
珙桐DiZF-CCCH1基因的克隆及表达分析
2020-06-06 08:18:16纪小敏曹福祥董旭杰
中南林业科技大学学报 2020年5期
纪小敏,李 建,李 萌,曹福祥,董旭杰
(1.中南林业科技大学 生命科学与技术学院,湖南 长沙 410004; 2.湖南农业大学 园艺园林学院,湖南 长沙 410128)
珙桐Davidia involucrata是珙桐科珙桐属高大乔木,我国特有的珍稀孑遗植物,国家一级保护植物[1]。珙桐种子休眠期长,败育率高,严重限制了其自然更新能力。随着分子生物学技术的快速发展,分子标记技术在分析珙桐遗传变异中得到应用,Long 等[2]从珙桐叶片mRNA 转录本中开发基因的简单重复序列(SSRs),共获得了30 411个基因的SSR 位点,12 208 对引物,该研究建立和完善了珙桐表达序列标签数据和新SSR 标记,为揭示珙桐物种的适应性进化、种群结构和解决遗传多样性等问题提供依据[3-4]。珙桐中的关键基因也不断被克隆鉴定,例如季红春等[5]从珙桐cDNA 文库中克隆获得了一个未知基因DiRCI,实验结果表明该基因编码低温诱导膜蛋白。刘美等[6]成功得到了珙桐热休克蛋白18(HSP18)的基因序列,对其进行生物信息学分析,推测该基因可能与珙桐种子对不良环境的适应性相关。戴鹏辉等[7]克隆了一个与花青素相关且编码MYB 转录因子的DiMYB1基因,表达分析发现DiMYB1在败育种子中的表达量均高于正常种子,推测该基因可能在珙桐种子败育过程中发挥重要作用。熊亚丽等[8]克隆了珙桐纤维素合酶CesA基因,表达模式分析发现其在败育种子中上调表达,推测该基因是调控纤维素合成的关键基因并且在珙桐败育过程中起到调控作用。吴晓波等[9]克隆得到了珙桐内果皮发育过程中差异表达基因DiCCoAOMT1, 功能分析显示该基因可能是珙桐内果皮木质素合成的关键基因。Ren 等[10]以珙桐不同器官为材料,定量分析了珙桐不同器官中稳定表达的内参基因,研究结果表明苞片发育中最稳定的内参基因为TIP41、CAC和CSD,正常和败育种子中最稳定的内参基因为SAND、CAC和eIF,不同颜色的叶片中最稳定的内参基因为SAND、18SrRNA和eIF。越来越多的基因被克隆鉴定,为从分子水平上研究珙桐提供了一定的理论支撑。
为研究珙桐种子发育的分子机制,本研究小组前期完成了珙桐转录组测序分析,构建了珙桐unigene 库,从unigene 库中发现大量与生殖发育相关的关键基因。其中筛选到一个编号为c37849.graph_c0的转录本,其在珙桐正常种子中高量表达,而在发育异常的种子中几乎检测不到表达。序列分析显示,该基因是一个编码CCCH 型锌指蛋白的转录因子,同源序列比对结果表明该基因含有植物特有的RR-TZF 结构域。
锌指蛋白(Zinc finger protein)是由一段小分子肽链组成,可与Zn2+结合形成指形结构,从而发挥特定功能的转录因子,最早发现于非洲爪蟾Xenopus laevis的卵母细胞中[11-12],其指形结构能与核酸、蛋白质及一些小分子结合而行使调控功能。根据结构域中半胱氨酸(Cys)及组氨酸(His)的数量与位置,锌指蛋白可分为C2H2、C2HC、C2HC5、CCCH、C3HC4、C4、C4HC3、C6和C89个亚类[13]。其中CCCH 型锌指蛋白在结构上含有一个或多个由3 个半胱氨酸(Cys)和1 个组氨酸(His)按序排列组成的锌指模体,这种结构可以被归纳为:C-X6-14-C-X4-5-C-X3-H (X 代表任意氨基酸残基)[14]。TZF 蛋白(Tandemn zinc finger protein)是CCCH 型锌指蛋白的一个亚家族,具有C-X7-8-C-X5-C-X3-H-X18-C-X5-C-X4-C-X3-H 的结构特征,如人类的hTTP、ZFP36L1 和ZFP36L2 均属于此亚家族[15-16]。根据结构域组成,TZF 蛋白可分为RR-TZF 与ANK-RR-TZF 2 种类型,其中RR-TZF 蛋白是其中最大的亚家族之一,是植物特有的一类TZF 蛋白,其主要特征是在TZF 结构域上游有一段富含精氨酸的50 多个氨基酸的保守区域C-X5-H-X4-C-X3-H motif(RR)[17]。大多数植物RR-TZF 编码的基因在不同组织的不同发育阶段特异表达。例如,拟南芥Arabidopsis thaliana中的AtTZF4、AtTZF5、AtTZF6基因被发现参与种子萌发[18];AtSZF1和AtSZF2可以增加拟南芥盐胁迫抗性[19]。水稻Oryza sativa中的OsLIC基因参与了叶鞘夹角形成[20];OsDOS能延缓叶片衰老[21]。 杨树Populus deltoides中的PdC3H17基因影响次生细胞壁的形成[22]。棉花Gossypium hirsutum中的GhZFP1超量表达,可通过改变钠钾离子平衡以增强拟南芥抗盐胁迫的能力[23]。本研究克隆了珙桐DiZF-CCCH1的cDNA 全长序列,利用生物信息学方法对其编码的氨基酸的理化性质、保守功能域、磷酸化位点、信号肽结构、二级结构组成等进行了分析预测,并且分析了该基因在不同组织器官和种子不同发育阶段的表达模式,以期为珙桐DiZF-CCCH1基因的功能验证和调控机制提供参考依据。
1 材料与方法
1.1 材 料
珙桐小叶、大叶、叶柄、小苞片、大苞片、生长点(顶端分生组织)、种子和花序等均取自湖南省张家界市桑植县八大公山国家级自然保护区。7—11月为种子的采集时间,4—5月为其余组织样品采集时间,每份样品设置3个生物学重复。将新鲜样品采集后迅速置于液氮中速冻,然后放置-70 ℃冰箱中保存。
1.2 珙桐总RNA 的提取及cDNA 的合成
选取约100 mg 样品,加入液氮快速研磨成粉,采用E.Z.N.A.TMPlant RNA 试剂盒提取总RNA,将各样品的RNA 的浓度稀释至合适的范围内后,采用Prime ScriptTM1stStrand cDNA Synthesis Kit 试剂盒反转录合成cDNA,产物于-20 ℃条件下保存备用。
1.3 目标基因的克隆
使 用Primer 5.0对目的基因ORF(Open reading frame,开放阅读框)序列设计特异引物 DiZF-F/DiZF-R(引物序列见表1),分别在引物两端加入BamHI 和SacI 酶切位点及保护位点。以珙桐种子cDNA 为模板进行PCR 全长扩增,将克隆片段与pMD18-T 载体连接的产物转化至大肠杆菌DH5α 感受态中,将经PCR 鉴定的阳性单菌落送至铂尚生物技术有限公司测序。
1.4 目标基因生物信息学分析
使用Expasy-ProtParam tool、SMART 等在线软件对NCBI 中的ORF Finder 工具预测的目标基因编码蛋白的相对分子质量、氨基酸组成、理论pI 值、亲水性/疏水性等理化性质进行初步分析。使用在线工具Netphos 3.1 server 和TMHMM Server 2.0 等对目的蛋白的磷酸位点和跨膜区域等进行预测分析。使用软件MEGA 7.0 对NCBI 数据库中与目标基因编码的氨基酸同源性较高物种的氨基酸序列进行序聚类分析,并采用软件SPOMA 与Swissmodel 对目标基因编码的氨基酸进行二级、三级结构预测分析。
1.5 目标基因表达模式分析
通过qRT-PCR 检测目标基因在珙桐不同组织部位以及种子不同发育阶段中的表达情况,qRTPCR 反应体系使用2×SYBR Green qPCR Master Mix(High ROX)(Biotool,US),引物序列见表1。选择在珙桐各组织中稳定表达的UBQ基因为内参基因[24],每组样品设置3 个生物学重复,通过2-ΔΔCT方法[25]分析数据。
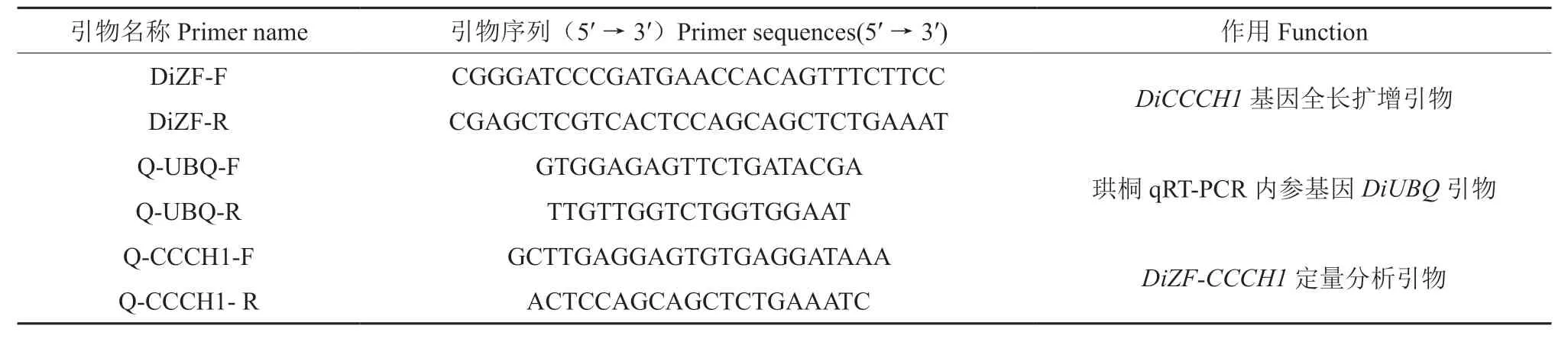
表1 引物序列及扩增信息Table 1 Primer sequences and amplification information
2 结果与分析
2.1 珙桐种子中差异表达基因的筛选
在珙桐转录组数据库中筛选到21 个在正常种子和发育异常种子中差异表达的关键转录本,对它们的表达模式进行分析,发现除了c20843.graph_c1 和c39573.graph_c0 在发育异常种子中上调表达外,其余19 个转录本均下调表达,最明显的是c37849.graph_c0 和c41816.graph_c0,其中编号为c37849.graph_c0 的转录本在正常种子中高量表达,而在发育异常的种子中表达量极低(图1)。通过c37849.graph_c0 在转录组中的功能注释及其序列在NCBI 中的比对结果,预测该基因编码一个与种子发育相关的CCCH 型锌指蛋白,将其选择为本研究的目的基因。
2.2 目标基因的克隆
分别以珙桐gDNA 和cDNA 为模板,通过PCR 扩增得到了2 条长度约为700 bp 的DNA 片段(图2),测序发现gDNA 中扩增获得的片段与从cDNA 中扩增获得的片段序列完全相同。结果证明该基因无内含子,ORF 长度为711 bp,编码236 个氨基酸。通过测序及BLAST 比对,确定该基因编码一个CCCH 型锌指蛋白,将其命名为DiZF-CCCH1。
2.3 生物信息学分析
2.3.1 蛋白质氨基酸序列的预测
通过ProParam 在线软件分析DiZF-CCCH1编码蛋白的理化性质,结果表明,DiZF-CCCH1编码的蛋白分子质量为26.74 kDa,理论等电点(PI)为9.02,包括3 656 个原子,分子式为C1184H1770N352O337S13。蛋白的脂肪指数为47.20,亲水指数为-0.664,不稳定指数为44.24,为不稳定蛋白。在DiZF-CCCH1 蛋白中含有2 个CCCH 锌指结构,C-x7-C-x5-Cx3-H 和C-x5-C-x4-C-x3-H 位于83 ~133 处(图3),由16 个氨基酸隔开,也就是TZF 结构域(C-X7-8-C-X5-C-X3-H-X16-C-X5-C-X4-C-X3-H)。TZF 结构域上游有一段富含精氨酸的50 多个氨基酸保守区域C-x5-H-x4-C-x3-H,位于48 ~63 处,并且含2 个高度保守的结构域SHDWTEC 和ARRRDPR,分 别 位 于53 ~59 和68 ~74 处,因此DiZF-CCCH1 属于植物特有的RR-TZF 蛋白。
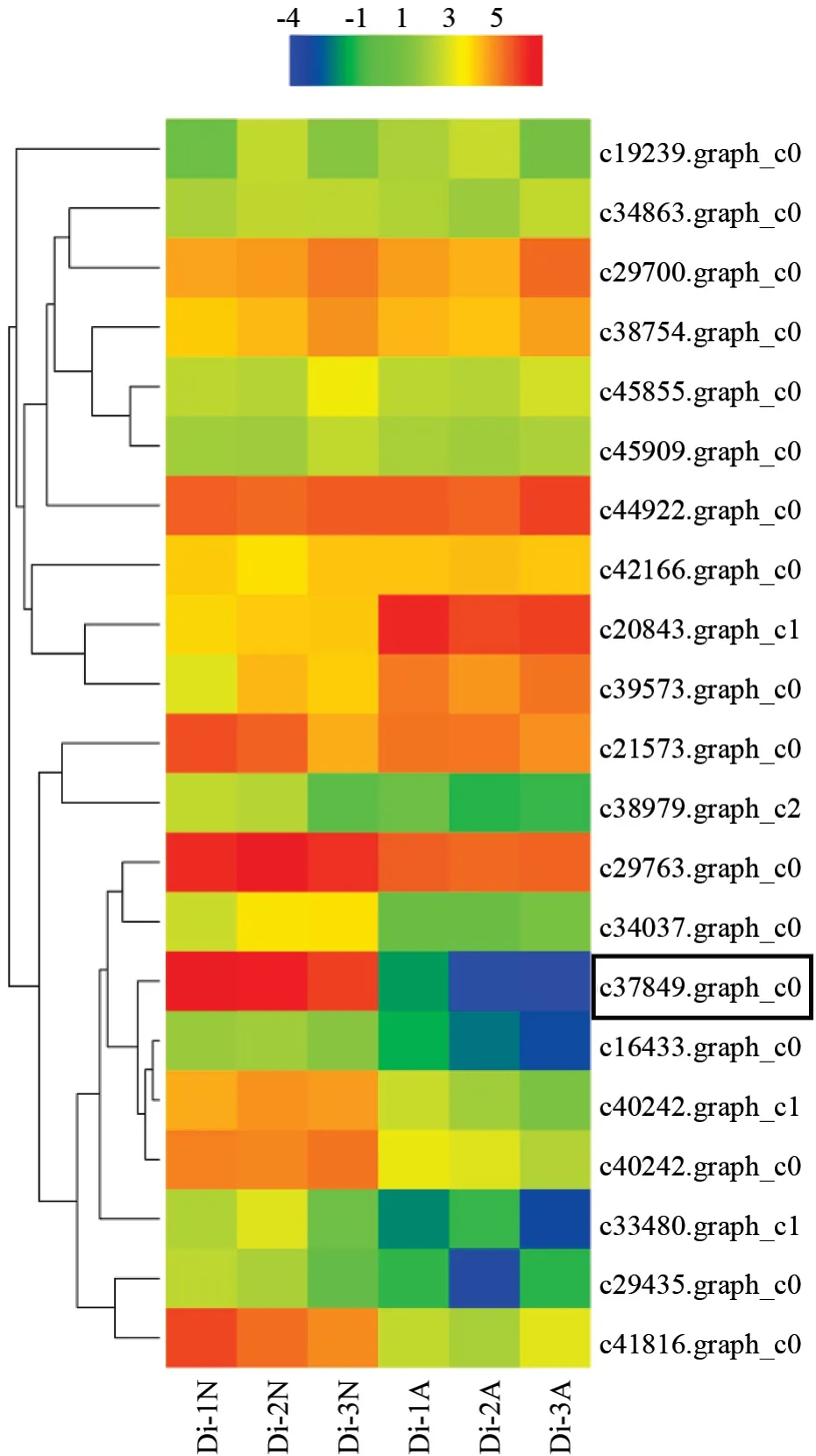
图1 珙桐种子发育相关关键转录本的表达模式Fig.1 Expression patterns of selected transcripts in normal and aborted seeds of Davidia
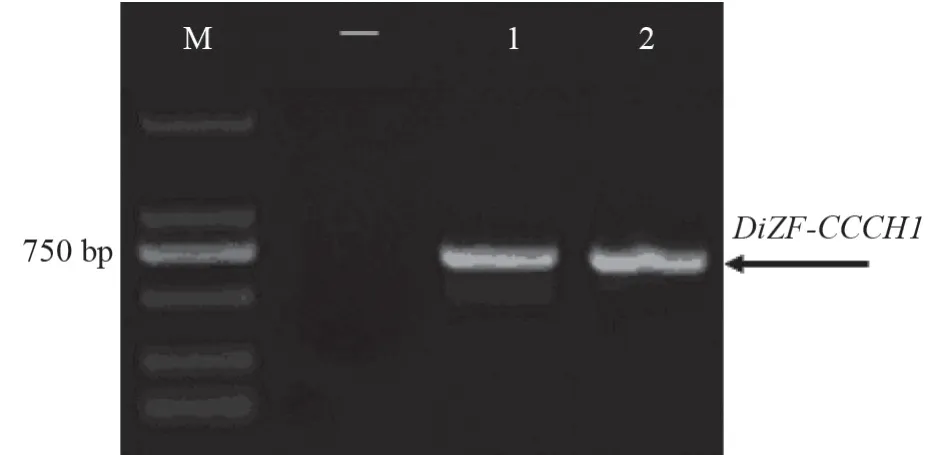
图2 DiZF-CCCH1 基因的克隆Fig.2 Cloning of DiZF-CCCH1gene

图3 推导出的DiZF-CCCH1 蛋白的氨基酸序列Fig.3 Deduced amino acid sequence of DiZF-CCCH1
2.3.2 序列分析及聚类分析和亚细胞定位
利用软件MEGA 7.0 构建系统发育树,对DiZF-CCCH1 蛋白进行序聚类分析,结果显示,DiZF-CCCH1 与来源于甜橙Citrus sinensis的亲缘关系最近,在一个支,与莲Nelumbo nucifera、银白杨Populus alba和毛果杨树Populus trichocarpa亲缘关系较近(图4)。用TargetP 1.1 Server 对DiZF-CCCH 蛋白进行亚细胞定位,结果显示,该蛋白定位于叶绿体的概率为0.134,定位于线粒体的概率为0.010,定位于细胞核或其他细胞器的概率为0.236,作为信号肽的概率为0.502。
2.3.3 磷酸化位点的预测和跨膜区域的预测
通过Netphos 3.1 server(http://www.cbs.dtu.dk/services/NetPhos/)软件预测了珙桐DiZF-CCCH1蛋白的磷酸化位点,结果(图5A)显示,DiZFCCCH1基因编码的氨基酸序列可能发生磷酸化的位点有20 个,丝氨酸(Ser)可能发生磷酸化的位点有11 个,苏氨酸(Thr)可能发生磷酸化的位点有5 个,酪氨酸(Tyr)可能发生磷酸化的位点有4 个。利用TMHMM Server 2.0 在线预测珙桐DiZF-CCCH1基因编码的氨基酸序列的跨膜结构域(图5B),发现其不存在跨膜结构,由此推测DiZF-CCCH1 蛋白不经过跨膜转运。

图4 DiZF-CCCH1 蛋白的系统进化分析Fig.4 Phylogentic analysis of DiZF-CCCH1
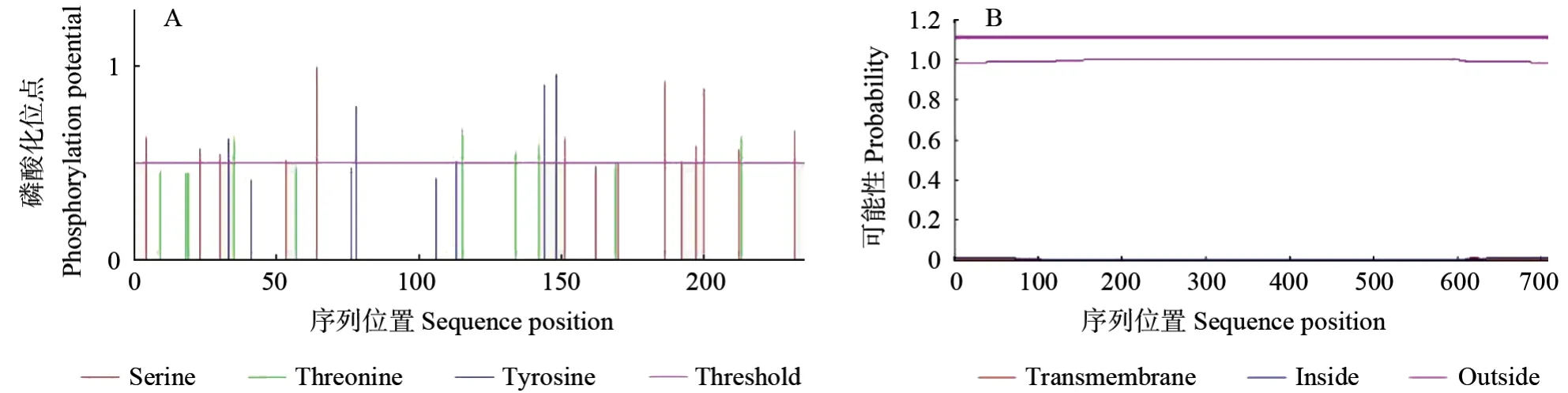
图5 DiZF-CCCH1 磷酸化位点的预测和跨膜区域的预测Fig.5 Transmembrance domain prediction and phosphotylation site prediction of DiZF-CCCH1
2.3.4 二级结构和三级结构的预测
通过SOPMA 对DiZF-CCCH1 的二级结构进行预测,结果如图6A所示。在其二维结构组分中,α-螺旋(H)占13.50%,β-折叠(E)占17.30%,无规则卷曲(C)占68.78%,其他结构占0.42%。无规则卷曲是该蛋白质二级结构中最大的结构元件。运 用SWISS-MODEL(http://swissmodel.expasy.org/)在线软件对DiZF-CCCH1 蛋白的三维结构进行预测,结果如图6B 所示。
2.4 珙桐DiZF-CCCH1 基因表达模式分析
使用qRT-PCR 在珙桐各组织中对DiZFCCCH1基因的表达量进行分析,结果表明,DiZF-CCCH1基因在种子中的表达量显著高于其他组织,在小叶、大叶、生长点中表达量较低,在叶柄、小苞片、大苞片、花序和子房中表达量极低。种子中DiZF-CCCH1基因的表达量约为小叶的6 倍,约为大叶、生长点的15 倍,约为叶柄、小苞片、大苞片、花序和子房中的80 倍(图7)。对珙桐种子不同发育阶段DiZF-CCCH1基因的表达量进行分析发现,DiZF-CCCH1基因在种子发育各个时期中均有表达,其中在发育前期种子中的表达量最高,约为在发育中期和后期种子中的2 倍,暗示该基因可能主要在种子发育前期发挥功能。

图6 DiZF-CCCH1 蛋白的二级结构和三级结构的预测Fig.6 Prediction of secondary structure and three-dimensional structure of DiZF-CCCH1

图7 DiZF-CCCH1 基因在不同组织中的表达水平Fig.7 Expression levels of DiZF-CCCH1gene in different tissues
3 结论与讨论
3.1 结 论
本研究克隆了在珙桐种子中差异表达明显的基因DiZF-CCCH1,序列分析发现该基因ORF 长度为711 bp,编码236 个氨基酸,无内含子。分析DiZF-CCCH1编码蛋白的理化性质,发现DiZFCCCH1编码的蛋白分子质量为26.74 kDa ,理论等电点(PI)为9.02,包括3 656 个原子,分子式为 C1184H1770N352O337S13。蛋白的脂肪指数为47.20,亲水指数为-0.664,不稳定指数为44.24,为不稳定蛋白,属于植物特有的RR-TZF 蛋白。DiZFCCCH1 蛋白的系统进化分析结果表明,DiZFCCCH1 与来源于甜橙Citrus sinensis的亲缘关系最近。DiZF-CCCH1基因编码的氨基酸序列可能发生磷酸化的位点有20 个且不存在跨膜结构,无规则卷曲是DiZF-CCCH1 二级结构中最大的结构元件。qRT-PCR 结果表明,DiZF-CCCH1基因在种子中的表达量显著高于其他组织,在其他组织中仅有微量表达,在种子发育前期的表达量最高约为发育中期和后期的2 倍,暗示该基因可能主要在种子发育前期发挥功能。
3.2 讨 论
3.2.1 珙桐DiZF-CCCH1 结构域分析
本研究从珙桐中克隆到一个编码CCCH 型锌指蛋白转录因子的基因DiZF-CCCH1,对该基因的核苷酸序列及其编码产物进行生物信息学分析,发现DiZF-CCCH1基因编码的蛋白含有2 个C-X6-14- C-X4-5-C-X3-H 结构域,并且在这2 个CCCH 结构域的上游还含有一段富含精氨酸的50 多个氨基酸保守区C-X5-H-X4-C-X3-H,因此认为该蛋白属于RR-TZF 型锌指蛋白。RR-TZF 是植物特有的一类TZF 蛋白,其在玉米、水稻、拟南芥、杨树等基因组分布广泛,玉米基因组中有68 个编码CCCH 锌指结构蛋白的基因,分为7 个亚家族,其中有12 个基因编码RR-TZF 蛋白[26];水稻中有67 个编码TZF 蛋白的基因,分为8 个亚家族,其中有9 个基因编码RR-TZF 蛋白[14];拟南芥中有68 个编码TZF 蛋白的基因,分为11 个亚家族,其中有11 个基因编码 RR-TZF 蛋白[14];杨树含有91 个编码TZF 蛋白的基因,分为13 个亚家族,其中有16 个结构保守的RR-TZF 蛋白[27]。目前关于珙桐CCCH1基因及其功能还未见报道,因此对DiZFCCCH1功能的研究有利于丰富对CCCH 型锌指蛋白功能的认识。
3.2.2DiZF-CCCH1 在珙桐种子中的特异性表达
有研究发现,CCCH 型锌指蛋白可参与植物种子发育,例如:拟南芥AtTZF4、AtTZF5、AtTZF6在种子中特异表达,并且参与种子的萌发,对种子的萌发调节至关重要[18];拟南芥的PEI1是胚胎特异性转录因子,主要在胚胎顶端区域发挥作用,参与胚的发育[28]。本研究分析DiZF-CCCH1表达模式,发现其在种子发育前期的表达量最高,后期表达量降低,在其他组织中仅有微量表达,并且该基因是珙桐种子中差异表达显著的一个基因,意味着该基因可能在种子发育中具有一定的功能。本研究对珙桐DiZF-CCCH1基因的克隆与表达分析,为研究该基因在珙桐种子发育过程中的功能鉴定奠定了基础,对珙桐DiZF-CCCH1基因进行的生物信息学分析,为后续验证DiZF-CCCH1基因的功能提供了参考,但该基因的功能及调控机制尚不明确。下一步将通过载体构建和遗传转化拟南芥,以期明确DiZF-CCCH1的具体功能。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28 07:46:10
实用临床医药杂志(2021年13期)2021-01-10 19:52:18
花卉(2020年16期)2020-08-22 02:34:50
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
现代园艺(2018年1期)2018-03-15 07:56:06
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
中国新技术新产品(2015年22期)2015-12-21 14:02:41
中华胰腺病杂志(2015年5期)2015-12-08 12:18:10
现代检验医学杂志(2015年2期)2015-02-06 02:01:01
