
家蚕miR-2769靶基因的鉴定及表达分析
2020-05-22 13:27:54孙妍妍王景娅李晓哲阚云超乔惠丽
昆虫学报 2020年3期
孙妍妍, 王景娅, 王 露, 李晓哲, 阚云超, 乔惠丽,*
(1. 郑州大学生命科学学院, 郑州 450001; 2. 南阳师范学院, 河南省伏牛山昆虫生物学重点实验室, 河南南阳 473061)
MicroRNAs(miRNAs)是一种内源性小分子非编码RNA,长度约22 nt,广泛存在于原生动物、病毒、植物和动物中,且物种间具有高度保守性(Ambros, 2004; Bartel, 2004)。miRNAs通过与靶基因的非翻译区或编码区结合,在转录后或翻译水平上调节靶基因mRNA的降解或蛋白的表达,参与调控细胞增殖分化(Lietal., 2013)、凋亡(Heetal., 2013; Lietal., 2018)、自噬(Nelsonetal., 2014; Texadaetal., 2019; Zhaoetal., 2019)、免疫(Silwaletal., 2019; Xuetal., 2019)、发育和代谢(He and Hannon, 2004; Ameres and Zamore, 2013)等几乎所有的生命活动过程。lin-4和lin-7是最早在秀丽隐杆线虫Caenorhabditiselegans中发现的miRNAs(Leeetal., 1993);在哺乳动物中,约50%的编码蛋白基因表达受miRNAs的调控(Kroletal., 2010);miRNAs同样在昆虫中广泛存在,并在昆虫变态发育和生殖过程中发挥重要作用(Behura, 2007; Lucas and Raikhel, 2013; Liuetal., 2018, 2019)。
昆虫的变态发育由蜕皮激素20E和保幼激素(juvenile hormone, JH)协同调控,研究发现,20E不仅可抑制PI3K-Akt信号通路而促进自噬的起始,同时可与20E受体EcR-USP的异源二聚体结合,启动20E初级应答基因Br-C,E74,E75和E93的表达,进而通过与特异DNA序列结合启动晚期应答基因的表达(Riddifordetal., 2000; Yin and Thummel, 2005)。作为20E信号通路中的一个初级应答基因,细胞核激素受体E75(nuclear hormone receptor E75)基因最早在黑腹果蝇Drosophilamelanogaster中被鉴定,该基因在果蝇中通过5′端外显子的可变剪接编码DmE75A, DmE75B, DmE75C和DmE75D 4个异构体,其中DmE75A和DmE75C的DNA结合结构域含有2个锌指结构,DmE75B含有1个锌指结构,DmE75D没有DNA结合结构域(Feigletal., 1989; Segraves and Hogness, 1990; Bernardoetal., 2009)。在家蚕中,E75基因具有BmE75A,BmE75B和BmE75C3个可变剪接体,且其编码蛋白同样具有相同的C末端和不同的N末端(Segraves and Hogness, 1990)。对BmE75A,BmE75B和BmE75C在家蚕BmN细胞、前胸腺和脂肪体中的表达研究结果也表明,BmE75A,BmE75B和BmE75C具有不同的组织和时期特异性,而且三者的表达与20E的滴度密切相关(Lietal., 2015)。 近年研究表明,E75基因可参与不同昆虫幼虫到蛹的变态发育过程(Guoetal., 2016; Liuetal., 2018; Pengetal., 2019)。
家蚕Bombyxmori作为重要的经济昆虫,是研究昆虫变态发育分子机制的重要模式生物。家蚕的变态发育过程由20E和JH协同调控。在昆虫变态发育过程中需经历复杂的器官重建过程,脂肪体的自噬和凋亡对家蚕变态发育过程十分重要(Ihry and Bashirullah, 2014; Huetal., 2016)。本研究通过生物信息学方法分析预测家蚕miR-2769在20E处理后家蚕脂肪体中差异表达基因中的关联靶基因,利用双荧光素酶报告基因检测系统对miR-2769与其靶基因BmE75B的互作进行鉴定,进一步通过qRT-PCR检测miR-2769、靶基因BmE75B及其不同剪接体BmE75A和BmE75C在20E处理后家蚕脂肪体及不同发育时期和不同组织的表达特征,从而阐明miR-2769及其靶基因BmE75B在家蚕变态发育过程中的作用。
1 材料与方法
1.1 材料与试剂
供试家蚕为二化性的大造p50品系,常规条件下对蚕卵进行催青,幼虫用新鲜桑叶饲养。收集5龄第2天家蚕幼虫进行血淋巴注射蜕皮激素20E(2 μg/μL),每头家蚕注射5 μg,对照注射等体积无菌水稀释10倍的无水乙醇溶剂。注射后2, 6, 12和24 h 后分别取处理和对照的家蚕幼虫脂肪体。家蚕头、表皮、丝腺、脂肪体、精巢、卵巢、马氏管、中肠和血淋巴的组织取自正常家蚕5龄第3天幼虫;在4龄幼虫、5龄幼虫、蛹期至羽化后3 d成虫期间,每天分别随机取整虫,幼虫去中肠。每个样品3组,每组来自5头个体。所有样品液氮速冻后-80℃冰箱保存备用。
大肠杆菌EscherichiacoliTop10细胞,人胚胎肾细胞HEK293T细胞,psiCheck2载体为实验室保存,用于基因克隆的载体和试剂有pMD-19T载体(TaKaRa, 大连),Trizol Reagent(Invitrogen, 美国),RevertAid First Strand cDNA Synthesis Kit(Thermofisher,美国),限制性内切酶(Fermentas,美国),质粒DNA提取试剂盒和胶回收试剂盒(Axygen, 美国),Taq酶(Vazyme, 南京),T4 DNA连接酶(Thermofisher, 美国);用于细胞培养的试剂有DMEM高糖培养基(Hyclone, 美国),胎牛血清(BI, 以色列),细胞转染试剂为X-tremeGENE siRNA Transfection Reagent(Roche, 瑞士),双荧光素酶报告基因检测试剂盒(Promega,美国),qRT-PCR试剂为FastStart通用型SYBR® Green预混液(Roche, 瑞士)和All-in-oneTMmiRNA qRT-PCR检测试剂盒(Genecopia, 美国)。
1.2 家蚕总RNA的提取
将1.1节不同家蚕样品分别用液氮研磨,取约100 mg粉末加入1 mL Trizol试剂,按照RNA提取说明书进行抽提,RNA沉淀充分溶解后用超微量核酸蛋白测定仪(NanoDrop ND-2000)对总RNA的浓度和纯度进行测定。
1.3 家蚕miR-2769的序列和表达分析
根据前期测序获得的成熟miR-2769的序列(AUAUAUUAUCAGAUUUUCGGUC)设计上游引物(5′-ATATATTATCAGATTTTCGGTC-3′),以All-in-oneTMmiRNA qRT-PCR检测试剂盒中的通用的接头引物(Genecopia, 美国)为下游引物,对1.2节家蚕脂肪体RNA进行反转录,以反转录合成的cDNA为模板,利用All-in-oneTMmiRNA qRT-PCR检测试剂盒进行PCR扩增,扩增条件: 预变性95℃ 5 min; 95℃ 15 s, 55℃ 30 s, 72℃ 30 s,共30个循环;最后72℃延伸10 min。扩增产物纯化后连接pMD-19T载体,连接产物转化大肠杆菌Top10感受态细胞,挑取鉴定后的阳性克隆送深圳华大基因科技有限公司进行测序。
利用All-in-oneTMmiRNA qRT-PCR检测试剂盒对miR-2769在1.1节中20E处理组与对照组的2, 6, 12和24 h的脂肪体组织中的表达进行qRT-PCR检测。内参基因为家蚕5S rRNA(GenBank登录号: M35394.1),引物序列为5S-F: 5′-CGTCCGATCA CCGAAGTCAAG-3′;5S-R: 5′-AGGCGGTCACCCAT CCAAGTA-3′。扩增条件: 95℃ 5 min; 95℃ 15 s, 55℃ 30 s, 72℃ 30 s,共40次循环;最后72℃延伸10 min,溶解曲线范围为65℃~95℃。
1.4 家蚕miR-2769靶基因的预测及表达分析
课题组前期对20E处理前后的家蚕脂肪体进行了转录组测序,据此利用软件miRanda和生物信息学方法对20E处理前后家蚕脂肪体转录组数据中差异表达基因中的miR-2769靶基因进行预测。利用ClustalW2在线分析软件(https:∥www.ebi.ac.uk/Tools/msa/clustalw2/)对miR-2769靶基因BmE75B(GenBank登录号: NM_001112610.1)及其不同剪接体BmE75A(GenBank登录号: NM_001112609.1)和BmE75C(GenBank登录号: NM_001043577.1)的序列进行比对分析。
以1.2节提取的家蚕幼虫脂肪体RNA为模板,利用RevertAid First Strand cDNA Synthesis Kit反转录试剂盒合成cDNA,利用FastStart通用型SYBR® Green预混液对miR-2769靶基因BmE75B及其不同剪接体BmE75A和BmE75C进行qRT-PCR检测。扩增条件: 95℃ 5 min; 95℃ 15 s, 55℃ 30 s, 72℃ 30 s, 共40次循环;最后72℃延伸10 min,溶解曲线范围为65℃~95℃。BmE75B的引物序列为BmE75B-F: 5′-CACTCGGTGCTAGTGAGCAT-3′; BmE75B-R: 5′-TGCAGGGCCGGTATTGTATC-3′。BmE75A的引物序列为BmE75A-F: 5′-GGAACGTGAACCTGAA TTGC-3′; BmE75A-R: 5′-TGCAGGGCCGGTATTGT ATC-3′。BmE75C的引物序列为BmE75C-F: 5′-AGTGTTATCCGAAGTTGTCAC-3′; BmE75C-R: 5′-GAATGATGGAGGCTTGAGGA-3′。上述引物序列分别对应于BmE75B,BmE75A和BmE75C的N端特异序列。内参基因为家蚕BmactinA3(GenBank登录号: U49854),引物序列BmA3-F: 5′-ATGTGCGA CGAAGAAGTTGC-3′;BmA3-R: 5′-GTCTCCTACGTA CGAGTCCT-3′。
1.5 家蚕miR-2769的合成和BmE75B 3′UTR双荧光素酶报告载体构建
根据成熟miR-2769的序列及其与预测靶基因BmE75B的互作结合位点序列,由上海吉玛生物技术有限公司设计并合成miR-2769的野生型模拟物miR-2769 mimics(5′-AUAUAUUAUCAGAUUUUCGG UC-3′)、互作位点突变后的miR-2769 mutant(5′-AAUAUAAUUCAGAUUUUCGGUC-3′)及阴性对照miR-2769 NC(5′-UUCUCCGAACGUGUCACGUTT-3′)。
以RevertAid First Strand cDNA Synthesis Kit反转录试剂盒合成的家蚕脂肪体cDNA为模板,利用BmE75B3′UTR序列的特异引物,PCR扩增277 bp目的片段,纯化后扩增产物和psiCheck2载体分别用限制性内切酶XhoⅠ和NotⅠ进行双酶切,酶切产物切胶纯化后用T4 DNA连接酶连接,连接产物转化大肠杆菌Top10感受态细胞,经菌液PCR和双酶切鉴定后的psiCheck2/BmE75B3′UTR阳性克隆送深圳华大基因科技有限公司进行测序验证。引物序列为BmE75B3′UTR-XhoⅠ-F: 5′-AGCTCGAGGC CCCGACCTACTTTAAA-3′;BmE75B3′-UTR-NotⅠ-R: 5′-ATGCGGCCGCAATTCGACAACCTGTC-3′(下划线为酶切位点)。
1.6 家蚕miR-2769与靶基因BmE75B的互作分析
HEK293T细胞于37℃,5% CO2的条件下,用10%胎牛血清的DMEM高糖培养基培养。将生长状态良好的细胞提前1 d铺于24孔板中用于转染实验。将合成的miR-2769 mimics, miR-2769 mutant和miR-2769 NC分别与psiCheck2/BmE75B3′UTR共转染HEK293T细胞,psiCheck2/BmE75B3′UTR质粒单转为对照,根据X-tremeGENE siRNA转染试剂的说明书,优化后每个孔中质粒DNA的量为100 ng,miRNA的终浓度为0.1 μmol/L,每组转染3个重复,实验重复3次。转染后48 h收集细胞,根据双荧光素酶报告基因检测试剂盒说明书,对细胞进行裂解,利用化学发光仪依次检测海肾荧光素酶活性与萤火虫荧光素酶活性,二者的比值为萤火虫荧光素酶的相对活性(杨娟娟等, 2018)。
1.7 家蚕miR-2769与靶基因BmE75不同剪接体的表达分析
以1.2节提取的家蚕5龄幼虫不同组织、4龄幼虫到3日龄成虫整虫的RNA为模板,利用All-in-oneTMmiRNA qRT-PCR检测试剂盒进行反转录后对miR-2769进行qRT-PCR检测,内参基因为家蚕5S rRNA(方法同1.3节)。同时,利用RevertAid First Strand cDNA Synthesis Kit反转录试剂盒合成上述RNA的cDNA,利用FastStart通用型SYBR® Green预混液对BmE75B及其不同剪接体BmE75A和BmE75C进行qRT-PCR检测,内参基因为家蚕BmactinA3 (方法同1.4节)。
1.8 数据分析
双荧光素酶活性检测结果,根据萤火虫荧光素酶活性进行校正,以海肾荧光素酶活性与萤火虫荧光素酶活性的比值为萤火虫荧光素酶的相对活性,利用Prism 5.0作图软件进行作图和统计分析。qRT-PCR扩增结果用2-△△Ct法进行数据分析,针对不同内参基因的qRT-PCR扩增结果,先通过计算两个内参基因Ct值的几何平均数对不同内参基因的扩增进行校正,再用2-△△Ct法进行数据分析,最后用Prism 5.0软件进行作图和统计分析。
2 结果
2.1 家蚕miR-2769的序列和表达分析
以家蚕脂肪体组织RNA反转录后的cDNA为模板,PCR扩增miR-2769,扩增产物连接pMD-19T克隆载体,连接产物经初步鉴定后进行测序,测序结果表明扩增后miRNA序列与成熟miR-2769序列完全一致。利用qRT-PCR检测miR-2769在20E处理后2, 6, 12和24 h的家蚕5龄第2天幼虫脂肪体组织中的表达情况。结果如图1所示,与对照相比,miR-2769在20E处理后2, 6 和12 h表达显著下调(P<0.05),24 h时差异不显著(P>0.05)。
2.2 家蚕miR-2769靶基因的预测及表达分析
通过生物信息学预测,发现miR-2769的一个靶基因是家蚕的核激素受体基因BmE75的一个可变剪接体BmE75B。miR-2769第2-8位种子序列与预测靶基因BmE75B3′UTR存在互补配对(图2: A)。利用ClustalW2在线分析软件对BmE75B不同剪接体的氨基酸序列比对,分析结果表明,BmE75A, BmE75B和BmE75C的氨基酸序列具有不同的N端序列和完全相同的C末端(图2: B)。同时BmE75A,BmE75B和BmE75C的3′UTR区DNA序列也完全一致,因此BmE75A和BmE75C的3′UTR区也存在miR-2769的结合位点。

图1 qRT-PCR检测miR-2769在20E处理后不同时间家蚕幼虫脂肪体中的表达模式Fig. 1 Expression profiles of miR-2769 in larval fat body of Bombyx mori at different time after treatment with 20E by qRT-PCR处理组中往5龄第2天家蚕幼虫血淋巴中注射蜕皮激素20E(2 μg/μL),每头注射5 μg,对照组注射等体积无菌水稀释10倍的无水乙醇溶剂。图3同。In the treatment group, 2 μg/μL of molting hormone 20E (5 μg per individual) was injected into the hemolymph of the day-2 5th instar larvae, while in the control group (CK), an equal volume of ethanol in 10-fold dilution with distilled water was injected. The same for Fig.3. CK: 无水乙醇Ethanol. 图中数据为平均值±标准误,用双尾配对t检验进行统计分析。Data in the figure are mean±SE, and statistics analysis was performed by two tailed, paired t-test. *P<0.05; ns无显著差异No significant difference (P>0.05).
为了比较预测靶基因BmE75B及其剪接体与miR-2769在20E处理后家蚕幼虫脂肪体组织中的表达,利用qRT-PCR检测BmE75B在20E处理组与对照组在2, 6, 12 和24 h脂肪体组织中的表达。结果如图3(A)所示,与miR-2769的表达(图1)相反,BmE75B在20E处理后2, 6和12 h表达显著上调(P<0.001),24 h时表达差异不显著(P>0.05)。同时也检测了BmE75A和BmE75C在相同样品中的表达情况,结果如图3(B)和3(C)所示,与BmE75B类似,BmE75A在20E处理后2, 6和12 h家蚕幼虫脂肪体中表达也显著升高(P<0.001),24 h时表达差异不明显(P>0.05);BmE75C仅在20E处理后6 h时显著上调(P<0.01), 2, 12和24 h时与对照相比无显著差异(P>0.05)。由此可见,在脂肪体中miR-2769与BmE75A和BmE75B的表达存在负相关,推测在家蚕脂肪体中miR-2769可通过与靶基因BmE75 3′UTR的互作调控BmE75A和BmE75B的表达。
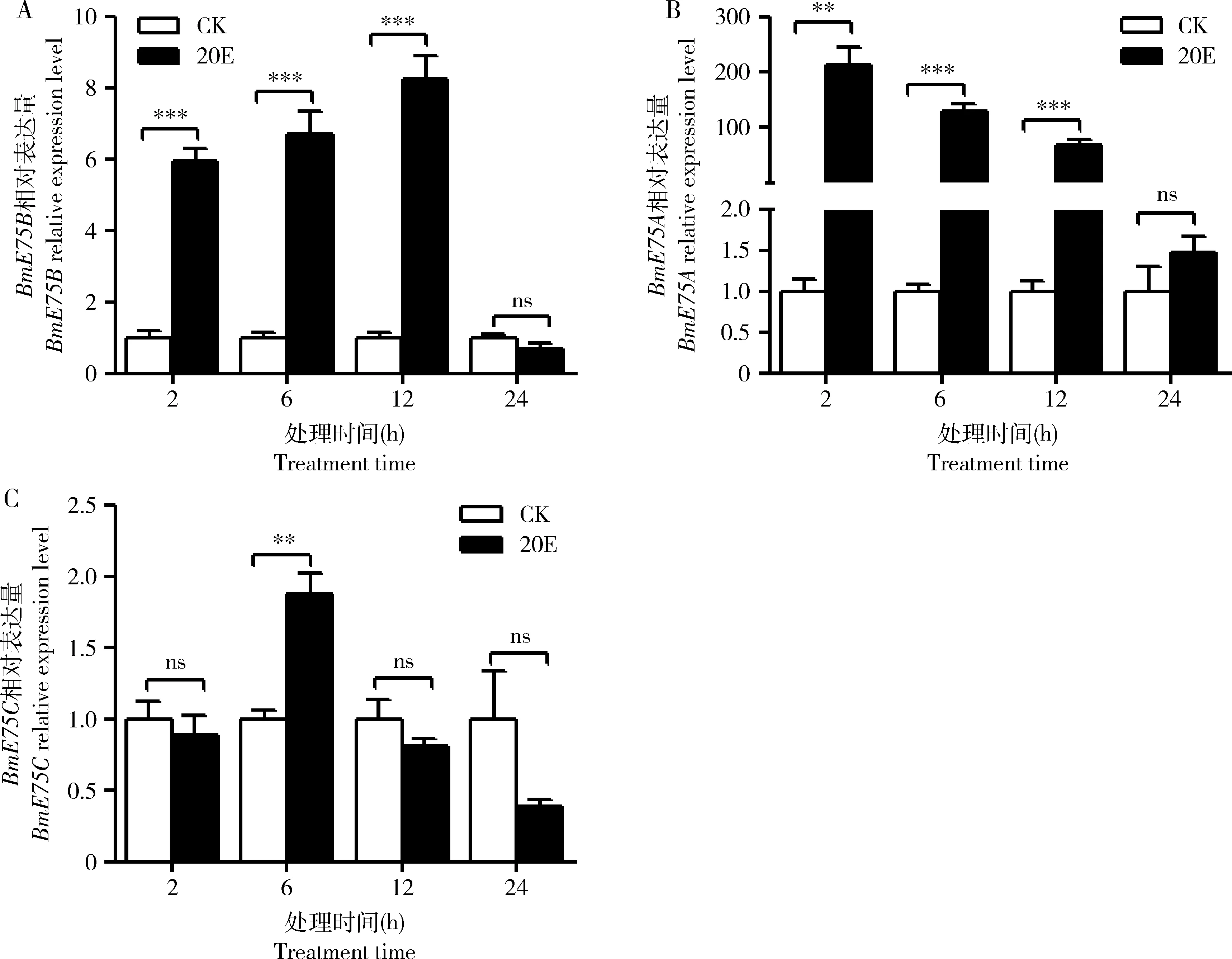
图3 qRT-PCR检测miR-2769靶基因BmE75不同剪接体在20E处理后不同时间家蚕幼虫脂肪体中的表达模式Fig. 3 Expression profiles of different splice isoforms of the target gene BmE75 of miR-2769 in larval fat body of Bombyx mori at different time after treatment with 20E by qRT-PCRA: BmE75B; B: BmE75A; C: BmE75C. 图中数据为平均值±标准误,用双尾配对t检验进行统计分析. Data in the figure are mean±SE, and statistics analysis was performed by two tailed, paired t-test. ***P<0.001; **P<0.01; ns无显著差异No significant difference (P>0.05).
2.3 双荧光素酶报告载体系统分析miR-2769与BmE75B的互作
BmE75B3′UTR序列的PCR扩增结果如图4(A)所示,扩增产物的分子量大小与理论一致。扩增产物连接psiCheck2双荧光素酶报告基因载体后的双酶切鉴定结果如图4(B)所示,酶切产物大小与理论分子量相符。构建好的psiCheck2/BmE75B3′UTR双荧光素酶报告载体经测序验证正确。
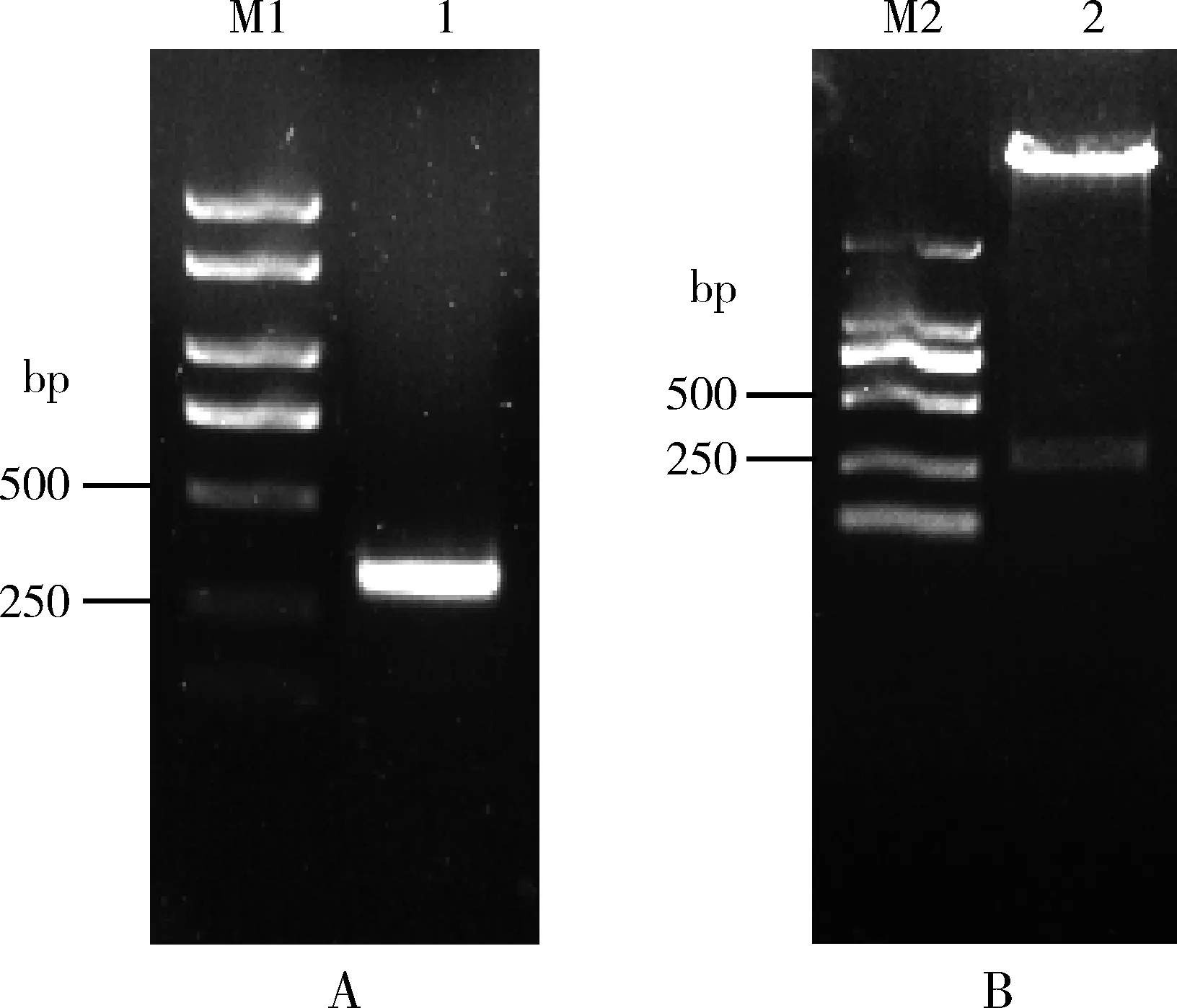
图4 BmE75B 3′UTR序列的PCR扩增(A)和双荧光素酶报告载体psiCheck2/BmE75B 3′UTR的酶切鉴定(B)Fig. 4 PCR amplification of BmE75B 3′UTR (A) and identification (B) of dual luciferase report vector of psiCheck2/BmE75B 3′UTR by restriction digestionM1: DL2000 Plus DNA Ladder; 1: BmE75B 3′UTR PCR扩增结果PCR product of BmE75B 3′UTR; M2: DL2000 DNA Ladder; 2: psiCheck2/BmE75B 3′UTR双酶切鉴定Identification of psiCheck2/BmE75B 3′UTR with double enzyme digestion.
将psiCheck2/BmE75B3′UTR双荧光素酶报告载体分别与合成的miR-2769 mimics, miR-2769 mutant和miR-2769 NC共转染HEK293T细胞,以psiCheck2/BmE75B3′UTR质粒单转为对照,利用双荧光素酶报告基因检测试剂盒检测转染后各组的海肾荧光素酶活性与萤火虫荧光素酶活性,二者的比值为萤火虫荧光素酶的相对活性。结果如图5所示,与psiCheck2/BmE75B3′UTR质粒单转染对照相比,只有psiCheck2/BmE75B3′UTR与miR-2769 mimics共转染时,报告基因的荧光素酶相对活性显著降低(P<0.05),而psiCheck2/BmE75B3′UTR与miR-2769 mutant或miR-2769 NC共转染后的酶活性与对照质粒无显著差异(P>0.05)。由此表明,miR-2769能够通过与靶基因BmE75B3′UTR区的互作负调控BmE75B在翻译水平的表达。由于BmE75A和BmE75C的3′UTR也具有miR-2769的作用位点,因此,推测miR-2769也可调控BmE75A和BmE75C的表达。
2.4 家蚕miR-2769与靶基因BmE75不同剪接体的表达分析
利用qRT-PCR进一步明确miR-2769及其靶基因BmE75不同剪接体在家蚕变态发育不同阶段的表达是否存在负相关,结果如图6所示,BmE75B在家蚕整个蛹期不同阶段的表达量相对较高,而miR-2769在所有的发育阶段的表达量都较低;BmE75A和BmE75C的表达特征也不同于BmE75B,BmE75A与miR-2769相似,在所有发育时期的表达量都较低,BmE75C则在5龄末和蛹早期具有极高的表达量;另由图中小图可见,虽然miR-2769与BmE75A的表达量较低,但二者的表达仍存在负相关。由此表明,miR-2769与BmE75A,BmE75B和BmE75C在家蚕变态发育的不同阶段均存在一定程度的表达负相关。
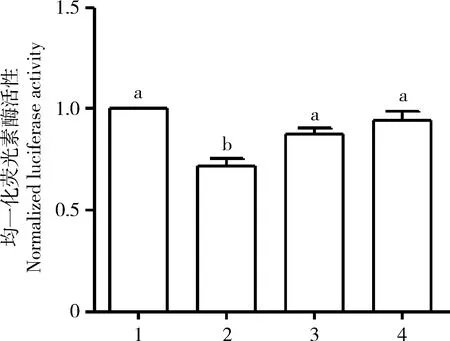
图5 psiCheck2/BmE75B 3′UTR质粒与miR-2769转染HEK293T细胞后荧光素酶活性分析Fig. 5 Analysis of luciferase activity in HEK293T cells after transfected with psiCheck2/BmE75B3′UTR and miR-27691: psiCheck2/BmE75B 3′UTR单转对照psiCheck2/BmE75B 3′UTR vector control; 2: psiCheck2/BmE75B 3′UTR+miR-2769 mimics; 3: psiCheck2/BmE75B 3′UTR+miR-2769 mutant; 4: psiCheck2/BmE75B 3′UTR+miR-2769 NC. 图中数据为平均值±标准误;柱上不同字母表示差异显著(P<0.05, 单因素方差分析, Tukey氏检验)。Data in the figure are mean±SE. Different letters above bars indicate significant difference (P<0.05, one-way ANOVA, Tukey’s test).

图6 qRT-PCR检测miR-2769及其靶基因BmE75不同剪接体在家蚕不同发育阶段的表达模式Fig. 6 Expression profiles of miR-2769 and different splice isoforms of its target gene BmE75 in different developmental stages of Bombyx mori by qRT-PCR 4L2D-5L8D: 分别为4龄第2天至5龄第8天2nd day of the 4th instar to the 8th day of the 5th instar, respectively; PP1-PP9: 分别为蛹期第1-9天1st-9th day of the pupal stage, respectively; A1-A3: 分别为成虫第1-3天1st-3rd day of the adult stage, respectively. 幼虫去中肠。图中数据为平均值±标准误;小图为miR-2769和BmE75A调整坐标轴刻度后的表达量。The midgut of larvae was removed. Data in the figure are mean±SE. Inset shows the expression of miR-2769 and E75A in adjusted Y axis scale.
为了进一步分析miR-2769与其靶基因BmE75不同剪接体在家蚕个体中的表达情况,首先以家蚕5龄第3天幼虫不同组织的cDNA为模板,利用qRT-PCR分别对miR-2769和BmE75不同剪接体进行检测。结果表明,miR-2769在5龄幼虫的精巢和卵巢中的相对表达量较高,其他组织的表达量相对较少;靶基因BmE75B在精巢、卵巢和脂肪体中的表达量相对较高,除头部、表皮、血淋巴和马氏管中表达量无显著性差异(P>0.05)外,其他组织中miR-2769与BmE75B的表达差异均为显著(P<0.05)。而BmE75A和BmE75C具有不同于BmE75B的表达特征,BmE75A在血淋巴和卵巢中表达量较高,而BmE75C的整体表达量都不高,但miR-2769与BmE75A和BmE75C在多个不同组织中的表达也存在显著差异(P<0.05)(图7)。

图7 qRT-PCR检测miR-2769及其靶基因BmE75不同剪接体在家蚕5龄幼虫不同组织中的表达模式Fig. 7 Expression profiles of miR-2769 and different splice isoforms of its target gene BmE75 in different tissues of the 5th instar larvae of Bombyx mori by qRT-PCR1: 头部Head; 2: 表皮Epidermis; 3: 血淋巴Hemolymph; 4: 中肠Midgut; 5: 马氏管Malpighian tubules; 6: 脂肪体Fat body; 7: 丝腺Silk gland; 8: 精巢Testis; 9: 卵巢Ovary. 图中数据为平均值±标准误;柱上不同字母表示差异显著(P<0.05, 单因素方差分析)。Data in the figure are mean±SE, and different letters above bars indicate significant difference (P<0.05, one-way ANOVA).
3 讨论
miRNAs是一类小分子量的非编码RNA,主要通过与靶基因的互作调控mRNA的降解和翻译抑制。本研究发现家蚕miR-2769在20E诱导早期的脂肪体中表达下调(图1),BmE75B作为miR-2769的预测靶基因,在20E诱导后家蚕脂肪体中的表达与miR-2769的表达相反。同时20E在家蚕脂肪体中可快速诱导BmE75A的高表达,而BmE75C在20E诱导后表达升高较慢,且表达水平相对较低(图3)。利用NCBI数据中的Blast比对分析表明,家蚕的BmE75A,BmE75B和BmE75C与黑腹果蝇DmE75A,DmE75B和DmE75C编码的氨基酸序列一致性约为50%。研究表明,黑腹果蝇DmE75A和DmE75B被20E诱导后快速高表达,DmE75C诱导后表达则较慢且表达量弱的结果一致(Bialeckietal., 2002; Bernardoetal., 2009),由此可见,E75不同剪接体在20E诱导后家蚕和果蝇中具有相似的表达特征。同时,前人也研究表明,20E也可诱导家蚕BmE75基因的高表达(Matsuoka and Fujiwara, 2000; Sweversetal., 2002; Shiraietal., 2012),且BmE75A,BmE75B和BmE75C对20E具有发育阶段和组织的特异表达特征(Lietal., 2015)。
利用双荧光素酶报告系统研究miR-2769与靶基因的互作,与家蚕中很多miRNAs调控靶基因的方式类似(Huangetal., 2012; Jiangetal., 2013; Lingetal., 2015; Liuetal., 2018, 2019),miR-2769能够通过与靶基因BmE75B3′UTR区的序列互作调控靶基因在翻译水平的表达,由于BmE75A,BmE75B和BmE75C的3′UTR具有相同的miR-2769互作位点,推测miR-2769也可调控BmE75A和BmE75C的表达。结果表明在家蚕幼虫脂肪体中,BmE75A和BmE75B在20E诱导家蚕脂肪体后不同时间的表达量均与miR-2769的表达具有负相关;在正常家蚕5龄幼虫的大部分组织中,miR-2769与BmE75B不同剪接体的表达也都存在负相关,由此表明,miR-2769可能通过与BmE75 3′UTR的互作调控其不同剪接体在不同组织中的表达。
不同发育阶段和组织中的表达特征表明,miR-2769与BmE75的不同剪接体在4龄第2天到成虫不同发育阶段和5龄幼虫不同组织中的表达均具有特异性表达特征。在血淋巴中miR-2769可促进BmE75A的表达,在脂肪体中miR-2769可促进BmE75B的表达,而在其他大部分组织中miR-2769抑制BmE75不同剪接体的表达(图7),推测可能与家蚕不同组织中20E的滴度变化有关。由于家蚕4龄幼虫到成虫为整虫取样,整虫样品中miR-2769的模板量可能被稀释,使miR-2769在家蚕不同发育时期的表达量整体较低,而BmE75A,BmE75B和BmE75C在不同阶段的表达量则各不相同,尤其BmE75C在5龄末和蛹初期表达量极高;BmE75B在5龄末、蛹期第1天、蛹期第4天,蛹期第7天和羽化前的变态发育时期表达量较高(图6),该结果与家蚕变态发育过程中20E的滴度变化(Burdette, 1962)基本一致;BmE75A的表达量则整体较低,但大部分时间点miR-2769与BmE75不同剪接体的表达均为负相关。在烟草天蛾Manducasexta幼虫蜕皮和化蛹过程中,检测到E75A和E75B的表达与20E水平的升高一致,但E75A的表达稍早于E75B(Zhouetal., 1998)。由于20E在家蚕不同发育阶段及不同组织的滴度不同,可能使miR-2769与BmE75的不同剪接体在不同组织和不同发育时期表现出不同的调控模式。另外,miR-2769和BmE75B在精巢和卵巢中的表达量非常高,而BmE75A在卵巢中的表达量与miR-2769相当(图7),而非负相关。在家蚕、烟草天蛾M.sexta和帕眼蝶Parargeaegeria中卵巢中特异表达的miR-2763和miR-989,在卵巢的发育中发挥作用(Jagadeeswaranetal., 2010; Zhangetal., 2012; Quahetal., 2015),家蚕miR-2769是否参与卵巢的发育还有待进一步证实。此外,在雌性果蝇中,DmE75A和DmE75B在卵室发育和凋亡过程中发挥相反的作用,DmE75A可诱导哺育细胞的凋亡,DmE75B可促进卵的发育(Terashima and Bownes, 2006),二者在家蚕卵发育中是否具有相似的功能还未见报道。最近,Liu等(2018)发现miR-14也可通过与靶基因E75和ECR-B3′UTR的互作参与家蚕幼虫的发育过程,由此推测家蚕BmE75基因可能还存在除miR-2769外的其他调控因子,参与其可变剪接和功能调控。
综上所述,miR-2769作为家蚕20E信号通路中关键基因BmE75新的调控因子,可通过与靶基因BmE75 3′UTR区的互作对BmE75不同剪接体进行负调控,同时miR-2769与靶基因BmE75不同剪接体在不同发育时期和不同组织中具有不同的表达特征。该研究不仅丰富了miRNAs参与家蚕变态发育调控的功能研究,同时也为家蚕BmE75不同剪接体的调控机制及其在昆虫变态发育过程中的功能研究提供了新的依据。
猜你喜欢
四川蚕业(2022年2期)2022-11-19 02:09:52
天津医科大学学报(2021年4期)2021-08-21 02:14:50
中国科学探险(2021年2期)2021-06-01 03:03:56
中日友好医院学报(2021年1期)2021-04-14 01:58:32
四川蚕业(2021年2期)2021-03-09 03:15:30
四川蚕业(2021年1期)2021-02-12 02:03:20
南方农业学报(2020年8期)2020-11-02 02:41:06
山东医药(2020年9期)2020-05-20 01:12:16
生物学教学(2016年2期)2016-04-10 07:30:00
蚕桑通报(2015年1期)2015-12-23 10:14:32
