
亚洲玉米螟卵黄原蛋白基因的克隆、表达谱及对UV-A胁迫的响应
2020-05-22 06:18孟建玉杨昌利张长禹
昆虫学报 2020年3期
刘 芳, 孟建玉, 杨昌利, 张长禹,*
(1. 贵州大学昆虫研究所, 贵州省山地农业病虫害重点实验室, 贵阳 550025;2. 贵州省烟草科学研究院, 贵阳 550081)
卵黄原蛋白(vitellogenin, Vg)由脂肪体合成,特异存在于性成熟的卵生动物雌性血液中的一种蛋白,是卵黄磷蛋白(vitellin, Vt)的前体(李继莲等, 2012)。昆虫Vg具有几个保守结构域,最显著的是N末端的多聚丝氨酸区,其被认为是合适的丝氨酸磷酸化位点(Tufail and Takeda, 2008)。作为昆虫卵黄发生的关键物质,Vg是具有明确生理功能的蛋白,最重要的特点是能为胚胎发育提供氨基酸、脂肪和维生素等(Giorgietal., 1999)。近年的研究则表明Vg具有多种生物学功能,例如灰飞虱Laodelphaxstriatellus的Vg被水稻条纹病毒(RSV)利用而参与其复制,在传递RSV过程中起着关键作用(Huoetal., 2014),家蚕Bombyxmori的Vg能抑制细菌的繁殖(Singhetal., 2013),Vg还可以作为酶的载体发挥作用(钱岑, 2010)。Vg在东方蜜蜂Apiscerana等社会性昆虫中也具有多重特殊功能,因此被称为“多效性蛋白”(Nelsonetal., 2007),Vg除与个体的生理特点、行为反应和寿命等差异有关外(严盈等, 2010),还涉及蜜蜂的气候适应(Amdanetal., 2005)、激活卵巢(Koywiwattrakul and Sittipraneed, 2009)、社会行为调节(Münchetal., 2015)和可增强免疫力并参与免疫启动(Havukainenetal., 2011; Harwoodetal., 2018)等功能。
趋光性是众多夜行性昆虫的一个重要生态学特征。鳞翅目、半翅目和鞘翅目等多种昆虫的成虫都具有趋光性(靖湘峰等, 2005; 唐良德等, 2016; 徐练, 2016)。郭虹等(2012)和王林聪等(2016)研究表明,用UV诱虫灯诱集农林业害虫的效果显著,且敏感波长均集中在UV-A(320~400 nm)波段。在农业生产中,以UV-A为主要波段的诱虫灯普遍用于害虫的预测预报及物理防控。另一方面,UV-A辐射能够破坏生物的DNA 结构,导致基因突变等,给生物生存构成一种环境压力,即UV-A对昆虫而言也是一种环境胁迫因子(Kuhlmann and Mülle, 2010; Wölfieetal., 2011; Jietal., 2012; Zhouetal., 2018)。近年来的研究则发现,UV-A辐射会增强南极蠓Belgicaantarctica对氧化应激的抵抗力(Lopez-Martinezetal., 2008),刺激赤拟谷盗Triboliumcastaneum体内应激反应基因Hsps和P450的表达(Sangetal., 2012),增强棉铃虫Helicoverpaarmigera体内的抗氧化酶活性、改变保幼激素(JH)的代谢水平并激活其MAPK信号通路等(Mengetal., 2010; Wangetal., 2012; 刘小飞等, 2019)。
亚洲玉米螟Ostriniafurnacalis是典型的趋光性昆虫,属于鳞翅目草螟科,寄主范围广,严重威胁玉米等旱粮作物的产量和质量(王振营等, 2000; 王雪艳和刘晓玲, 2017)。关于亚洲玉米螟趋光的行为特点的研究表明其成虫对UV-A十分敏感(杨桂华和王蕴生, 1995; 徐练, 2016)。近期的研究则发现UV-A照射还会影响趋光昆虫的繁殖,棉铃虫成虫在响应UV-A胁迫后生殖力上升,出现生殖补偿现象(张长禹, 2010; 王智健和牛长缨, 2014)。朱亮等(2016)研究发现UV-A照射会促进烟粉虱Bemisiatabaci、西花蓟马Frankliniellaoccidentalis和桃蚜Myzuspersicae的种群增长。然而,关于UV-A如何影响亚洲玉米螟生殖尚未见报道。本研究克隆亚洲玉米螟Vg基因、分析其序列特征和表达谱,研究其在UV-A胁迫下的表达变化,测定UV-A胁迫后亚洲玉米螟的生殖力及子代的发育情况,为深入研究UV-A胁迫对亚洲玉米螟繁殖的影响奠定基础。
1 材料与方法
1.1 供试昆虫及试剂
所用亚洲玉米螟为本实验室长期饲养的昆虫,幼虫期用人工饲料饲养,饲料配方参照乔利等(2008)的配方,以蘸有蜂蜜水的棉球饲喂成虫,室内饲养条件为:温度27±1℃、相对湿度70%±5%、光周期14L∶10D。
主要试剂包括Eastep Super RNA提取试剂盒(上海普洛麦格生物产品有限公司);HiFiScript cDNA第1链合成试剂盒(北京康为世纪生物科技有限公司);Master Mix(2×)[生工生物工程(上海)有限公司];SanPrep柱式DNA胶回收试剂盒[天根生化科技(北京)有限公司];SMARTer RACE 5′/3′Kit User Manual试剂盒、pMD19-T Vector、大肠杆菌EscherichiacoliDH5α、Lineareized RACE Vector、Stellar Competent Cells及TB Green Premix DimerEraser荧光试剂等购自大连宝生物工程有限公司;所有引物送生工生物工程有限公司合成。
1.2 总RNA提取和cDNA的合成
取8头亚洲玉米螟雌成虫,用震荡研磨器磨碎后,采用Eastep Super试剂盒提取试虫的总RNA,用NanoPhotometer P-Class分光光度仪检测RNA的浓度,以1%琼脂糖凝胶电泳检测其质量。根据HiFiScript cDNA第1链合成试剂说明将提取的RNA进行反转录合成cDNA第1链。
1.3 亚洲玉米螟Vg基因的克隆
在NCBI(https:∥blast.ncbi.nlm.nih.gov/Blast.cgi)中下载已知昆虫同源Vg序列并比对,用Primer Premier 6.0软件在保守区内设计RT-PCR引物(表1),以1.2节合成的cDNA第1链为模板,PCR扩增亚洲玉米螟Vg基因中间片段。反应体系(25 μL): Master Mix (2×)酶12.5 μL, cDNA模板(300 ng/μL)3 μL, 正反向引物(10 mmol/L)各1 μL, ddH2O 7.5 μL。反应条件: 95℃ 3 min; 95℃ 30 s, 55℃ 30 s, 72℃ 1 min, 35个循环; 72℃ 10 min。PCR产物用1%琼脂糖凝胶电泳检测, 确定接头用下划线标出。The adaptor is underlined.

表1 本研究所用引物Table 1 Primers used in this study
目的条带后用SanPrep柱式DNA胶回收试剂盒进行纯化回收,连接到pMD19-T Vector上并转化至大肠杆菌DH5α感受态细胞,挑取单一阳性克隆经PCR检验后送菌液到生工生物工程有限公司测序。
根据已经扩增获得的亚洲玉米螟Vg基因片段序列,用Primer Premier 6.0设计5′/3′RACE特异引物(GSP)和嵌套特异引物(NGSP)(表1),引物前加上接头GATTACGCCAAGCTT。根据SMARTer RACE 5′/3′Kit User Manual试剂盒说明书分别进行3′RACE和5′RACE半巢式PCR扩增。25 μL反应体系按说明书加样。反应条件: 94℃ 30 s, 55℃ 30 s, 72℃ 2 min, 共28个循环。电泳检测分离、纯化回收后连接到Lineareized RACE Vector载体中,转化到Stellar Competent Cells,克隆并测序鉴定。
1.4 亚洲玉米螟Vg基因序列拼接与分析
将获得的序列用软件DNAMAN 6.0进行拼接,然后在序列两端设计上下游引物(表1)PCR扩增全长序列,反应体系(25 μL): SeqAmp PCR Buffer(2×) 12.5 μL, ddH2O 4.5 μL, cDNA模板(300 ng/μL)3 μL, 正反向引物(10 mmol/L)各2 μL, SeqAmp DNA Polymerase 1 μL。反应条件: 94℃ 30 s, 57.5℃ 30 s, 72℃ 3.5 min, 28个循环。单向测通后经NCBI中Blast工具进行比对确认后获得亚洲玉米螟Vg基因的全长序列,放入ORF Finder(http:∥www.ncbi.nlm.nih.gov/projects/gorf/)中分析其开放阅读框;使用ExPASy ProtParam(http:∥www.expasy.org/tools/protparam.html)对蛋白质的理化性质进行在线预测;采用 MEGA 6.0软件对22个物种Vg氨基酸序列用邻接法构建系统发育树。
1.5 亚洲玉米螟Vg基因的表达
用RT-qPCR对亚洲玉米螟不同发育阶段、不同组织及不同UV-A照射下的Vg基因的表达量进行分析。各组样品来源或处理如下:(1)不同发育阶段:卵、1-5龄幼虫、雌/雄蛹、雄成虫(羽化后0, 12, 24, 36, 48 和72 h)和雌成虫(羽化后0, 12, 24, 36, 48, 72, 96和120 h);(2)不同组织:羽化24 h雌成虫的头、足、表皮、卵巢、中肠和脂肪体;(3)UV-A照射:羽化后24 h的雌成虫暗适应2 h后,用UV-A(320-400 nm)灯管以300 μW/cm2的强度进行照射0(CK), 0.5, 1, 1.5, 2, 2.5, 3, 3.5和4 h,照射期间条件和饲养条件一致。每组样品或实验处理均设置3个生物学重复,每个生物学重复分别为:卵50粒,1龄幼虫30头,2龄幼虫15头,3-5龄幼虫、蛹和成虫各8头个体。提取试虫RNA后反转录(方法同1.2节),并以亚洲玉米螟actin基因(GenBank登录号: XM_028313197.1)为内参基因进行实时荧光定量PCR,反应体系(20 μL): cDNA模板(300 ng/μL)1 μL, TB Green Premix DimerEraser(2×)10 μL, 上下游引物(10 mmol/L)各1 μL, ddH2O 7 μL。每个生物学重复进行3个技术重复。 反应条件: 95℃ 30 s; 95℃ 5 s, 60℃ 30 s, 72℃ 30 s, 40个循环; 65℃ 5 s。
1.6 UV-A照射后亚洲玉米螟生殖及子代发育情况测定
蛹羽化后一雌一雄配对置于130 mL塑料杯中,每天暗期1 h后用UV-A灯管以300 μW/cm2的强度分别照射0(CK), 1, 2, 3和4 h,每天记录产卵量直到成虫死亡,每处理6对,重复3次。待雌虫开始产出大量的卵,各处理中挑选出同一天刚产出的卵100粒,进行饲养观察后,各处理中挑选出同一天刚孵出的幼虫60头,接着饲养后各处理中挑选出同一天刚化成蛹40头。观察卵、幼虫和蛹的存活数和发育历期,每个实验处理设置3个生物学重复,每个生物学重复分别为:卵100粒,幼虫60头,蛹40头。
1.7 数据分析
在Excel 2016中将所得CQ值利用2-△△CT法(Livak and Schmittgen, 2001)对亚洲玉米螟Vg基因的表达量进行相对定量分析。实验所得数据采用SPSS17.0进行处理,采用Duncan氏检验法对数据进行差异显著性分析(P<0.05)。卵孵化率、幼虫化蛹率和蛹羽化率在方差分析前先进行反正弦平方根转换。
2 结果
2.1 亚洲玉米螟Vg基因的克隆和序列分析
以亚洲玉米螟雌虫cDNA作为模板,用一对简并引物OfVg-F和OfVg-R扩增并克隆、测序得到1 013 bp序列,经Blast工具比对发现其核苷酸序列与其他同源昆虫的Vg序列具有较高的一致性,表明该序列为亚洲玉米螟Vg基因的片段序列。RACE扩增并克隆、测序分别得到5′端366 bp和3′端4 381 bp序列,DNAMAN拼接获得5 760 bp的全长序列,命名为OfVg(GenBank登录号: MK782978)。
OfVg基因开放阅读框(ORF)长5 331 bp,编码1 776个氨基酸,5′端非编码区和3′端非编码区分别为53和376 bp,具有加尾信号AATAAA。编码蛋白相对分子量为202.10 kD,等电点为9.06,分子式为C8961H14169N2499O2680S72;其中负电荷氨基酸残基数(Asp+Glu)为204,正电荷氨基酸残基数(Arg+Lys)为236;稳定性系数为43.15,说明该蛋白质性质相对稳定;总平均疏水指数为-0.558,表明其为亲水性蛋白质。NCBI-CDD蛋白保守结构域分析显示,亚洲玉米螟Vg蛋白包含已知昆虫Vg中普遍存在的3个功能结构域Vitellogenin-N(Vg-N, 第31-729位氨基酸)、DUF 1943D(第762-1 037位氨基酸)和von Willebrand factor type D domainvon(VWD, 第1 431-1 607位氨基酸);利用NetOGIyc工具得到20个O-糖基化位点(图1)。应用KinasePhos工具,分析得到磷酸化位点Serine(S)为128,Threonine(T)为90,Tyrosine(Y)为76;并用TMHMM在线预测,显示该蛋白无跨膜区结构。

图1 亚洲玉米螟Vg基因序列推导的氨基酸序列Fig. 1 Deduced amino acid sequences of Vg from Ostrinia furnacalis信号肽用双下划线标记;O-糖基化位点用单下划直线标记;阴影部分表示3个功能域。The signal peptide is double-underlined, the O-glycosylation site is single underlined, and the three functional domains are shaded.
2.2 亚洲玉米螟Vg的系统进化分析
在NCBI数据库中下载半翅目、膜翅目、鞘翅目、双翅目和鳞翅目共5个目20种昆虫及外群双须骨舌鱼Osteoglossumbicirrhosum的Vg氨基酸序列,并与亚洲玉米螟Vg推导的氨基酸序列一起采用NJ(neighbor-joining)法构建系统发育树(图2)。结果表明,进化树分为脊椎动物和昆虫纲(Insecta)两个大支,昆虫与外群明显区分。昆虫纲的半翅目、膜翅目、鞘翅目、双翅目和鳞翅目都分别聚在一个分支上,其中鳞翅目的亚洲玉米螟与半翅目、膜翅目、鞘翅目和双翅目昆虫亲缘关系较远;与鳞翅目稻纵卷叶螟Cnaphalocrocismedinalis、豆荚螟Marucavitrata和二化螟Chilosuppressalis、棉铃虫、甜菜夜蛾Spodopteraexigua、粉纹夜蛾Trichoplusiani、柑橘凤蝶Papilioxuthus、玉带凤蝶Papiliopolytes和柳蚕Actiasselene聚为一支,说明昆虫Vg具有较高的保守性,其亲缘关系较近;且与稻纵卷叶螟和豆荚螟在同一小分支,其亲缘关系最近。

图2 以邻接法构建的基于氨基酸序列的昆虫Vg系统发育树Fig. 2 Phylogenetic tree of insect Vg proteins constructed by neighbor-joining method based on amino acid sequences 发育树的分支置信度检测是利用Bootstrap进行1 000次计算,分支上的数值表示置信度。The topology of phylogenetic tree was tested by bootstrap analysis (1 000 replicates). The numerical values on the branches indicate the confidence level.
2.3 亚洲玉米螟Vg基因的表达模式分析
OfVg在亚洲玉米螟不同发育阶段的相对表达量分析结果显示,雌成虫中表达量显著高于卵、幼虫、蛹及雄成虫中表达量(P<0.05),且在幼虫、蛹和雄成虫中低表达;雌成虫中表达量先上升后下降,在羽化24 h时表达量最高(图3)。亚洲玉米螟雌成虫在羽化24 h时不同组织中OfVg的表达量存在显著差异,在脂肪体中表达量远远高于其他组织中表达量(P<0.05),在其他组织中仅少量表达(图4)。
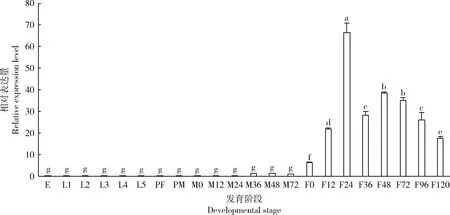
图3 OfVg在亚洲玉米螟不同发育阶段中的表达模式Fig. 3 Expression pattern of OfVg in different developmental stages of Ostrinia furnacalisE: 卵 Egg; L1-L5: 分别为1-5龄幼虫1st-5th instar larva, respectively; PF: 雌蛹Female pupa; PM: 雄蛹 Male pupa; M0-72: 分别为羽化后0-72 h的雄成虫 Male adult at 0-72 h after eclosion, respectively; F0-120: 分别为羽化后0-120 h的雌成虫Female adult at 0-120 h after eclosion, respectively. 不同发育阶段间的基因表达量以羽化后72 h的雄成虫中表达量为基准。图中数据为平均值±标准误,柱上不同字母表示不同发育阶段间的表达量存在显著性差异(P<0.05, Duncan氏检验)。The expression levels of gene in different developmental stages were normalized to that in male adult at 72 h after eclosion. Data in the figure are mean±SE, and different letters above bars indicate significant differences in the expression level between different developmental stages (P<0.05, Duncan’s test).
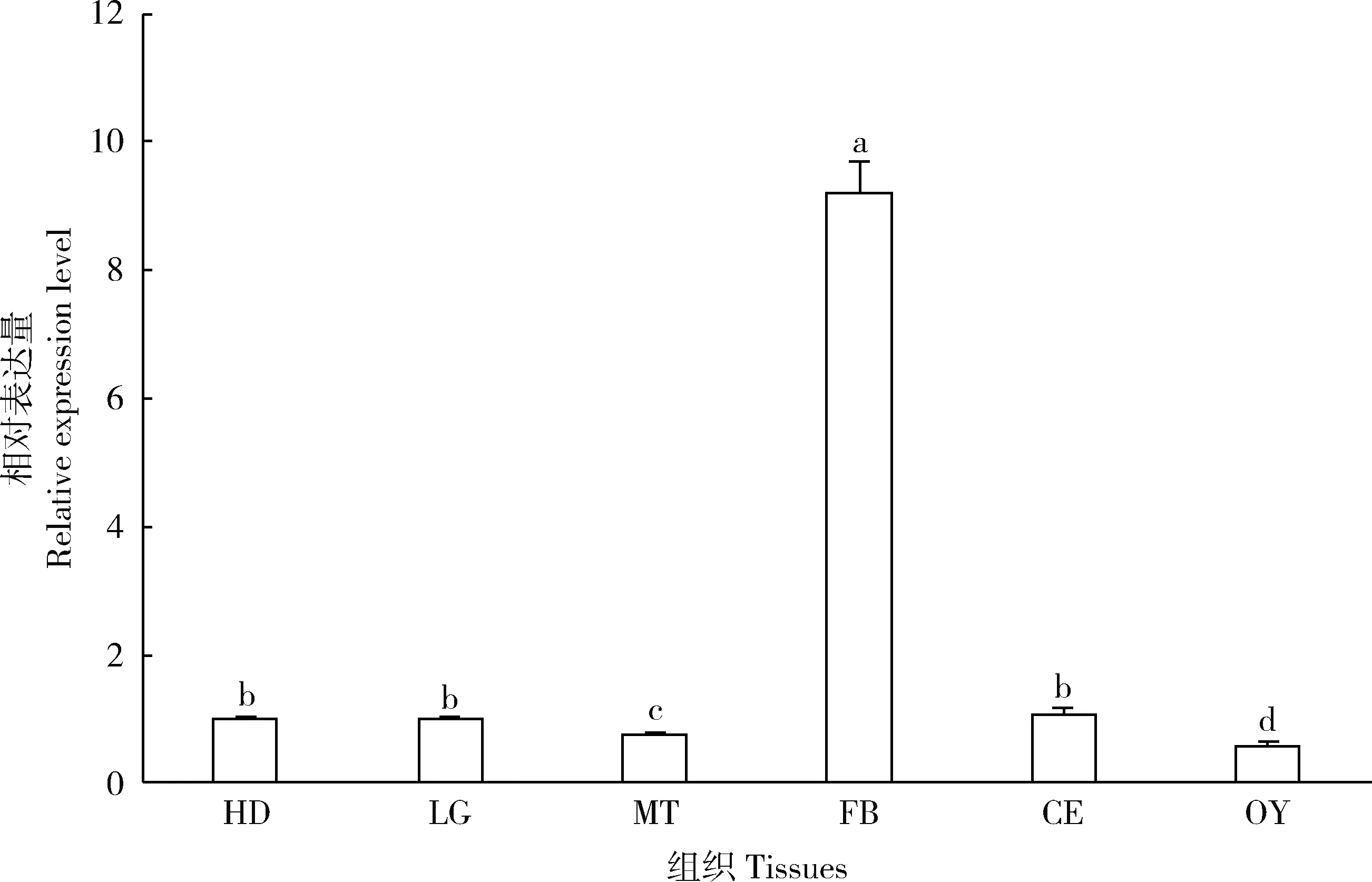
图4 OfVg在亚洲玉米螟雌成虫不同组织中的表达模式Fig. 4 Expression pattern of OfVg in different tissues of female adults of Ostrinia furnacalisHD: 头Head; LG: 足Leg; MT: 中肠 Midgut; FB: 脂肪体Fat body; CE: 表皮Cuticle; OY: 卵巢Ovary. 不同组织中的表达量以头中表达量为基准。图中数据为平均值±标准误,柱上不同字母表示不同组织间的表达量存在显著性差异(P<0.05, Duncan氏检验)。The expression levels of gene in different tissues were normalized to that in the head. Data in the figure are mean±SE, and different letters above bars indicate significant differences in the expression level between different tissues (P<0.05, Duncan’s test).
羽化24 h的玉米螟雌成虫在不同时长UV-A胁迫下,OfVg的表达量随照射时间的延长呈先缓慢下降后快速上升、再骤降的趋势;与对照(照射0 h)相比,照射时间为0.5, 1和1.5 h时,表达量均下降,在2, 2.5, 3和3.5 h时表达量逐渐快速升高,在3.5 h时达到最高(P<0.05),是对照组表达量的3.62倍(图5)。当照射时间延长至4 h时,表达量又骤降到对照组水平以下(P<0.05)。
2.4 UV-A照射对亚洲玉米螟生殖力及子代发育的影响
UV-A处理1, 2和 3 h/d雌成虫的总产卵量和每日单雌产卵量均显著高于未照射处理的对照组(P<0.05),且均随照射时间延长先增多后减少,在3 h/d达到最大值(表2)。
UV-A照射对亚洲玉米螟卵孵化率、化蛹率均没有显著影响。UV-A处理组子代蛹的羽化率均显著降低(P<0.05),且照射4 h/d的蛹羽化率最低。UV-A照射后子代累积存活率都明显低于对照组(表3)。UV处理组1, 2和 3 h/d子代幼虫的发育历期都显著长于对照组(P<0.05),在3 h/d时最长,达26.11 d;UV-A处理组子代蛹的发育历期与对照组无显著差异(P>0.05)(表4)。

图5 OfVg在亚洲玉米螟雌成虫不同时长UV-A胁迫下的表达模式Fig. 5 Expression pattern of OfVg in female adults of Ostrinia furnacalis exposed to UV-A for different time羽化后24 h的雌成虫暗适应2 h后,用UV-A(320-400 nm)灯管以300 μW/cm2的强度进行照射0(CK), 0.5, 1, 1.5, 2, 2.5, 3, 3.5和4 h; 雌成虫在不同UV-A照射时间的表达量以羽化24 h的未处理的对照(0 h)中的表达量为基准。图中数据为平均值±标准误,柱上不同字母表示不同照射时间之间的表达量存在显著性差异(P<0.05, Duncan氏检验)。The female adults at 24 h after eclosion were acclimated to darkness for 2 h, and then exposed to 300 μW/cm2 of UV-A for 0 (CK), 0.5, 1, 1.5, 2, 2.5, 3, 3.5 and 4 h, respectively. The expression levels of OfVg in female adults exposed to UV-A for different time were normalized to that in the untreated control (0 h). Data in the figure are mean±SE, and different letters above bars indicate significant differences in the expression level between different exposure time (P<0.05, Duncan’s test).
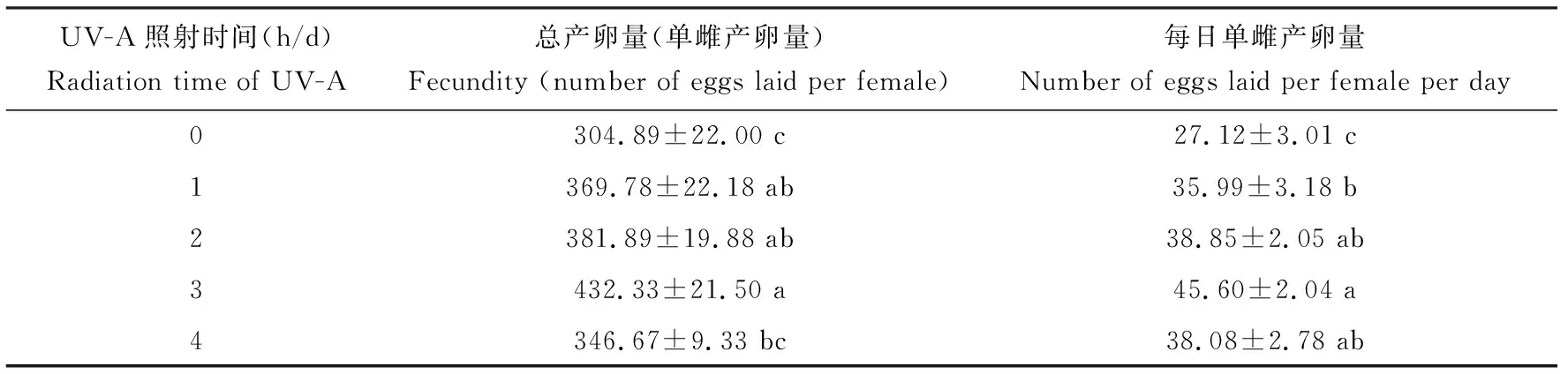
表2 UV-A照射对亚洲玉米螟雌成虫繁殖的影响Table 2 Effect of UV-A radiation on the reproduction of female adults of Ostrinia furnacalis
蛹羽化后1雌1雄配对置于塑料杯中,每天暗期1 h后用UV-A灯管以300 μW/cm2的强度分别照射0(CK), 1, 2, 3和4 h,每天记录产卵量直到成虫死亡。表中数据为平均值±标准误,同列数据后的不同字母表示差异显著(P<0.05, Duncan氏检验)。One female and one male after pupal eclosion were paired in plastic cups, and exposed to 300 μW/cm2of UV-A for 0 (CK), 1, 2, 3, and 4 h/d, respectively. The daily number of eggs laid was recorded until adult death. Data in the table are mean±SE, and different letters after the data in the same column indicate significant differences (P<0.05, Duncan’s test). 表3和表4同The same for Tables 3 and 4.

表3 UV-A照射对亚洲玉米螟F1代卵孵化率、化蛹率、蛹羽化率和累积存活率的影响Table 3 Effects of UV-A radiation on the egg hatching rate, pupation rate, pupal eclosion rate and cumulative survival rate of F1 generation of Ostrinia furnacalis

表4 UV-A照射对亚洲玉米螟F1代幼虫和蛹发育历期的影响Table 4 Effects of UV-A radiation on the developmental duration of larvae and pupae of F1 generation of Ostrinia furnacalis
3 讨论
OfVg基因在雌成虫中表达量显著高于雄成虫及其他发育阶段,这与小菜蛾Plutellaxylostella和二化螟雌成虫Vg基因具有性别特异性的结论(邹明民, 2015; Huangetal., 2016)一致。雌成虫在羽化24 h时表达量达到最高,这可能和亚洲玉米螟雌成虫羽化8-24 h是其二级卵粒成熟期有关(钱仁贵, 1982),在此期间大部分的半成熟卵和透明卵室需要大量的Vg蛋白参与完成卵黄发生,形成成熟的卵粒,这导致了OfVg基因在雌成虫羽化24 h时高表达。雌成虫的不同组织中,脂肪体中的表达量最高,其他组织中仅少量表达,证实OfVg的表达具有组织特异性,这与邓瑶(2018)研究得出的Vg基因在雌成虫的脂肪体中高表达结果相同。亚洲玉米螟OfVg在不同发育阶段的差异表达和在雌成虫的脂肪体组织中特异表达,表明Vg在亚洲玉米螟生殖发育中发挥着重要作用。
紫外线是一种能对生物造成伤害的胁迫因子(Schauenetal., 2007)。在本研究中,亚洲玉米螟雌成虫在UV-A(320-400 nm)照射不同时长下OfVg表达量呈先缓慢下降后快速上升、再骤降的趋势。OfVg基因表达量在UV-A照射0.5 h和1 h后呈下降趋势,可能是由于UV-A对亚洲玉米螟造成了胁迫,为保护自身免受UV胁迫带来的损伤,不断增强其体内的抗逆能力和防御系统来提高自身的防御能力(王海鸿和雷忠仁, 2005; Mengetal., 2009),这必定会消耗机体大量的能量,从而导致OfVg基因表达量下降。在UV-A照射1.5, 2, 2.5, 3和3.5 h时,OfVg表达量逐渐快速升高,且在3.5 h时表达量达到最高(图5),表明亚洲玉米螟适应一段时间过后,UV-A照射可以诱导OfVg基因的表达,这与棉铃虫在UV-A胁迫后卵黄原蛋白基因的表达量显著增加结果(王智健和牛长缨, 2014)相同。UV-A照射4 h后OfVg的表达呈下降趋势,这可能是由于UV-A照射的时长超过了亚洲玉米螟的最大耐受性,对虫体造成了损伤,导致了OfVg基因表达量降低。在某些昆虫中,JH可介导Vg在脂肪体中的合成,如在赤拟谷盗中JH调节Vg合成(Parthasarathyetal., 2010)。张长禹等(2012)发现UV-A短时胁迫导致棉铃虫JH含量上调,长时胁迫导致JH含量降低。这与本研究结果一致。
Vg参与卵黄发生,卵黄发生能够直接影响昆虫的繁殖力(Amdanetal., 2006)。本研究结果表明,UV-A照射显著影响亚洲玉米螟的总产卵量和平均产卵量,随照射时间的延长先增多后减少,且在3 h/d时达到峰值(表2)。这与以上亚洲玉米螟OfVg表达量升高又下降的结果是相符的,表明一定时长UV-A照射可以促进生殖,该结果与棉铃虫H.armigera成虫在一定时间的UV-A照射后增强生殖力结果(Zhangetal., 2011)一致。亚洲玉米螟成虫在UV-A照射不同时长后,其F1代累积存活率和对照相比都有所降低(表3),且F1代幼虫的发育历期有所延长(表4)。这与东方粘虫Mythimnaseparata在UV-A照射后F1代累积存活率降低和F1代生长发育历期延长的结果(Alietal., 2016)相同。有研究显示昆虫能够延长其发育历期,以防止不利环境条件对它们生存造成影响(van Ooiketal., 2007)。亚洲玉米螟生殖力、F1代存活率和F1代发育历期均受UV-A照射影响,表明了UV-A是影响其种群发生的重要环境因子。
总而言之,本研究表明亚洲玉米螟OfVg主要存在于雌成虫的脂肪体中,参与其繁殖期卵的形成。UV-A照射一定时长会诱导OfVg表达量显著上调,促进生殖力,而UV-A照射超过一定时间,OfVg表达量受到抑制,其生殖力开始降低。本研究初步阐明了亚洲玉米螟OfVg在响应UV-A胁迫的作用,为深入研究UV-A胁迫对亚洲玉米螟发育和繁殖的影响奠定基础。
猜你喜欢
资源导刊(信息化测绘)(2022年4期)2022-11-24
河北科技师范学院学报(2022年2期)2022-08-26
今日农业(2021年14期)2021-11-25
安徽林业科技(2021年2期)2021-06-15
今日农业(2020年14期)2020-08-14
现代农业科技(2017年13期)2017-08-09
小溪流(画刊)(2017年6期)2017-07-10
农业科技与装备(2017年1期)2017-06-09
现代园艺(2016年7期)2017-01-09
现代园艺(2016年9期)2016-07-16
