
不同产量八角林土壤的生物学性状与细菌多样性特征
2020-05-25 02:59任奎瑜伍思宇梁文汇杨尚东
西南农业学报 2020年12期
肖 健,任奎瑜,伍思宇,梁文汇*,杨尚东*
(1.广西大学农学院,广西 南宁 530004;2.广西林业科学研究院/广西特色经济林培育与利用重点实验室/国家林业和草原局八角肉桂工程技术研究中心/广西木本香料工程技术研究中心,广西 南宁 530002)
【研究意义】八角(IlliciumverumHook.f)又称大茴香,俗称大料,是八角茴香科(Illiciaceae)八角属(IlliciumLinn.)常绿阔叶乔木,原产于我国西南地区和越南等地的亚热带地区,以果入药具有温中开胃及祛寒等疗效[1-2],其果皮、种子、叶片、枝梢和果实均含有丰富的茴香油[3],在工业上主要用于提炼大茴香[4],并进一步合成大茴香醇和大茴香醛等系列香料,用于食品、医药、啤酒、香皂和香水等产业。八角茴香油对细菌和真菌均具有一定的抗菌作用和抗氧化活性[5-6],可用于提炼莽草酸制造抗禽流感药物达菲[7]。广西既是八角的原产地也是主产区,有世界八角之乡的美称,但现阶段广西的大部分八角林管理粗放,导致八角产量大小年现象突出,产量和品质波动大,严重阻碍了广西八角产业的可持续发展。土壤生物学性状及细菌多样性特征可用于指示土壤的健康和肥力状况[8],但目前对低产八角林与其根际土壤生物学性状及细菌多样性是否存在某种关联,以及低产八角林根际微生物是否诱导八角林产量差异的了解甚少。因此,分析不同产量八角林土壤生物学性状及细菌的多样性特征,对改良低产八角林土壤生态环境,构建稳定、高产的八角林栽培管理技术体系具有重要意义。【前人研究进展】广西林业科学研究院通过多年研究发现,适宜八角生长的土壤pH为4.5~6.5,土壤施用、叶面喷施稀土和硼砂均能不同程度提高八角产量和促进林木生长[9]。已有研究发现,八角喜酸性土壤环境,适宜土壤pH为4.5~5.5,对土壤养分的要求为:有机质2.0 %,速效氮100 mg/kg,速效磷30 mg/kg,速效钾90 mg/kg[10]。也有相关研究提出了我国八角产业的优势区域布局[11]。袁霞和何斌[12]调查发现,土壤酶活性可作为评价八角林地土壤肥力的指标之一,提高土壤酶活性和增加土壤生物学肥力对提高八角林地土壤有机质含量和有效养分供应水平均具有重要作用。已有研究表明,土壤微生物与其他作物的增产有一定相关性,如王帅帅等[13]研究发现,不同防除杂草模式能通过改善土壤细菌占比来提高大葱产量和改善大葱根际土壤的健康状况;赵亚丽等[14]研究显示,土壤微生物量和酶活性可通过影响土壤肥力来影响作物产量。【本研究切入点】广西八角栽培中存在盲目施肥、缺乏规范化管理及生产出的八角品质参差不齐和产量不稳定等现象,急需通过系统研究八角林土壤的生态系统来提高八角产量和品质,但目前关于影响八角林土壤健康及土壤肥力持续平衡供给状况的研究鲜见报道。【拟解决的关键问题】以高产和低产八角林为研究对象,分析其土壤生物学性状和细菌多样性特征,从土壤健康和肥力状况角度系统地探究影响八角产量的土壤因素,为广西八角林的优质、稳产和高产栽培提供理论依据和技术支持。
1 材料与方法
1.1 采样地概况
采样点广西梧州市藤县古龙镇合隆村位于东经110°52′02″、北纬23°37′21″,属亚热带季风气候,年均气温21 ℃,日照1721.90 h,年均≥10 ℃活动积温7471.3 ℃,适宜种植八角。土壤类型为砂页岩母质发育而成的赤红壤,pH 5.6,全氮含量0.60 g/kg,全磷含量0.48 g/kg,全钾含量14.40 g/kg,速效磷含量5.00 mg/kg,速效钾含量50.00 mg/kg。
1.2 样品采集与处理
2019年7月28日采集高产八角林(品种为柔枝红花八角,14年生,产量17 010 kg/hm2)和低产八角林(品种为柔枝红花八角,14年生,产量8370 kg/hm2)0~30 cm土层的土壤样品,每种类型八角林土壤随机选择6个取样点,用无菌袋收集后装入放有冰袋的冰盒中带回实验室,将每份土壤样品分为两部分,一部分在室内风干后过40目筛,用于土壤理化性质测定,另一部分过10目筛后,分别用于土壤生物学性状和细菌群落结构分析。
1.3 土壤生物学性状分析
土壤碳、氮和磷循环相关酶β-葡糖苷酶活性采用Hayano和Koichi[15]的方法进行测定;氨肽酶活性参考Ladd[16]的方法进行测定;磷酸酶活性参考Tabatabai和Bremner[17]的方法进行测定。土壤微生物生物量碳、氮和磷含量分别采用氯仿熏蒸提取—容量分析法[18]、茚三酮比色法[19]和磷钼蓝比色法[20]测定。
1.4 土壤细菌多样性分析
土壤细菌多样性分析由上海美吉生物医药科技有限公司测序完成。具体测序流程:参照FastDNA®Spin Kit for Soil试剂盒(MP Biomedicals,U. S.)说明进行总DNA抽提,使用NanoDrop2000检测DNA浓度和纯度,用1 %琼脂糖凝胶电泳检测DNA提取质量;用338F和806R引物对V3~V4可变区进行PCR扩增。Illumina Miseq测序:回收PCR产物(2 %琼脂糖凝胶检测)→纯化(Extraction Kit)→Tris-HCl洗脱→电泳检测→检测定量(QuantiFluorTM-ST)→构建PE2×300文库。其中,构建文库步骤为:①连接“Y”字形连接器;②用磁珠过滤,自连接片段;③通过PCR扩增文库模板;④氢氧化钠变性,产生单链DNA片段。利用Illumina Miseq测序公司MiseqPE300平台进行测序,原始数据上传至NCBI数据库进行在线比对。本研究选用Ace指数、Chao指数、Simpson指数和Shannon指数分析不同产量八角林的土壤细菌多样性。参考Chao等[21]的方法计算Chao指数,参考Grice等[22]的方法计算Ace指数和Shannon指数:参考Simpson[23]的方法计算Simpson指数, Chao1、Ace和Shannon指数越大,Simpson指数越小,说明样品的物种丰富度和多样性越高。
1.5 统计分析
试验数据采用Excel 2019和SPSS 21.0进行统计,并以上海美吉生物医药科技有限公司的I-sanger云数据分析平台进行在线分析。
2 结果与分析
2.1 不同产量八角林的根际土壤生物学性状特征
2.1.1 土壤酶活性特征 由表1可知,低产八角林土壤的β-葡糖苷酶活性虽然高于高产八角林,但二者间差异不显著(P>0.05,下同);磷酸酶活性低于高产林土壤,但二者间差异不显著;氨肽酶活性显著高于高产八角林土壤14.4 %。说明低低产八角林土壤中存在较高产八角林土壤更活跃的氮素循环过程,其更高活性的氨肽酶活性可能会导致低产八角林土壤中持续的氮素供给能力逊于高产八角林土壤的现象。
2.1.2 土壤微生物生物量碳、氮和磷含量 土壤微生物生物量及其变化情况是表征土壤肥力及其变化情况的重要依据之一。由表2可知,高产八角林土壤的微生物生物量碳、氮和磷含量均高于低产八角林,分别为低产八角林的1.64、1.73和1.23倍。其中,高产八角林土壤的微生物生物量氮含量明显高于低产八角林,微生物生物量碳和磷含量均显著高于低产八角林。表明高产八角林土壤肥力比低产八角林更高,且是八角产量更高的重要保障。
2.2 不同产量八角林的根际土壤细菌多样性分析
2.2.1 Alpha多样性 由表3可知,基于高通量测序技术分析,八角林土壤样品中的细菌群落隶属于28门74纲185目317科565属1162种3981 OTU。其中,高产八角林土壤的细菌群落隶属于27门70纲177目304科538属1080种3366 OTU,低产八角林土壤的细菌群落隶属于27门71纲174目294科520属1047种3460 OTU。说明高产八角林土壤细菌的丰富度大于低产八角林。
2.2.2 丰富度和多样性 基于I-sanger云分析平台,计算出不同产量八角林土壤细菌的丰富度、多样性指数和覆盖率。以Chao1和Ace指数指示细菌的丰富度,Shannon和Simpson指数指示细菌的多样性,Coverage指数指示细菌的覆盖率。由表4可知,高产八角林土壤中指示细菌丰富度的Ace和Chao1指数高于低产八角林,但二者间差异不显著;高产八角林土壤中指示细菌多样性的Shannon指数显著高于低产八角林,而Simpson指高于低产八角林,但二者间无显著差异。表明八角林的土壤细菌丰富度对八角产量的影响可能逊于细菌多样性的影响,更丰富的细菌多样性可能是高产八角林产出更高产量的重要原因之一。

表1 不同产量八角林的土壤酶活性比较

表2 不同八角林的根际土壤微生物生物量比较

表3 不同产量八角林根际土壤样品的物种丰富度统计结果
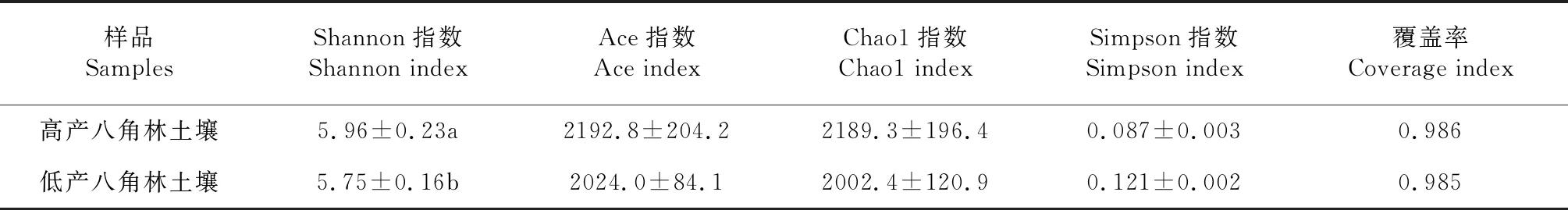
表4 不同产量八角林的土壤细菌丰富度和多样性指数比较
2.3 群落组成分析结果
2.3.1 门分类水平的优势菌群分析 从图1-A和图1-B可看出,在门分类水平上,高产和低产八角林土壤的优势细菌门分别有10和11个。其中,高产八角林土壤的优势细菌在门分类水平上占比排序为变形杆菌门(Proteobacteria,36.06 %)>放线菌门(Actinobacteria,21.80 %)>酸杆菌门(Acidobacteria,16.67 %)>绿弯菌门(Chloroflexi,10.29 %)>浮霉菌门(Planctomycetes,3.44 %)>疣微菌门(Verrucomicrobia,3.10 %)>拟杆菌门(Bacteroidetes,1.95 %)>WPS-2(1.90 %)>芽单胞菌门(Gemmatimonadetes,1.60 %),其他门(others)占3.20 %(图1-A);低产八角林土壤的优势细菌在门分类水平上占比排序为变形杆菌门(Proteobacteria,31.23 %)>放线菌门(Actinobacteria,21.52 %)>绿弯菌门(Chloroflexi,15.99 %)>酸杆菌门(Acidobacteria,15.22 %)>浮霉菌门(Planctomycetes,3.44 %)>疣微菌门(Verrucomicrobia,2.71 %)>WPS-2(2.47 %)>Patescibacteria(1.47 %)>芽单胞菌门(Gemmatimonadetes,1.37 %)>厚壁菌门(Firmicutes,1.34 %)=拟杆菌门(Bacteroidetes,1.34 %),其他门(others)占1.89 %(图1-B)。可见,高产和低产八角林土壤的优势细菌在门分类水平上占比存在明显差异,主要表现在低产八角林土壤优势细菌在门分类水平上的变形杆菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes)和其他门细菌占比分别较高产八角林土壤下降4.83 %、1.45 %、0.61 %和1.31 %(绝对值,下同),而绿弯菌门(Chloroflexi)细菌占比急剧上升至15.99 %,较高产八角林土壤提高5.70 %;同时,Patescibacteria门和厚壁菌门(Firmicutes)细菌亦由高产八角林土壤中的非优势细菌门,转变成低产八角林土壤中的优势细菌门。由此推测,低产八角林土壤中优势细菌在门分类水平上占比及排序的变化,尤其是其他门细菌占比的下降,可能是低产八角林土壤细菌多样性下降的重要原因。
2.3.2 属分类水平的优势菌群分析 在属分类水平上,高产八角林土壤的优势细菌占比排序为未分类纱丽杆菌科(norank_f__Xanthobacteraceae,9.41 %)>嗜酸栖热菌属(Acidothermus,8.29 %)>norank_o__Elsterales(3.88 %)>norank_o__Subgroup_2(3.87 %)>norank_f__JG30-KF-AS9(3.66 %)>未分类酸杆菌属(norank_o__Acidobacteriales,3.31 %)>norank_c__AD3(3.23 %)>Candidatus_solibacter(2.98 %)>norank_p__WPS-2(2.47 %)>norank_o__IMCC26256(2.41 %)>慢生根瘤菌属(Bradyrhizobium,2.38 %)>不动杆菌属(Acidibacter,2.19 %)>norank_f__Gemmataceae(2.17 %)>Conexibacter(1.94 %)>未分类放线菌属(norank_c__Actinobacteria,1.74 %)=HSB_OF53_F07(1.74 %)>norank_c_TK10 (1.31 %)>FCPS473(1.27 %),其他属(others)占比36.04 %(2-A);低产八角林土壤的优势细菌占比排序为未分类纱丽杆菌科(norank_f__Xanthobacteraceae,11.67 %)>嗜酸栖热菌属(Acidothermus,6.53 %)>norank_o__Elsterales(4.04 %)>Candidatus_solibacter(3.71 %)>norank_o__Subgroup_2(3.59 %)>慢生根瘤菌属(Bradyrhizobium,3.14 %)>未分类酸杆菌属(norank_o__Acidobacteriales,2.87 %)>norank_o__IMCC26256(2.75 %)>norank_f__JG30-KF-AS9(2.74 %)>norank_f__Gemmataceae(2.32 %)>不动杆菌属(Acidibacter,2.00 %)>Conexibacter(1.98 %)>norank_p__WPS-2(1.90 %)>未分类放线菌属(norank_c__Actinobacteria,1.84 %)>norank_o__Gaiellales(1.72 %)>norank_f__67_14(1.65 %)>Candidatus_Xiphinematobacter(1.57 %),其他属(others)占比35.74 %(图2-B)。可见,与高产八角林土壤相比,低产八角林土壤的优势细菌在属分类水平上占比发生了明显变化,其中除排序前3位的优势细菌未发生变化外,高产林土壤中优势细菌占比排第4位的属由原来的norank_o__subgroup_2变成低产八角林土壤中的Candidatus_Solibacter;低产八角林土壤的部分优势细菌属缺失,如norank_c__AD3(3.23 %)、HSB_OF53_F07(1.74 %)、norank_c_TK10(1.31 %)和FCPS473(1.27 %)等,以及其他属(others)细菌占比下降。由此推测,优势细菌群落结构发生变化和细菌多样性下降可能是低产八角林产量低的原因之一。

图1 高产八角林土壤(A)和低产八角林土壤(B)在门分类水平上的细菌组成

图2 高产八角林土壤(A)和低产八角林土壤(B)在属分类水平上的细菌组成

图3 高产八角林土壤和低产八角林土壤细菌的属分类水平(A)和种分类水平(B)Venn图
此外,由属分类水平及种分类水平Venn图可看出,在属分类水平(图3-A)上,高产和低产八角林土壤的共有细菌属为490个,在低产八角林土壤中特有的细菌属为30个,而高产八角林土壤中特有的细菌属为48个;在种分类水平(图3-B)上,高产和低产八角林土壤的共有细菌种为962个,低产八角林土壤中特有的细菌种为85个,而高产八角林土壤中特有的细菌种达118个。由此推测,土壤中更丰富的细菌种和属分类水平,亦可能是高产八角林产量高于低产八角林的原因之一。
3 讨 论
种植八角是我国南方山区群众经济收入的主要来源和脱贫致富的有效途径[13]。广西八角栽培面积和产量占全国的90 %以上,其中,广西藤县种植八角历史悠久,近年来虽已加强管理和注重新技术引进与应用,但由于树龄老化或环境恶化等原因仍出现八角产量下降、品质劣化等不良现象[14,24]。已有研究证实,土壤酶参与土壤中几乎所有营养元素的循环[25],其活性不仅能指示土壤生态系统功能变化的多样性和稳定性[26],亦是评价土壤健康和肥力水平的重要指标[27]。本研究发现,低产八角林土壤的β-葡糖苷酶和氨肽酶活性虽然高于高产八角林,但仅氨肽酶活性显著高于后者,且土壤磷酸酶活性无显著差异。由此推测,低产八角林土壤中部分氮素循环虽然强于高产八角林,但作为氮源生物学性状指标的微生物生物量氮含量差异不显著,表明低产八角林土壤与高产八角林土壤相比,极可能会出现持续供氮能力不足现象。土壤微生物生物量同样是衡量土壤质量、维持土壤肥力和作物生产力的一个重要指标[28]。土壤中微生物生物量越大,土壤通过有机养分矿化提供给植物所需养分的能力就越强[29]。其中,土壤微生物生物量碳不仅可促进土壤形成活性较高的新生腐殖质,而且还可在土壤全碳含量变化前反映土壤的微小变化[30];土壤微生物生物量氮能反映土壤氮素的有效性,在土壤氮的供应和循环中发挥重要作用[30];土壤微生物生物量磷可反映土壤磷素的供应水平[31]。此外,微生物生物量磷虽然不能被植物直接吸收利用,但能缓慢释放无机磷,因此一直被认为是土壤中的有效磷来源,对于植物生长非常重要[32]。本研究中,低产八角林土壤中微生物生物量碳和磷含量均显著低于高产八角林,但二者的土壤微生物生物量氮间无显著差异。由此推断,低产八角林土壤的有机碳含量较低、持续供给养分能力逊于高产八角林,尤其是土壤磷素持续供应能力不足可能是其低产的重要原因之一。
已有研究证实,土壤微生物群落结构越丰富、物种越均匀及多样性越丰富,植物对抗病原菌的综合能力就越强[33-34]。本研究基于高通量测序技术分析发现,低产八角林土壤的细菌丰富度虽然与高产八角林无显著差异,但Shannon指数显著低于后者,由此推测,低产八角林土壤中显著贫乏的细菌多样性可能导致其土壤生态系统功能降低、养分供给平衡紊乱等影响产量的障碍因素发生概率升高,从而出现产量不高现象;基于门分类水平分析发现,与高产八角林土壤相比,低产八角林土壤的变形杆菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes)和其他门细菌占比下降,绿弯菌门(Chloroflexi)细菌、Patescibacteria门和厚壁菌门(Firmicutes)细菌占比急剧上升。其中,群落组成占比急剧上升的绿弯菌门细菌是一个新的系统发育分支,虽然其在环境中普遍存在,但主要生活在深海、湖泊及淹水的泥滩等厌氧环境[35-36],尤其是高氮低碳环境更有利于其生存和大量繁殖[37];厚壁菌门细菌是一个大类细菌,可降解多种有机污染物,如多氯联苯和石油烃等,且其在贫瘠土壤中丰度更大[38]。由此推测,氮的非均衡供应及碳、磷供给不足,可能均是导致低产八角林产量不高的重要原因;基于属分类水平分析发现,与高产八角林土壤相比,低产八角林土壤的优势细菌占比同样发生明显变化,尤其是Candidatussolibacter属细菌占比上升,部分优势细菌属缺失及其他属(other)细菌占比下降,细菌群落与功能多样性降低,也是低产八角林产量不高的重要原因。
4 结 论
与高产八角林相比,低产八角林土壤中氮的非均衡供应、碳和磷储备偏低,以及细菌多样性下降,部分优势细菌属缺失导致的细菌群落结构失衡,均可能是低产八角林八角低产量的重要原因。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
农业知识(2022年9期)2022-10-13
今日农业(2022年4期)2022-06-01
今日农业(2021年19期)2022-01-12
今日农业(2021年13期)2021-11-26
今日农业(2021年16期)2021-11-26
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
农民致富之友(2018年11期)2018-06-28
