
添加Fe2+对克雷伯氏菌发酵产氢特性的影响
2020-06-04 03:11:34丁鹏飞李艳宾蒋卉张琴
应用化工 2020年4期
丁鹏飞,李艳宾,蒋卉,张琴
(1.安徽工程大学 生物与化学工程学院,安徽 芜湖 241000;2.塔里木大学 生命科学学院,新疆 阿拉尔 843300)
近年来,采用木质纤维素开发清洁能源已受到国内外广泛关注[1-2]。氢能的开发已成为各国竞争的焦点[3-4]。木质纤维素水解液发酵产氢已有一定研究[5-6],然其产氢量仍有待提高。氢酶在调控细菌产氢中起重要的催化作用[7-8],其活性中心可与Fe、Ni金属元素结合[9]。Fe可作为氢酶的激活剂[10],还是氢酶电子载体——铁氧还蛋白活性中心的组成成分[11],可通过影响氢酶的活性改善微生物产氢性能[12],从而提高生物氢产量[13]。
本研究通过检测不同浓度Fe2+对克雷伯氏菌发酵棉秆水解糖液产氢过程的影响,阐明Fe2+添加对菌株发酵产氢的调控作用,对棉秆水解糖液产氢的高效调控具有重要指导意义。
1 实验部分
1.1 菌种和棉秆原料
发酵菌种克雷伯氏菌(Klebsiellasp.WL1316)为一株分离自塔里木河流域野生鲤鱼肠道的产氢细菌[14],其活化、种子、发酵相关的培养基参考文献[14-15],发酵培养基中通过添加FeSO4提供Fe2+来源,将Fe2+质量浓度换算为0,10,20,30,40,50 mg/L添加至发酵培养基中,每个重复处理3次;棉秆原料采自新疆阿拉尔棉田。
1.2 棉秆水解糖液的制备
棉秆水解糖液制备方法参照Zhang等报道的棉秆酸解优化工艺[16]进行,获得的糖液经脱毒、脱色后再经旋转蒸发仪浓缩至所需糖浓度备用。
1.3 菌种的活化、接种与发酵产氢
参考文献[15]的方法,菌种活化并制备种子液,进一步接种发酵培养基,于生化培养箱37 ℃培养。每24 h收集氢气并取发酵液,检测菌体OD600,并测定上清液中的葡萄糖和木糖浓度。发酵周期为120 h,在各发酵时间点将相应发酵时间段的产氢量累加换算为累积产氢量∑Vt。
1.4 分析检测和计算方法
葡萄糖或木糖浓度分别采用DNS[17]、地衣酚比色法[18]测定。葡萄糖(或木糖)利用率表示为发酵过程中利用的葡萄糖(或木糖)浓度占初始葡萄糖(或木糖)浓度的百分比。基于还原糖利用的产氢效率Y(H2/S)表示为H2产生终浓度与利用的还原糖浓度的比值,基于还原糖利用的菌体生长效率Y(OD600/S)表示为菌体生长的OD600变化值与利用的还原糖浓度的比值。
不同Fe2+添加浓度下的累积产氢量采用改进的Gompertz方程进行动力学拟合[19],方程如下:
其中,H代表累积产氢量(mL/L),P代表产氢潜力(mL/L),Rm代表最大产氢速率[mL/(L·h)],λ代表延滞期时间(h),e=2.718 28;采用Sigmaplot 12.0软件对动力学参数P、Rm和λ值进行计算和分析,并获得相关系数R2。采用Origin 8.0软件对各处理数据进行计算、分析及作图。
2 结果与讨论
2.1 Fe2+添加对克雷伯氏菌发酵棉秆水解糖液产氢过程的影响
本研究采用的Klebsiellasp.WL1316是一株具有Ni-Fe氢酶的产氢细菌[20],因此,在发酵产氢体系中添加一定浓度Fe2+可能提高氢酶活性,从而促使菌株合成生物氢。对该菌株发酵120 h内的产氢量进行了监测,换算为累积产氢量,结果见图1。

图1 不同Fe2+添加浓度下的累积产氢量随时间的变化
由图1可知,在较低Fe2+浓度添加范围内(10~20 mg/L),菌株发酵120 h 的累积产氢量高于对照处理(0 mg/L)的,尤以Fe2+添加浓度为20 mg/L的累积产氢量最高,达2 200 mL/L以上,较之0 mg/L处理的提高约20%,可见,添加该浓度Fe2+的处理能有效促进菌株合成生物氢。当Fe2+添加浓度提高至30 mg/L及以上,累积产氢量则降低,低于0 mg/L处理的,并且Fe2+添加浓度越高,累积产氢量越低。由此说明,当Fe2+添加浓度高于一定值时,则可能抑制菌株生物氢的合成。本研究获得的Fe2+添加对棉秆水解糖液暗发酵产氢的作用规律与一些文献报道中采用葡萄糖作为发酵基质产氢的作用规律类似[21-23],即在一定添加浓度范围内,适当浓度的Fe2+能促进生物氢的合成,由此看来,Fe2+添加至不同的发酵产氢体系对生物氢合成的作用效果可能一致。
表1为Gompertz方程拟合结果。

表1 不同Fe2+添加浓度下Gompertz方程拟合累积产氢量的参数和相关系数Table 1 The Gompertz equation fitted parameters and correlation coefficients of cumulative hydrogen production at different Fe2+ addition concentrations
由表1可知,模型的相关系数R2均达0.98以上,表明Gompertz方程能够较好地对各Fe2+浓度处理的累积产氢量进行拟合。Fe2+添加浓度为10 mg/L和20 mg/L处理的动力学参数P、Rm值均高于0 mg/L处理的,说明添加10~20 mg/L的Fe2+能较大程度地提高菌株的产氢潜力。从延滞期λ值来看,10 mg/L处理的λ值略低于0 mg/L处理的,表明低浓度Fe2+可能使延滞期缩短,从而促进菌株产氢。随着添加浓度的增加,延滞期有可能延长以增强自身对增加的Fe2+的适应能力,当然,λ值适当地延长,菌株仍可保持较高的产氢潜力,如Fe2+添加浓度为20 mg/L的处理,其λ值较之0 mg/L处理的高,但仍获得了最高的P、Rm值,分别达2 550.63 mL/L,25.08 mL/(L·h)。这些产氢动力学表现与Taherdanak等的报道[23-24]一致。
2.2 Fe2+添加对棉秆水解糖液中还原糖利用和菌体生长的影响
以棉秆水解糖液的还原糖组分主要为葡萄糖和木糖[14,16],因此,本研究对发酵过程的葡萄糖和木糖浓度进行监测,同时动态监测克雷伯氏菌的生长OD600值,并换算为葡萄糖、木糖利用率和OD600增长值ΔOD600,结果见图2。
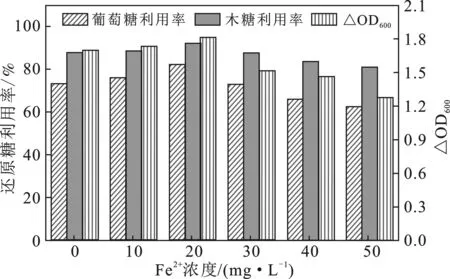
图2 不同Fe2+添加浓度下的葡萄糖、木糖利用率和ΔOD600值
由图2可知,当Fe2+添加浓度为10,20 mg/L时,葡萄糖、木糖利用率和ΔOD600值均高于浓度0 mg/L处理的,尤以20 mg/L处理获得的葡萄糖、木糖利用率和ΔOD600值最高,较之0 mg/L处理的分别提高12%,4.3%和6.5%。可见,该添加浓度有利于发酵基质中还原糖的利用及菌体的生长。随着Fe2+添加浓度的进一步提高,葡萄糖、木糖利用率和ΔOD600值低于浓度0 mg/L处理的,且浓度越高其值越低。由于棉秆水解糖液是主要由葡萄糖和木糖组成的混合糖液,其为克雷伯氏菌利用的规律与一些以葡萄糖为简单碳源的利用规律不同:以葡萄糖为碳源的基质利用率在添加不同浓度Fe2+处理中的差异较小,差异值在5%以内[23-25]。
2.3 Fe2+添加对克雷伯氏菌基于还原糖利用的产氢效率和菌体生长效率的影响
对不同Fe2+添加浓度下的基于还原糖利用的产氢效率Y(H2/S)和菌体生长效率Y(OD600/S)进行了换算,结果见图3。
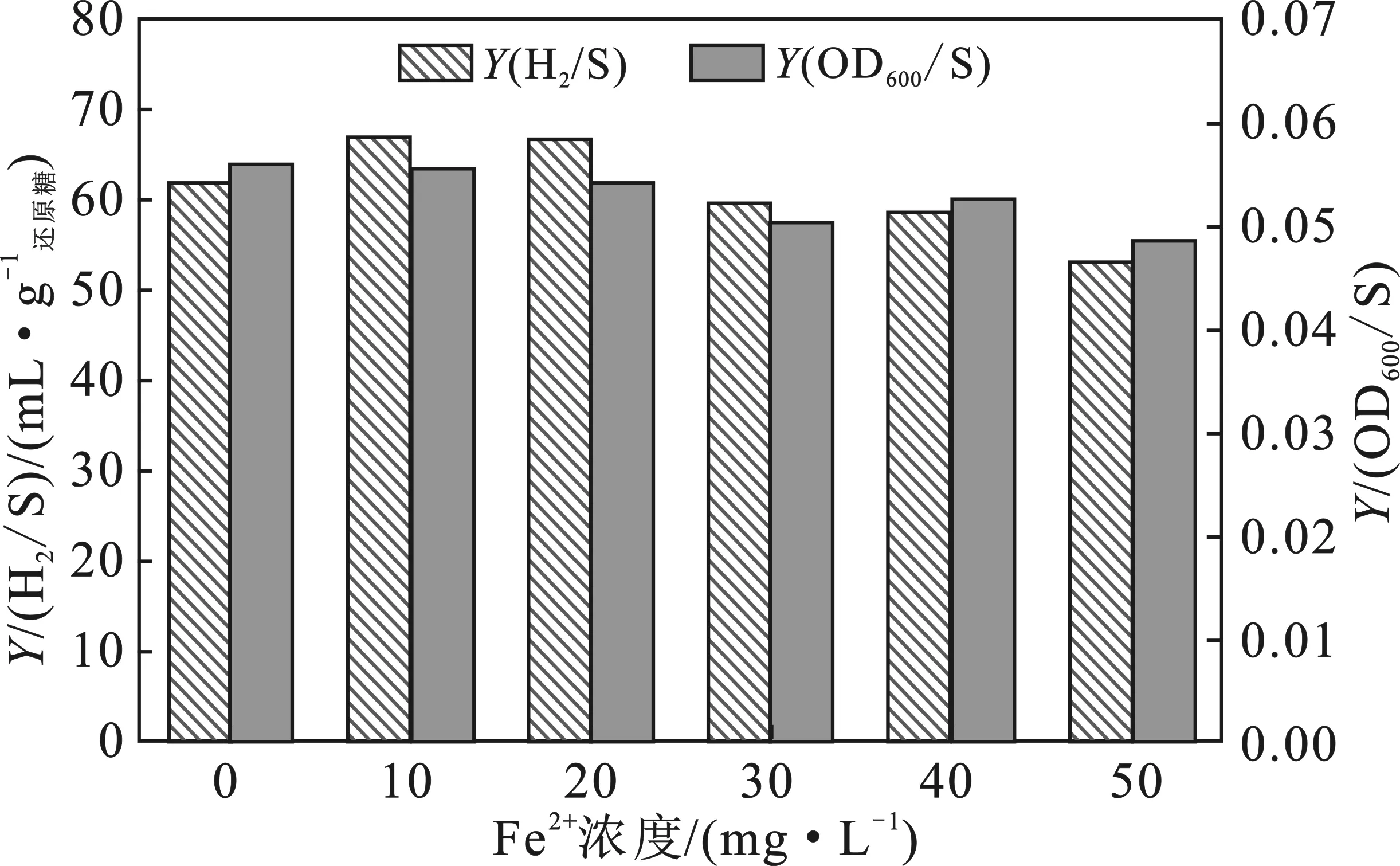
图3 不同Fe2+添加浓度下的基于还原糖利用的产氢效率和菌体生长效率
由图3可知,Fe2+添加浓度为10,20 mg/L处理的Y(H2/S)明显高于0 mg/L处理的,其值分别提高8.3%,7.9%。30,40 mg/L处理的Y(H2/S)与0 mg/L处理的较接近,而50 mg/L处理的明显降低。由此可认为,较低Fe2+添加浓度(10~20 mg/L)在促使克雷伯氏菌保持较高产氢量和还原糖利用率前提下,仍可使其有较高的从还原糖基质合成氢气的转化效率,彰显出该浓度范围较有效地促进产氢的作用。从菌体生长效率Y(OD600/S)来看,其值在低Fe2+添加浓度(10~20 mg/L)范围与0 mg/L处理的接近,而在相对较高浓度处理下则有所降低。可见,尽管克雷伯氏菌在低Fe2+添加浓度范围有较高的ΔOD600值和还原糖利用率,其由还原糖基质用于菌体生长的转化效率实则不够,但其值与0 mg/L处理的接近亦可说明添加Fe2+后,产氢体系仍可保持一定的稳定性。
3 结论
较低Fe2+添加浓度(10~20 mg/L)可促进克雷伯氏菌发酵棉秆水解糖液合成生物氢,尤以Fe2+添加浓度为20 mg/L的累积产氢量最高,达2 200 mL/L以上,较之未添加Fe2+处理的提高约20%,而相对较高浓度Fe2+添加则造成累积产氢量降低,对菌株生物氢的合成产生抑制作用。基于Gompertz模型对添加不同Fe2+浓度下的累积产氢量进行的动力学拟合验证了Fe2+添加对菌株产氢的作用效果,仍以添加20 mg/L Fe2+能最大程度地提高菌株的产氢能力,该浓度处理下的P、Rm值分别可达2 550.63 mL/L,25.08 mL/(L·h)。
Fe2+添加浓度为20 mg/L的处理,菌株获得的葡萄糖、木糖利用率和ΔOD600值最高,较之未添加Fe2+处理的分别提高12%,4.3%和6.5%,表明该添加浓度不仅能有效促进菌株生物氢的合成,亦有利于发酵基质中还原糖的利用和菌体的生长。不同浓度Fe2+添加处理的葡萄糖和木糖利用率差异较大,这与一些以葡萄糖为简单碳源的利用规律不同,彰显出以棉秆水解糖液这种混合糖液为产氢细菌利用中的独特性。在此基础上进行的基于还原糖利用的产氢效率Y(H2/S)证实较低Fe2+添加浓度(10~20 mg/L)在促使克雷伯氏菌保持较高产氢量和还原糖利用率前提下,仍可使其有较高的从还原糖基质合成氢气的转化效率,而基于还原糖利用的菌体生长效率Y(OD600/S)在低Fe2+添加浓度(10~20 mg/L)范围与未添加Fe2+处理的接近,表明其值虽无明显提高,但仍可使产氢体系保持一定的稳定性。
猜你喜欢
物理化学学报(2024年11期)2024-11-06 00:00:00
农业机械学报(2023年5期)2023-05-28 02:28:06
中国农机化学报(2021年7期)2021-08-13 09:47:24
农机化研究(2020年4期)2020-10-19 07:02:12
无机盐工业(2017年5期)2017-05-25 00:37:34
化工管理(2017年25期)2017-03-05 23:32:36
食品界(2016年10期)2016-09-10 01:26:57
山东农机化(2014年4期)2014-04-04 20:07:53
食品工业科技(2014年15期)2014-03-11 18:17:36
影像科学与光化学(2014年5期)2014-03-11 16:03:22
