
松果菊苷对db/db糖尿病小鼠心肌细胞脂质代谢紊乱的改善作用
2020-06-03 09:54郝亚荣
医学研究杂志 2020年4期
张 祥 杨 洛 廖 敏 郝亚荣
糖尿病心肌病(diabetic cardiomyopathy,DCM)是糖尿病的主要心脏并发症之一,其发生率高、危险性高,与糖尿病患者心血管疾病的高发生率、高病死率密切相关[1]。大量研究发现,有5年糖尿病病史的患者DCM发生率在44.4%以上,8年病史时则增加到56%[2, 3]。
DCM是由广泛的心肌血管疾病和糖尿病引起的心肌代谢紊乱引起的心肌坏死性疾病。由于DCM发病机制复杂,通常在发病后期才发现,目前尚无有效的治疗方法[4]。然而,高血糖是DCM发病机制的启动因素。研究发现,在长期的高血糖状态下,糖尿病患者心肌细胞葡萄糖和脂质代谢紊乱,导致心肌细胞能量供应失衡,血管内皮细胞和其他心肌组织损伤,最后导致心肌细胞肥大、坏死、凋亡和心肌间质纤维化等病理变化[5, 6]。因此,糖脂代谢紊乱是DCM病理机制中的关键环节,有效控制和改善心肌细胞能量代谢是糖尿病心肌病的重要治疗策略。
松果菊苷(echinoside)是从管花肉苁蓉茎中分离出来的天然化合物,其主要化学成分是苯乙醇苷[7]。通过体内外研究其已被证明具有抗氧化、抗疲劳、抗炎、提高记忆力与神经保护、抗肿瘤、保护肝脏及免疫调节等作用[8]。笔者以往的研究已经证实,ECH可以调节TGF-β及其下游基因的表达水平,改善db/db小鼠糖尿病肾病的肾纤维化进程[9]。因此,为了进一步揭示ECH对糖尿病心肌病心肌细胞损伤是否具有保护作用,并探讨其可能的分子机制。本研究使用db/db小鼠作为DCM疾病模型,探究心肌细胞脂质代谢紊乱关键调控蛋白PPAR-α及其下游信号通路中重要蛋白CPT -Ⅰ是否参与松果菊苷改善心肌细胞脂质代谢的调控,以期为糖尿病心肌病的治疗提供理论依据。
材料与方法
1.实验动物:本研究采用从南京大学南京生物医学研究所(中国南京)购买的成年雄性C57BLKS/J db/db小鼠和db/m小鼠(SPF级),实验动物许可证号为SCXK(苏)2015-0001。购买后,动物分笼饲养于武汉大学人民医院实验动物中心标准饲养间,室温20℃,相对湿度50%~60%, 24h光照昼夜循环,适应性喂养1周,不给予任何降糖药物。实验程序经武汉大学人民医院实验动物伦理学委员会批准,隶属于武汉大学人民医院动物饲养中心。
2.化学药品和试剂:ECH原粉(批号150623)由上海融禾医药科技发展有限公司提供。本品为白色结晶粉末,采用高效液相色谱法(HPLC)测定其纯度为94%。以乙醇和水为溶剂,在动物实验中进行灌胃给药。2%戊巴比妥钠(上海嘉禾公司,批号10023418);4%多聚甲醛(武汉谷歌生物技术公司,批号AS1075);脱脂奶粉(中国伊利股份有限公司);RT-PCR反转录试剂盒;荧光定量PCR试剂盒;苏丹红Ⅱ染色液(上海歌凡生物技术有限公司)。
3.主要仪器:全自动生化分析仪(ADVID2400,日本东京);全自动显微镜BX63(日本Olympus公司);基因扩增仪(美国ABI公司);实时荧光定量PCR检测仪(7500序列检测系统)(美国ABI公司);DYY7C型稳压稳流电泳仪(北京六一仪器厂);成像系统(Q-IMAGING公司);血糖仪和试纸(美国新泽西州新布伦瑞克强生公司)。
4.动物分组及干预:动物隔离后,采用数字表法将db/db小鼠随机分为糖尿病模型组(db/db组,n=10)和ECH治疗组(db/db+ECH组,n=11),db/m小鼠设为正常对照组(db/m组,n=9)。8周龄时,db/db组和db/m组小鼠按0.05ml/10g体质量标准予以0.9%NaCl溶液灌胃10周,db/db+ECH组小鼠按等体质量予以ECH 300mg/(kg·d) 灌胃10周。每周监测3组小鼠的体质量,观察小鼠的摄食、饮水和活动情况。禁食8h后,每2周采取剪鼠尾法用血糖仪和试纸从鼠尾末端取血样,测定空腹血糖水平。连续干预10周后,用2%戊巴比妥钠腹腔注射(0.1ml/10g)麻醉小鼠。将毛细针插入眼眶后采集血液,快速分离血清后,放入-80℃冰箱保存待测。心脏灌注后,将心肌组织分离称重,检测心脏湿重。取少量左心室心肌组织采用4%多聚甲醛固定后,用于心肌组织切片染色观察病理形态。其余心脏组织置于液氮中1h,-80℃冰箱保存,用于聚合酶链反应(RT-PCR)和Western blot法检测。
5.血清生化指标检测:18周龄时,将采集血样在4℃离心20min(1200r/min)后,收集上层血清。采用全自动生化分析仪检测血清总胆固醇(total cholesterol,TC)、甘油三酯(triglyceride,TG)、游离脂肪酸(triglyceride,FFA)、高密度脂蛋白胆固醇(high density lipoprotein-cholesterol,HDL-C)、低密度脂蛋白胆固醇(low density lipoprotein-cholesterol,LDL-C)。
6.组织病理学观察(HE染色和苏丹红Ⅱ染色):心脏组织切片,各组大鼠取5μm大小心脏组织,用HE染色制片,放置在光学显微镜下观察心肌组织病理改变及心肌细胞凋亡程度。用苏丹红Ⅱ染色法观察心肌细胞的脂质积累。将左心室组织用10%多聚甲醛固定30min,用苏丹红Ⅱ染色液稀释按比例3∶2稀释保存液和水。将保存液置于室温下10min,75%乙醇脱色,PBS冲洗15min。最后甘油明胶封片后,置于显微镜下观察。
7.Western blot法检测组织中PPAR-α、M-CPT-1、CD36和GLUT-4蛋白质表达水平:取各组小鼠心脏组织(50mg),用组织剪刀剪成小块。加入蛋白裂解液,搅拌均匀。加入适量终浓度为1mmol/L的PMSF抑制组织蛋白降解,冰浴裂解20min;1200r/min离心10min,吸取上清液,用BCA法测定蛋白浓度。蛋白样品与SDS-PAGE缓冲液(2×) 1∶1配平后,沸水浴变性10min。取50μg蛋白样品进行SD-PAGE电泳,湿转至PVDF膜上。5% 脱脂牛奶摇床封闭1h, 4℃下过夜。加入一级稀释(1∶800)。洗膜后,分别加入兔抗兔二抗稀释液(1∶5000),室温孵育1h,洗膜后进行扫膜显色,并分析各组条带蛋白灰度值。
8.RT-PCR检测组织中PPAR-α、M-CPT-1、CD36和GLUT-4 mRNA水平:取各组小鼠心脏组织100mg, 按照Trizol试剂盒说明书提取总RNA,紫外分光光度计检测纯度和浓度,电泳检测完整性。按照反转录试剂盒的说明进行反转录。预变性95℃30s,扩增40个循环:PCR反应95℃5s, 60℃40s;溶解95℃15s, 60℃60s,95℃15s。选取管家基因GAPDH作为内参照,使用ABI7500软件2-ΔΔct法处理数据,引物序列详见表1。
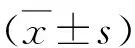
结 果
1.ECH对db/db小鼠一般情况的影响:通过实验期间的持续观察,发现db/db小鼠表现出明显的多食、多食、多尿和肥胖症状,且活动度降低,体型明显大于db/m组小鼠。本实验每2周检测各组小鼠空腹血糖。与db/m组小鼠比较, db/db组小鼠的空腹血糖从8周龄开始持续显著增长,表现出明显的高血糖症状(P<0.01,图1)。在接下来的10周,db/db组小鼠和db/db+ECH组小鼠空腹血糖持续升高,而db/m组血糖水平保持稳定水平。相较于db/db组小鼠,db/db+ECH组小鼠空腹血糖水平显著下降(P<0.01)。db/db组心脏湿重较db/m组明显减轻(P<0.01,表2);而与db/db组小鼠比较,db/db+ECH组小鼠心脏湿重显著增加(P<0.01)。相较于db/m组小鼠,db/db组小鼠体质量明显增加(P<0.01);而经过10周的ECH干预治疗后,db/db小鼠体质量显著降低(P<0.05)。
2.ECH对db/db小鼠血清脂质水平的影响:血清脂质代谢水平检测结果显示,与db/m组小鼠比较,db/db组小鼠血清TC、TG、FFA、LDL-C显著升高,HDL-C明显降低(P<0.01,表3)。而经ECH灌胃治疗后,db/db+ECH组小鼠血清TC、TG、FFA、LDL-C水平显著降低,HDL-C明显升高(P<0.01)。

表2 ECH对db/db小鼠体质量和心脏湿重的影响
与db/m组比较,*P<0.01;与db/db组比较,#P<0.05,##P<0.01

表3 ECH对db/db小鼠血清脂质代谢的影响
与db/m组比较,*P<0.01;与db/db组比较,#P<0.01
3.ECH对db/db小鼠心脏病理形态变化的影响:实验结束时,固定部分左心室心肌组织进行HE染色和苏丹红Ⅱ染色,各组光镜下心肌组织结构变化如图2所示。HE染色显示db/m组心脏正常,心肌细胞排列清晰致密,结构清晰,细胞外基质及纤维组织少。而db/db组心肌细胞明显肥大坏死,心肌细胞排列紊乱,细胞外基质沉积较少。与db/db组比较,db/db+ECH组心肌细胞形态减轻,细胞间隙缩小,排列规则,细胞外基质沉积改善。苏丹红Ⅱ染色显示,db/m组心肌细胞脂滴较少,无脂质变性。db/db组小鼠心肌细胞发生严重的脂质变性,可见不同大小的脂质液泡。治疗后小鼠心肌细胞脂质沉积明显减少。

图2 ECH对db/db糖尿病小鼠心肌组织病理形态学变化的影响(×200)
4.各组db/db小鼠中PPAR-α、M-CPT-1、CD36和GLUT-4表达水平的变化:本研究分别通过免疫蛋白印迹和RT-PCR技术检测PPAR-α、M-CPT-1及其下游关键因子GLUT-4、CD36的蛋白和mRNA表达水平。实验结果表明, 相较于db/m组,db/db组小鼠PPAR-α、M-CPT-1、GLUT-4蛋白质和mRNA水平显著降低,CD36的表达明显增加(P<0.05)。经过ECH灌胃10周干预治疗后,PPAR-α介导的脂质代谢通路激活,PPAR-α、M-CPT-1、GLUT-4蛋白质和mRNA水平显著增加,CD36的表达明显减少(P<0.01,图3、图4)。

图3 各组小鼠心肌细胞PPAR-α、M-CPT-1、CD36和GLUT-4蛋白表达水平与db/m组比较,*P<0.01;与db/db组比较,#P<0.01

图4 各组小鼠心肌细胞PPAR-α、M-CPT-1、CD36和GLUT-4 mRNA水平与db/m组比较,*P<0.05,**P<0.01;与db/db组比较,#P<0.05,##P<0.01
讨 论
糖尿病患者长期高血糖状态所致的糖尿病心肌病早期病理改变表现为心肌细胞代谢紊乱和简单的心脏组织结构变化,后由于左心室心肌细胞体积、心室壁厚和重量增加、舒张功能降低,逐渐发展为心肌细胞凋亡、坏死和肥大、纤维化,导致明显的舒张和收缩功能障碍,最终可能发展成缺血性心脏病和心力衰竭[5, 10]。在本研究中,db/db组小鼠的血糖和血脂水平随着实验的进行而显著升高,db/db组小鼠表现出明显的肥胖症状,体质量较db/m组明显增加。db/db组心肌细胞HE染色显示明显的病理变化,如心肌细胞明显肥大坏死,排列紊乱,细胞外基质少量沉积。苏丹红Ⅱ染色结果显示,db/db组小鼠心肌细胞发生严重的脂质变性,可见不同大小的脂质空泡。这些结果都表明,db/db小鼠在长期高血糖状态下表现出早期糖尿病心肌病症状。
心肌细胞脂质堆积是糖尿病心肌病中一个重要的细胞损伤过程,加重了糖尿病心肌病心力衰竭的进程。PPAR-α被认为是多种脂肪酸氧化系统相关基因的关键调节因子,在脂质代谢、细胞炎症和免疫调节中发挥重要作用[11]。其中PPAR-α激活和改善细胞脂质代谢的主要机制包括:①控制多种线粒体和过氧化物酶体脂肪酸β氧化的关键酶, 如脂肪酸氧化限制酶、肉毒碱棕榈酰转移酶-Ⅰ(carnitinepalmitoyltransferase-1, CPT -Ⅰ)等;②促进长支链脂肪酸的氧化[12, 13];③激活后调节脂蛋白的合成和组装,降低VLDL的含量[14];④激活后间接调节胆汁酸合成,促进胆固醇排泄[15];⑤激活后抑制炎症基因的表达等。
CPT-1作为PPAR-α的下游关键基因共同参与了心肌细胞长链脂肪酸的新陈代谢, 抑制心肌细胞的脂质毒性[12,14]。ATP是心脏正常运转的基本能量来源,成人心脏功能主要依赖于游离脂肪酸(free acid,FA)的氧化分解,约占心脏能量供应的70%[16]。其余能量供应来源于碳水化合物,如葡萄糖和乳酸[17]。糖尿病患者心脏组织能量消耗的增加和心肌供能之间的不平衡能力是心肌代谢障碍的重要原因,主要是葡萄糖代谢供能的降低和FA新陈代谢的增加,导致心肌组织过度摄入FA氧化供能,心肌内脂肪代谢产物积累,增加心肌细胞脂毒性,最终导致糖尿病心肌病的发生、发展[18]。
本研究中笔者观察到在db/db糖尿病小鼠心肌细胞CD36表达相较于db/m小鼠显著增加,表明db/db小鼠存在心肌细胞摄取过多的FA, 而由于M-CPT-1表达水平显著降低,无法及时清除脂质代谢产物,故导致心肌细胞脂质堆积严重。另一方面,在糖尿病心肌病小鼠中, 由于PPAR-α表达被抑制,下游葡萄糖代谢关键基因GLUT-4表达水平降低,心肌葡萄糖氧化供能减少。为了维持心肌组织的正常供能,代偿机制势必会增加心肌细胞对FA的摄取,这也可以从另一个角度解释CD36在心肌细胞中过表达的原因。
经过ECH 10周干预治疗后,笔者观察到db/db+ECH小鼠的体质量明显低于db/db小鼠。虽然没有降低到与db/m组相同的血糖水平,但与db/db组比较,ECH能够显著改善db/db小鼠高血糖状态。HE染色和苏丹红Ⅱ染色显示,与db/db组比较,ECH干预后db/db小鼠心肌细胞形态改善,细胞间隙缩小,排列较规则,细胞外基质沉积改善,心肌细胞脂质积累明显减少。
ECH对db/db型糖尿病心肌病小鼠的脂质代谢也有明显的改善作用。在本研究中,从生化参数中积累的证据表明,ECH对DCM小鼠高血脂水平还具有改善作用。通过分析免疫印迹和RT-PCR的检测结果,可以确认ECH通过调控PPAR-α表达水平改善心肌细胞脂质积累的调控机制主要包括:①激活下游基因M-CPT-1 表达,抑制CD36基因的表达,从而抑制心肌细胞的FA过量摄入;②上调GLUT-4基因的表达,促进心肌细胞对葡萄糖的摄取和利用,改善心肌细胞糖脂代谢平衡。
综上所述,db/db糖尿病小鼠存在心肌细胞糖脂代谢紊乱。本研究证实,ECH可显著降低db/db小鼠的长期高血糖状态,改善心肌细胞糖脂代谢紊乱的潜在机制可能与PPAR/M-CPT-1信号通路有关。虽然本研究为糖尿病性心肌病的治疗提供了新的方向,但是ECH对改善DCM心肌损伤的保护作用还需要开展更深入的研究来进一步验证。
猜你喜欢
食品安全导刊(2020年14期)2020-12-04
食品安全导刊·中旬刊(2020年5期)2020-06-04
中国临床医学影像杂志(2019年1期)2019-04-25
现代食品(2018年16期)2018-11-02
中成药(2018年9期)2018-10-09
健康博览(2017年12期)2018-02-06
中成药(2018年1期)2018-02-02
中成药(2017年4期)2017-05-17
中国当代医药(2015年31期)2015-03-01
中国医药科学(2015年15期)2015-02-27
