
红火蚁入侵强度对本地蚂蚁群落物种的发现-支配权衡的影响
2020-05-22 13:28:26卢志兴于潇雨张念念陈又清
昆虫学报 2020年3期
燕 迪, 卢志兴, 王 庆, 于潇雨, 张念念, 李 巧, 陈又清,*
(1. 西南林业大学生物多样性保护学院, 昆明 650224; 2. 中国林业科学研究院资源昆虫研究所, 昆明 650224;3. 贵州省林业科学院, 贵阳 550011)
蚂蚁入侵是世界各地生物多样性尤其是本地蚂蚁的主要威胁因素(Holwayetal., 2002)。往往在这些入侵种侵入的地区,本土种蚂蚁的多度减少了90%以上(Porter and Savignano, 1990)。红火蚁Solenopsisinvicta隶属于膜翅目(Hymenoptera)蚁科(Formicidae)切叶蚁亚科(Myrmicinae),被列为世界百大入侵物种之一(GISD, 2019),其强大的适应性和极其快速的繁殖使得红火蚁几乎能在地球上任何地方扩散和繁殖(Bertelsmeieretal., 2017; Lyu and Lee, 2017)。
在蚂蚁入侵机制方面,大量研究揭示了入侵蚂蚁和本土蚂蚁在觅食能力、温度耐受性及攻击性等特性之间存在差异(Holwayetal., 2002; 许益镌等, 2009; Chenetal., 2014; 雷妍圆等, 2016)。事实上,由于蚁群的社会化和模块化的性质缓冲了它们被天敌捕食以及来自环境的压力(Andersen, 1991; Davidson, 1998),而蚂蚁不同种之间具有高度的互动性(Kingetal., 2009),竞争成为蚂蚁群落中物种间相互作用最重要的特征(Davidson, 1998)。蚂蚁群落中物种间的竞争主要体现在干扰竞争和资源利用竞争这两个方面。干扰竞争是物种通过直接侵略或者通过维持领土间接阻止其他种类获取资源,而资源利用竞争则是物种具有先于其他物种发现和利用资源的能力(Fellers, 1987)。
在原生群落(长期稳定、未受入侵)中,不同蚂蚁物种之间通过干扰竞争或资源利用竞争实现共存,被称为发现-支配权衡(discovery-dominance trade-off)(Adleretal., 2007)。原生蚂蚁群落的不同物种在发现资源的能力与支配资源的能力上能达到平衡(Fellers, 1987; Lebrun and Feener, 2007; Parr and Gibb, 2012)。在红火蚁的原生群落中,红火蚁的发现能力较弱(相较于其他蚂蚁种类而言),但支配能力较强,达到了一个平衡,因此原生群落的蚂蚁能够共存(Calcaterraetal., 2008)。
入侵蚂蚁在进入新蚂蚁群落后,可能不仅在其中一个方面表现出色,而是两者均表现出更强的能力。阿根廷蚁Linepithemahumile已被证实打破了美国西部蚂蚁群落的发现-支配权衡,即该蚂蚁比本土蚂蚁具有更强的发现能力和支配能力(Holway, 1999),这一特征为入侵蚂蚁带来了极大的竞争优势,使得本土蚂蚁逐渐被淘汰,这一优势揭示了一种直接的入侵机制(Human and Gordon, 1996)。入侵种蚂蚁更强的侵略性和干扰能力有助于其保护和垄断资源(Del Toroetal., 2012; Calcaterraetal., 2016),这些都为入侵蚂蚁争夺生态位并逐渐取代本土蚂蚁提供了重要条件(Wilderetal., 2013; Wittman, 2014)。
红火蚁入侵中国已久,其庞大的种群数量在短时间内就可以对本地蚂蚁群落造成破坏(Hölldobler and Wilson, 1990; Wangetal., 2019),但在红火蚁入侵早期,其种群数量尚低时是否同样具有强大的竞争力鲜有报道。同时,不同种群数量的红火蚁对本地蚂蚁群落物种的发现-支配权衡具有怎样的影响也不得而知。本研究采用陷阱法和诱饵法,通过量化入侵区域和未入侵区域中不同蚂蚁种类的发现能力和支配能力,探讨: (1)红火蚁入侵是否会打破本地蚂蚁群落物种的发现-支配权衡; (2)这种权衡是否会随红火蚁入侵强度的强弱而不同; (3)不同入侵强度区域的红火蚁在发现能力和支配能力上是否具有差异。
1 材料与方法
1.1 研究地概况
野外实验于2017年10-11月在云南省楚雄州牟定县的滇橄榄Phyllanthusemblica种植基地(25°38′36″N,101°45′03″E)进行。基地总面积约33 hm2,地处云南省中北部,楚雄州中部,海拔约1 625 m,亚热带季风气候,平均年降雨量约872 mm,平均年气温为15.8℃,一年中无霜期238 d,全年日照时数2 359 h,日照充足,气候温和。样地内为滇橄榄单一种植模式,地面生长着较为稀疏的杂草,杂草盖度为30%。入侵区域内红火蚁生殖类型均为多蚁后型,其中,强入侵区(strong invasion plot)活动蚁巢密度为5.6个/100 m2,按照《红火蚁疫情监测规程》标准对红火蚁发生级别的划分(根据单位面积活动蚁丘数量划分)(陆永跃和曾玲, 2015),红火蚁发生等级属于4级危害区,即重度危害区。弱入侵区(weak invasion plot)活动蚁巢密度为0.14个/100 m2,属于2级危害区,即中度危害区。为了去除边缘效应,保证对照区的蚂蚁种类和入侵区的蚂蚁种类差异不大,我们将没有观测到红火蚁蚁巢的对照区(control plot)设置在与入侵区间隔100 m的邻近区域,以尽可能消除生境不同带来的差异。
1.2 蚂蚁诱集和种类选取
参考McGlynn(2000)的方法,用金枪鱼罐头(山东蓝润水产有限公司)作为诱饵。将标准培养皿(直径9 cm)沿着底部去除边壁制成诱饵平台,将约1 g的金枪鱼肉放于培养皿中央,由于金枪鱼罐头含有油脂,能更好地将气味发散出去,用于诱饵试验效果较好。试验时选取晴好天气进行,气温20~30℃,时间在上午10∶00-12∶00时和下午14∶00-17∶00时进行。强入侵区、弱入侵区和对照区的总面积分别为9.73, 8.62和9.03 hm2,在每个样地中设置3条样带,样带间间隔大于50 m。每条样带选取6个样点,试验时在每个样点上摆放4个诱饵,诱饵间隔2 m,以尽可能让更多的蚂蚁发现诱饵并过来取食。在诱饵设置完成后的5, 10, 15, 30, 60和120 min时检查每个样点的蚂蚁种类和数量并拍照记录,若前来的同一种蚂蚁数量超过5头,使用镊子快速抓取其中1头放入装有75%的酒精中保存,留待鉴定。记录每个诱饵上发现的第一个蚂蚁物种,蚂蚁个体数量按照1, 2, 5, 10, 20, 50, 100, 200和300数量级估计(Morrison, 2002)。在所有诱饵试验完成之后,在离诱饵相近的位置设置陷阱,陷阱的规格为直径6 cm、高度7 cm的塑料杯,使用50 mL 50%的乙二醇溶液作为保存液,杯口与地面平齐,放置48 h后,收集陷阱中蚂蚁标本带回实验室进行标本鉴定并统计其数量,通过形态特征将采集到的蚂蚁鉴定到种。陷阱法可以提供蚂蚁群落的总体分布情况,而诱饵法与陷阱法相结合可以消除蚂蚁多度的影响,从而更准确地反映蚂蚁的觅食活动(Andersen, 1991)。本试验通过陷阱法采集蚂蚁28种2 944头,我们选取了陷阱中出现频率较高的7种蚂蚁进行数据分析,分别是红火蚁Solenopsisinvicta(34.71%)、巴瑞弓背蚁Camponotusparius(5.13%)、扁平虹臭蚁Iridomyrmexanceps(33.63%)、中华小家蚁Monomoriumchinense(5.74%)、伊大头蚁Pheidoleyeensis(5.26%)、拉帕铺道蚁Tetramoriumlaparum(6.59%)和红头弓背蚁Camponotussingularis(5.95%)。出现频率较低的蚂蚁物种在本研究中的意义并不高,因此没有选取。
1.3 行为指数的定义
工蚁(包括兵蚁)的“相对发现能力”利用一个物种在诱饵中出现的数量和在陷阱中出现的数量的比例来量化,所有蚂蚁物种在诱饵上出现的总次数除以所有蚂蚁物种在陷阱中记录的总次数,所得到的比例用于标准曲线(零模型)的建立(Lebrun and Feener, 2007)。假设区域内所有的蚂蚁物种都是同样优秀的探索者,那么任何蚂蚁物种都会符合该预测模型,即完全分布在预测直线上,用蚂蚁实际出现在诱饵上的数量与预测值之间的差值(发现残差,RD)来定义蚂蚁物种的相对发现能力,这样可以消除不同种类在环境中由于多度不同造成的误差(特别是红火蚁这一种群数量极大的物种)。
蚂蚁发现诱饵后为了支配(垄断)资源会招募同伴,招募能力的强弱用临界数量(Ncrit)来判断,Ncrit值的选取通常取决于蚂蚁物种的大小和行为,对于种群数量较大的物种定义为10头,而种群较小的定义为5头(如猛蚁类)(Santinietal., 2007)。在本研究中,Ncrit值定为5用于解释招募能力弱的物种。在每个观察时间段内记录蚂蚁的种类和数量,当某种蚂蚁的数量超过5头时认为该蚂蚁开始了招募行为。 我们并非进行连续观察,因此无法得知是否出现多次招募现象,参考前人的方法(Calcaterraetal., 2008),本研究中也默认所有发生招募的蚂蚁种类都只进行了一次招募,我们用每个时间段内观察到的蚂蚁数量来表示招募的动态变化。
通常采用行为优势来代表对资源的掌握或支配能力(behavioral dominance),即A蚂蚁物种通过打斗等行为保护了它们支配的资源或被B蚂蚁物种驱赶从而B物种支配了该资源(Lebrun and Feener, 2007; Feeneretal., 2008)。但对于一个蚂蚁种群,只有完全支配资源才能为蚁巢内的大部分群体提供更多的营养,其过程可以不考虑(Parr and Gibb, 2012)。在本研究中,我们观察到的物种之间相互作用很少发生,因此采用行为优势来代表支配能力是不合适的,所以我们参考Parr和Gibb (2012)的方法,用支配比例来定义支配能力。支配意味着这种蚂蚁完全占有,判定的依据是诱饵平台中仅有一种蚂蚁存在且其数量至少为5头。在120 min后,记录了每个物种支配的诱饵数量,由于入侵区红火蚁的种群数量太大而导致了绝大部分诱饵被吃完,因此入侵区内我们统计的是60 min时的垄断数量。为消除多度的影响,以每种蚂蚁支配的诱饵数量和该物种发现的诱饵数量的比值来代表相对支配能力。
为了进一步分析不同强度的入侵区中红火蚁竞争能力的差异,我们还统计了红火蚁首次发现食物时间段的频次、发生招募现象的频次以及发生支配现象的频次。
1.4 数据分析
对红火蚁3种入侵强度的区域中所有蚂蚁物种的相对发现能力进行量化,物种偏离直线的程度代表它相对于竞争对手的发现能力,与其多度无关。位于虚线上方的物种在发现能力上比预期更强,而下面的物种则更弱,偏离程度越大,表明与预期差异越大。
3个样地类型中蚂蚁物种的相对发现能力的预测曲线通过协方差分析来检验它们之间是否平行(即是否具有差异)。
我们将代表发现能力的残差值(RD)和代表支配能力的支配指数的数据进行排序(rank),将发现能力最强的蚂蚁物种标记为1,然后依次进行标记;同样,将支配能力最强的蚂蚁物种标记为1,依次进行标记。使用Spearman秩相关性检验蚂蚁物种发现能力和支配能力的秩相关性。未观察到垄断现象的物种用最高等级标记,若存在多个物种则标记为相同等级(Parr and Gibb, 2012; Bertelsmeieretal., 2015)。
对红火蚁在强入侵区和弱入侵区的发现频率、招募频率和支配行为频率进行卡方检验,当卡方检验的最低期望频率低于5时。采用Fisher精确检验校正P值。
所有分析均在R语言中进行(version 3.6.1)。
2 结果
2.1 蚂蚁群落物种的发现-支配权衡
3种红火蚁入侵强度区域的预测曲线如图1,在强入侵区中,预期发现能力=0.50×实际陷阱中出现频率;在弱入侵区中,预期发现能力=0.41×实际陷阱中出现频率;而在对照区中,预期发现能力=0.43×实际陷阱中出现频率。这3种红火蚁入侵区的预测曲线没有显著差异(F=2.70,P=0.77)。
用残差值代表“发现指数”,将得到的每种蚂蚁种类的残差值进行排序,即可得到相应的排名(表1)。
同样,我们统计了3个区域的相对支配能力,该能力用某种蚂蚁支配(没有其他种类蚂蚁存在于诱饵附近)的诱饵数量和该物种出现的诱饵数量的比值得到,其数值分布在0-1的范围内,若值为1,表明该物种完全占据了它们发现的所有诱饵;若值为0,表明该物种在一定时间后被其他种赶走,迫使其无法继续获得诱饵平台的食物资源(表2)。

图1 红火蚁3种入侵强度区域中蚂蚁群落物种的相对发现能力“预测曲线”Fig. 1 The relative discovery ability “expected curve” of species of ant community in areas ofSolenopsis invicta with three invasion intensitiesA: 强入侵区(5.6个活动蚁巢/100 m2) Strong invasion plot (5.6 ant colonies/100 m2); B: 弱入侵区(0.14个活动蚁巢/100 m2) Weak invasion plot (0.14 ant colonies/100 m2); C: 对照区(无蚁巢)Control plot (without ant colonies). Si: 红火蚁S. invicta; Cp: 巴瑞弓背蚁Camponotus parius; Ia: 扁平虹臭蚁Iridomyrmex anceps; Mc: 中华小家蚁Monomorium chinense; Py: 伊大头蚁Pheidole yeensis; Tl: 拉帕铺道蚁Tetramorium laparum; Cs: 红头弓背蚁Camponotus singularis. 虚线表示根据“零模型”得到的预测曲线。The dashed line indicates the expected curve obtained from the “zero model”.
强入侵区Strong invasion plot: 5.6个活动蚁巢/100 m2(5.6 ant colonies/100 m2); 弱入侵区Weak invasion plot: 0.14个活动蚁巢/100 m2(0.14 ant colonies/100 m2); 对照区Control plot: 无蚁巢Without ant colonies. 下同The same below. 残差值(RD)量化不同蚂蚁种类的相对发现能力,正值表示高于预测值,负值表示低于预测值。The residual value (RD) quantifies the relative discovery ability of different ant species, positive values indicate higher than the predicted values and negative values indicate lower than the predicted values.
根据表中的排名,我们对3个区域内蚂蚁群落不同物种的相对发现能力和相对支配能力进行了秩相关性分析,结果如图2所示。

表2 红火蚁3种入侵强度的区域中蚂蚁的相对支配能力Table 2 Relative dominance ability of ant species in areas of the red imported fire ant Solenopsis invictawith three invasion intensities
支配能力用该种蚂蚁以每种蚂蚁支配的诱饵数量和该物种发现的诱饵数量的比值表示,越接近最大值1表示其能力越高,0表示完全不能支配。The dominance ability is expressed by the ratio of the monopoly frequency and the frequency of appearance of the species in the baits. The ability gradually increases as the value approaches the maximum value 1. The value 0 means that it is completely uncontrollable.

图2 红火蚁3种入侵强度区域中蚂蚁群落物种的发现-支配权衡Fig. 2 The discovery-dominance trade-off of species of ant communities in areas of the red imported fire ant Solenopsis invicta with three invasion intensitiesA: 强入侵区Strong invasion plot; B: 弱入侵区Weak invasion plot; C: 对照区Control plot. 箭头位置表示红火蚁(Si)所对应的点;星号表示有发现指数与支配指数有显著相关性(P<0.05),负值表示负相关。The position indicated by the arrow indicates the point corresponding to S. invicta (Si). Asterisk indicates that there is a significant correlation between the discovery index and the dominance index (P<0.05), and the negative value indicates negative correlation.
在强入侵区中,蚂蚁物种的相对发现能力和相对支配能力几乎不存在相关性(ρ=-0.01,P=0.97)。在弱入侵区,因为红火蚁种群数量的减少,蚂蚁物种的相对发现能力和相对支配能力存在一定的负相关性,但是依然不显著(ρ=-0.60,P=0.21)。在红火蚁入侵区,蚂蚁物种不存在或者不遵循发现-支配权衡。但在没有红火蚁的对照区,这些本地种蚂蚁的相对发现能力和相对支配能力存在着显著地负相关性(ρ=-0.81,P=0.04),遵循发现-支配权衡。
2.2 不同强度入侵区内红火蚁竞争能力的差异
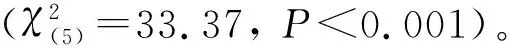

表3 2种入侵强度区域中红火蚁首次发现诱饵的时间段的频率Table 3 Frequencies of the earliest time period of discovering baits by the red imported fire antSolenopsis invicta in areas with two invasion intensities

表4 2种入侵强度区域中红火蚁发现、招募和支配行为频率Table 4 Frequencies of discovery behavior, recruitment behavior and dominance behavior of the red imported fire ants Solenopsis invicta in areas with two invasive intensities
括号中的值表示未发生该行为的频率。The values in brackets indicate the frequency without this behavior.
3 结论与讨论
IUCN目前已列出了19种入侵性蚂蚁(IUCN ISSC入侵物种专家组, 2018)及其对生物多样性、农业、人类健康和经济的巨大影响,迫切需要更好地了解入侵的机制。入侵物种往往在二元竞争试验中表现出强大的能力,战胜并杀死大多数的本土种蚂蚁(Holway, 1999; Bertelsmeieretal., 2015; 雷妍圆等, 2016),这表明入侵种蚂蚁相较于本地蚂蚁物种来说是竞争优势种。但蚂蚁群落往往并非是两个物种进行竞争,而是存在着多个物种。
多数研究表明,在蚂蚁群落中物种之间存在着线性的支配优势,这使得竞争优势种经常可以占领从属种的食物或巢穴(Human and Gordon, 1996; Del Toroetal., 2012; Calcaterraetal., 2016)。这在入侵种成功侵入的地区很常见,例如红火蚁。在未被入侵的本地蚂蚁群落中,不同物种在发现资源和保护资源的能力上有一个充分的权衡(Adleretal., 2007),就像本研究中的对照区(图2: C),这与前人的研究一致,在红火蚁的原生群落中同样如此(Calcaterraetal., 2008; Feeneretal., 2008)。
在一些蚂蚁群落中物种间竞争的研究中,红火蚁发现的诱饵数量是要低于预测值的(Lebrun and Feener, 2007; Calcaterraetal., 2008),本研究中,根据“零模型”预测曲线,无论入侵强度的高低,红火蚁发现的诱饵数量总是高于预期值,造成此差异的原因可能主要是因为寄生天敌的缺失引起的,在红火蚁的原产区,正是由于竞争优势种存在着寄生蜂等天敌,其所在的蚂蚁群落才得以保持着“平衡”(Calcaterraetal., 2008)。而在我们的研究中,红火蚁是一个外来入侵种,缺乏原产地的天敌的威胁,因此其发现能力表现出偏高的水平。蚂蚁群落中物种间竞争往往是不对称竞争,获胜者往往是数量占优势的一方(Hölldobler and Wilson, 1990),这也解释了在弱入侵区中红火蚁的相对支配能力要低于强入侵区。在相对发现能力上得分较高的物种在相对支配能力上得分较低,表明在没有红火蚁入侵的情况下,本地蚂蚁群落中物种间的竞争是遵循发现-支配权衡规律的。无论红火蚁强度的高低,本地蚂蚁群落中存在的发现-支配权衡确实被打破了,甚至红头弓背蚁这一物种似乎被淘汰了(无法出现在任何一个诱饵及陷阱之中),强入侵区的权衡完全被破坏,而弱入侵区受到的影响相对较小。有一点值得注意的是,强入侵区的红火蚁在发现能力和支配能力上表现出共同的优势,即都领先于其他物种;然而弱入侵区的红火蚁表现出遵循发现-支配权衡的规则(发现能力与支配能力正好相反),但其他蚂蚁出现了部分偏移。出现这种结果的原因可能是,在强入侵区,由于红火蚁的种群数量太过庞大,蚁群的活动范围覆盖了绝大多数的诱饵(Holwayetal., 2002; Wittman, 2014);而在弱入侵区,红火蚁种群数量相对较少(基数差距过大),同其他蚂蚁的竞争主要在于其支配能力强,能够迫使其他蚂蚁放弃对食物资源的争夺。在有些蚂蚁种群中(包括红火蚁),存在着一部分蚂蚁个体作为搜索者(scouts),这部分个体主要的作用是外出搜寻食物资源并提醒巢穴中其他不活跃个体(Gordon, 1983),其他个体则主要集中在蚁巢中等待着被招募,该部分个体被称为应征者(recruits)。如何对搜索者和应征者进行兵力投入定义了该种群的觅食策略(van Oudenhoveetal., 2018)。种群对搜索者投入的兵力越多,它就越能够有效地发现食物,相反,对应征者投入更多意味着该种群将更有效地收获资源(取决于食物资源的规模),而这将转化为干扰竞争的优势(Adleretal., 2007)。蚁群往往可以根据内部和外部环境调整任务的分配(Gordon, 1996),能够进行集体招募的种群更可以有效地将任务分配与环境相匹配(Lanan, 2014)。
Oudenhove等(2018)的研究发现,蚂蚁种群在有干扰竞争的群落中,在食物资源稀缺的环境下投入更多的搜索者,在食物资源丰富的情况下投入更多的应征者。在我们的研究地中,蚂蚁的食物资源相对较少。本研究在提供额外食物资源的情况下发现:强入侵区的红火蚁发现了更多的食物资源且发现时间明显要更快(表3和表4),该结果表明强入侵区的红火蚁种群比弱入侵区的种群投入了更多的搜索者,但无法解释红火蚁在搜索者和应征者的分配上是否不同。我们在比较红火蚁的招募频率时,强入侵区和弱入侵区不存在显著的差异(表4),这在一定程度上表明强入侵区和弱入侵区投入的应征者的相对兵力基本相同。强入侵区的红火蚁种群在支配能力上显著强于弱入侵区(表4),这说明尽管相对兵力基本一致,但较少的数量不足以让弱入侵区的红火蚁种群支配绝大多数的食物资源,这也很好地解释了强入侵区和弱入侵区中的发现-支配权衡的差异。
我们的研究表明,红火蚁入侵确实打破了本地蚂蚁群落物种的发现-支配权衡,而且随着红火蚁种群规模的增大,红火蚁逐渐从能够影响到完全打破这种发现-支配权衡,为揭示红火蚁的入侵机制提供了的理论依据。但我们的研究仍存在一定的缺陷,如未能完全解释不同种群数量红火蚁在新入侵地的觅食策略,需要后续进行种群数量控制的试验进行补充。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:08:22
科普童话·神秘大侦探(2022年4期)2022-05-26 19:57:56
作文小学高年级(2022年2期)2022-03-03 08:54:32
中学生数理化·八年级物理人教版(2021年12期)2021-12-31 03:23:00
中老年保健(2021年4期)2021-08-22 07:08:26
今日农业(2021年8期)2021-07-28 05:56:12
学苑创造·A版(2021年4期)2021-04-18 13:48:32
销售与市场(营销版)(2021年4期)2021-04-16 03:27:04
系统工程与电子技术(2016年7期)2016-08-21 13:59:16
现代防御技术(2016年1期)2016-06-01 12:13:27
