基因可能受ptc-miR393 的调控,而PtFBL5—PtFBL8则不受其调控。进一步通过ptc-miR393 和FBL基因家族成员的组织表达模式以及PtFBL1-8基因的5′-RACE 检测,验证出PtFBL1、PtFBL2、PtFBL3和PtFBL4受到ptc-miR393 的剪切,可以确定是ptc-miR393 的靶基因。已有研究表明,与杨树PtFBL1—PtFBL4同源的拟南芥AtTIR1、AtAFB1、AtAFB2、AtAFB3基因和水稻OsTIR1与OsAFB2基因都受到miR393 的调控[6,11]。说明miRNA 与靶基因之间的碱基配对率会直接影响miRNA 对靶基因的作用效果。
从FBL基因家族的系统进化树上可以看出,受到miR393 调控的FBL成员都聚类在一个分支上,并且它们在miR393 的结合位点处有相同的序列,推测它们可能在植物发育过程中具有相似的作用和功能。相反,杨树PtFBL5—PtFBL8基因都不受ptc-miR393 的剪切调控。PtFBL7和PtFBL8与拟南芥和水稻的FBL基因聚在一起,而PtFBL5和PtFBL6却单独和小黎碗藓的FBL基因聚类,且表达模式也与杨树其他FBL基因不一样,说明PtFBL5—PtFBL8的生物学功能可能与PtFBL1—PtFBL4不同。 此外,PtFBL1—PtFBL4基因的剪切位点位于FBL基因关键的F-box、ANN superfamily1、ANN superfamily2 结构域之后,说明PtFBL1-PtFBL4受到ptc-miR393 剪切后,其5′端主要的结构域就会被降解,因此ptc-miR393 对PtFBL1—PtFBL4的调控作用较强。
本研究从FBL基因家族的物种进化关系、基因结构分析以及与ptc-miR393 之间调控作用的鉴定等方面进行研究,揭示了ptc-miR393 与杨树FBL基因家族成员间的剪切作用。这为进一步研究ptc-miR393 和FBL基因的生物学功能,以及揭示ptc-miR393 与杨树FBL基因之间的调控功能奠定基础。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
教学考试(高考生物)(2020年6期)2020-11-23 05:25:56
食品与生物技术学报(2020年8期)2020-01-06 08:00:56
科学24小时(2019年5期)2019-06-11 08:39:38
发明与创新(2019年9期)2019-03-26 02:22:48
时代英语·高一(2019年1期)2019-03-13 10:29:48
生命科学研究(2018年1期)2018-05-29 01:12:47
作文评点报·低幼版(2017年24期)2017-06-19 14:59:13
喜剧世界(2017年10期)2017-06-01 12:39:29
上海农业学报(2017年3期)2017-04-10 12:39:14
