
外来西方蜜蜂放蜂密度对云南罗平本地传粉蜂群落结构及物种多样性的影响
2020-05-22 06:18:24胡晓玲任晓晓马方舟孟柏达黄永权吴水菊顾忠堂
昆虫学报 2020年3期
胡晓玲, 任晓晓, 马方舟, 孟柏达, 张 睿, 黄永权,吴水菊, 顾忠堂, 吴 军,*, 董 坤,*
(1. 云南农业大学东方蜜蜂研究所, 云南农业大学动物科学技术学院, 昆明 650201; 2. 环境保护部南京环境科学研究所, 南京 210042;3. 红河学院生命科学与技术学院, 云南蒙自 661199; 4. 云南省罗平县蜂业管理站, 云南罗平 655800)
西方蜜蜂Apismellifera于20世纪初被引入到我国,目前已成为我国饲养量较大的外来经济蜂种(季荣等, 2003),其饲养密度在大部分地区已超过当地原有蜂种,部分地区本土中华蜜蜂Apisceranacerana(简称“中蜂”)种群数量锐减达80%以上,部分地区山林中已很难找到野生中蜂种群(任晓晓等, 2016)。而人工饲养的中蜂现已逐渐被排挤到山区和半山区,原先的集群成片分布渐渐转变成分散的零星分布(余林生和孟祥金, 2001)。西方蜜蜂作为外来物种,其采集力和生产性能较好,在利益驱动下人为地不断扩大饲养规模和种群数量,加之采用大转地追花夺蜜的放蜂模式,已对我国宝贵的中蜂资源造成了严重影响,甚至对本土其他野生传粉蜂的物种多样性造成诸多不利影响(何伟志等, 2013),导致一些野生植物或非放蜂期的农作物出现授粉危机,进而导致某些地区或局部的物种消失、生物多样性降低,久而久之可能对我国农业生产和生态平衡造成严重威胁。
近年来,越来越多的专家学者关注外来物种对引入地生态后果的评价,并认识到人类有意或无意引入外来物种已成为严重干扰本地原有传粉昆虫物种多样性最为重要的因素(Steffan-Dewenter and Tscharntke, 2000)。外来蜂种对我国传粉昆虫影响的研究大多关注在中蜂和熊蜂资源上。由于受到取样方法、生境间数据可比性较低等方面的限制,国内很少开展西方蜜蜂不同放蜂强度对我国本土传粉蜂群落结构及物种多样性影响的比较研究。尽管本地传粉蜂类已与当地各相关物种长期协同进化,相互适应,但引进的外来西方蜜蜂通常会与本地其他传粉蜂生态位形成大部分重叠,从而在食物(季荣等, 2003)、巢址(Jason and Liu, 2012)、交尾(袁晓龙等, 2018)等方面存在竞争。美国、印度等国学者研究表明,西方蜜蜂凭借其种群数量优势迫使本地传粉蜂访问回报较少、质量较差的蜜粉源植物,最终导致本地传粉蜂种繁殖力下降(Gross, 2001; Fayez and Shereen, 2006);其次,西方蜜蜂有较高的取食效率和竞争优势,一次可取食90%左右的花蜜,大大降低了本地传粉蜂在单位时间内获取的食物量(Fayez and Shereen, 2006)。当这种食物竞争与干扰随着时间、强度不断增加时,可能会使很多无竞争优势的传粉蜂繁殖力下降,甚至影响其生存(Yangetal., 2010)。
云南特殊的地形、气候条件及多样化的植被类型孕育了与之相适应的丰富的野生蜜蜂资源,云南作为动植物王国,本地传粉蜜蜂对于维持其植物群落结构及稳定性具有不可替代的重要作用。近年来,随着交通条件的改善,每年早春全国大量西方蜜蜂蜂场开始涌向云南蜜源条件较好的罗平、玉溪、楚雄等地放蜂,甚至到一些深山区放蜂,已对本地传粉网络形成了周期性干扰,促使我们进一步开始关注大量西方蜜蜂对云南本地传粉蜂的影响及其生态后果。本研究以云南罗平为研究区域,基于罗平主要蜜源油菜花期西方蜜蜂放蜂密度,利用马来氏网(Malaise trap)收集、监测本地传粉蜂种类、数量周年变化,分析西方蜜蜂放蜂密度对本地传粉蜂群落结构及其物种多样性的影响,以期为合理管理西方蜜蜂放蜂、保护云南本地传粉蜂物种多样性,维持植物群落结构及其稳定性提供参考。
1 材料与方法
1.1 研究区域概况及试验点选择
云南省罗平县(103°57′~104°43′E, 24°31′~25°25′N)位于滇东高原向黔西高原过渡的斜坡上,地势西北高,东南低,属北温带与北亚热带高原季风气候,年平均降雨量1 743.9 mm,是全省多雨区之一,年平均相对湿度85%,年平均日照时数1 685 h,年平均气温15.1℃,年均无霜期为280 d左右。山林植被以针叶林为主,农田作物以油菜、烤烟、生姜、玉米为主,境内生物资源种类繁多,生物多样性较丰富。
罗平县年种植油菜70 余万亩,作为“全国蜜蜂春繁基地”,每年油菜花期平均接收西方蜜蜂春繁蜂群5万余群。本研究根据往年罗平县油菜花期西方蜜蜂放蜂情况,选择了鲁特(LT)、走马田(ZMT)、马把(MB)、外纳(WN)、下阿列(XAL)和下庄科(XZK) 6个自然村为试验点,这些相邻试验点距离为5~6 km。我们围绕每个试验点调查了半径为3 km范围内西方蜜蜂的蜂场数量、蜂群数量、平均群势等信息(表1),其中每个试验点的平均群势统计方法为,首先对于一个蜂群,通过对蜂群开箱检查,蜂箱内一个完整巢脾其两侧脾面爬满蜜蜂且不重叠,无空隙,脾面上的这些数量的成年工蜂即为一足框(脾)蜂,以此标准检查所有巢脾并估算整个蜂群的群势。对于一个蜂场,随机抽取蜂群总数的10%开箱检查,以出现频次最多的蜂群群势作为整个蜂场的平均群势。对于每个试验点,基于所有蜂场出现频次最多的群势作为整个试验点的平均群势。由于每个试验点蜂群群势变化不大,因此主要根据试验点周围的蜂群数量,确定了3个放蜂密度梯度,其中鲁特(LT)和走马田(ZMT)为高放蜂密度区域,外纳(WN)和下阿列(XAL)为中放蜂密度区域,马把(MB)和下庄科(XZK)为低放蜂密度区域。
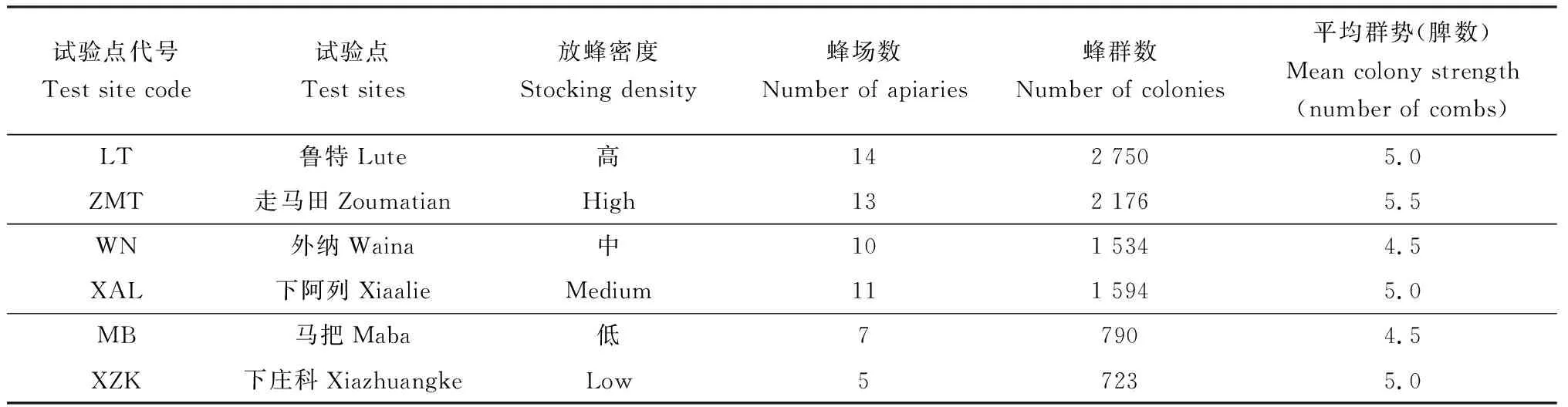
表1 云南省罗平县6个试验点周围3 km范围内的西方蜜蜂放蜂情况Table 1 Stocking states of Apis mellifera in an area with a radius of about 3 km around each of the six test sites in Luoping County, Yunnan Province
1.2 传粉蜂类样本采集
2015年3月至2017年5月,在每个试验点中心位置附近分别选择山林和农田2种生境类型,在每种生境分别架设一张马来氏网,同一试验点2种生境中的马来氏网相距约1 200 m,对本地传粉蜂类开展周年连续调查。马来氏网由昆虫收集瓶、白色纱网和黑色纱网组成,前高后低。收集瓶中盛装无水乙醇,便于收集及保存飞入纱网中的昆虫。每隔一周更换一次收集瓶,标记采集日期、地点、编号等信息,带回实验室4℃保存备用。
1.3 传粉蜂类标本分类鉴定及数量统计
对收集瓶内的昆虫进行初步分类,分拣出所有的传粉蜂类。请相关专家对传粉蜂类进行形态鉴定,所有标本的鉴定尽量鉴定到种或属,统计其种类和数量。
1.4 数据处理
Shannon-Wiener多样性指数(H′):H′=-∑(Pi×lnPi)(其中Pi=Ni/N),表示群落复杂程度(Shannon and Weaver, 1963);Pielou均匀度指数(J′):J′=H′/lnS,表示群落内物种分布的均匀程度(Pielou, 1966);Margalef物种丰富度指数(Dm):Dm=(S-1)/lnN,表示群落中生物种类丰富程度(Margalef, 1958)。式中:N=物种总个体数;Ni=第i种的个体数;Pi=群落中第i个物种的个体数占群落总个体数的比率;S=群落中物种数目。
Berger-Parker物种优势度指数(Mmax):根据物种个体数占总个体数的百分比Mmax来确定,当Mmax≥10%时为优势种,Mmax<1%为稀有种,1%≤Mmax<10% 为常见种。优势类群以特定类群个体数占所有类群个体数的百分比M来确定,当M≥10%时为优势类群,M<1%时为稀有类群,1%≤M<10%为常见类群(戈峰, 2008)。
利用单因素方差分析(one-way ANOVA) 和最小显著差数法(LSD)比较不同放蜂密度下本地传粉蜂的物种多样性差异;采用 Pearson相关系数检验本地传粉蜂物种数、个体数及物种多样性指数与西方蜜蜂放蜂密度(各试验点的放蜂密度用蜂群数量表示)之间的关系;利用成对样本t检验法比较不同生境类型下的传粉蜂个体数及物种数差异。所有数据、图形使用 Excel 2007和SPSS 23.0进行处理。
2 结果
2.1 罗平本地传粉蜂类物种组成
通过2年多的(2015年3月-2017年5月)连续调查,对采集到的所有本地传粉蜂种进行形态鉴定,共采集到本地传粉蜂1 715头,经鉴定隶属于10科53种,其中有42种能确定属名,26种能确定种名。对这些本地传粉蜂外部形态特征对比,可以确定所采集标本为独立的种类,但因涉及类群较广,部分本地传粉蜂目前尚无法确定具体的属、种名。这些本地传粉蜂中,蜜蜂科、隧蜂科和胡蜂科3个类群为优势类群,所占比例为68.1%;切叶蜂科、土蜂科、泥蜂科、方头泥蜂科、青蜂科6个类群为常见类群,所占比例为31.49%,分舌蜂科及地蜂科为稀有类群,所占比例为0.41%。物种优势度指数Mmax大于3%的主要传粉蜂种共6种,分别为东方蜜蜂Apiscerana、黄芦蜂Ceratinaflavipes、拟黄芦蜂Ceratinahieroglyphica、切淡脉隧蜂Lasioglossumexcisum、柑马蜂Polistesmandarinus和变侧异腹胡蜂Parapolybiavaria,其中东方蜜蜂和切淡脉隧蜂为优势种(表2)。
2.2 西方蜜蜂不同放蜂密度对罗平本地传粉蜂的影响
通过对西方蜜蜂不同放蜂密度下的罗平本地传粉蜂进行调查统计,可知西方蜜蜂高放蜂密度下,采集到罗平本地传粉蜂共36种313头,中放蜂密度下,采集到传粉蜂共43种522头,低放蜂密度下,采集到传粉蜂共48种830头(表3),表明各科传粉蜂的物种数量和个体数量均随西方蜜蜂的放蜂密度变化而变化,表现为西方蜜蜂放蜂密度降低,本地传粉蜂物种数量和个体数量随之增加,尤其在蜜蜂科、隧蜂科和胡蜂科3个优势类群的个体数量上变化更为明显。
对本地传粉蜂种类和数量分别进行方差分析和多重比较,结果表明西方蜜蜂低密度放蜂时,本地传粉蜂物种数显著多于西方蜜蜂高密度和中密度放蜂时的情况(P<0.05),但中密度和高密度放蜂时的本地传粉蜂物种数量之间差异不显著(P=0.079)(图1: A);西方蜜蜂低放蜂密度时的本地传粉蜂个体数也显著多于高密度和中密度放蜂时的情况(P<0.05),但中密度和高密度放蜂时的本地传粉蜂个体数量之间差异不显著(P=0.096)(图1: B)。
采用Pearson相关系数进行相关分析,结果表明本地传粉蜂物种数与西方蜜蜂放蜂密度(蜂群数量)呈显著负相关(r=-0.707,P<0.05);本地传粉蜂个体数也与西方蜜蜂放蜂密度梯度呈显著负相关(r=-0.743,P<0.01)。
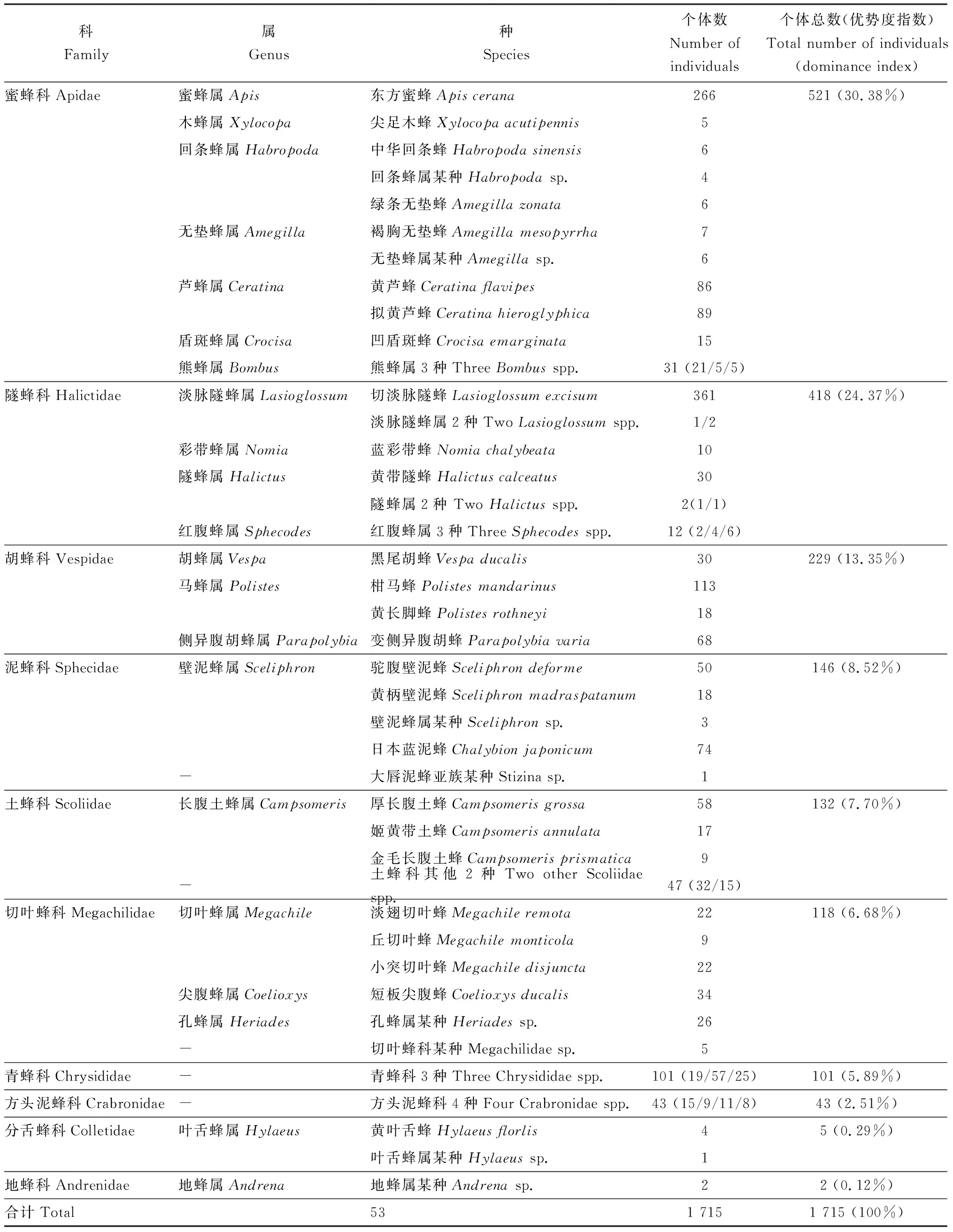
表2 云南罗平本地传粉蜂物种组成Table 2 Species composition of native pollinating bees in Luoping, Yunnan
-: 尚未鉴定到属的种 Have not yet been identified to genus. 优势度指数为特定类群个体数占所有类群个体数的百分比 Dominance index is the percentage of the number of individuals in a specific group accounting for the number of individuals in all groups.

表3 西方蜜蜂不同放蜂密度下云南罗平本地传粉蜂的物种和个体数量Table 3 Numbers of species and individuals of native pollinating bees under different stocking densities of Apis mellifera in Luoping, Yunnan
西方蜜蜂高放蜂密度试验点(鲁特和走马田)周围3 km的蜂群数量为2 176~2 750群,中放蜂密度试验点(外纳和下阿列)周围3 km的蜂群数量为1 534~1 594群,低放蜂密度试验点(马把和下庄科)周围3 km的蜂群数量为723~790群。The number ofA.melliferacolonies within 3 km around the test sites Lute and Zoumatian with high stocking density ofA.melliferais 2 176-2 750, that ofA.melliferacolonies within 3 km around the test sites Waina and Xiaalie with medium stocking density ofA.melliferais 1 534-1 594, and that ofA.melliferacolonies within 3 km around the test sites Maba and Xiazhuangke with low stocking density ofA.melliferais 723-790. 下同The same below.

图1 西方蜜蜂不同放蜂密度下云南罗平本地传粉蜂物种数量(A)及个体数量(B)比较Fig. 1 Comparison of species number (A) and individual number (B) of native pollinating bees under different stocking densities of Apis mellifera in Luoping, Yunnan图中数据为平均值±标准误;柱上不同小写字母表示差异显著(P<0.05, 单因素方差分析和最小显著差数法)。Data in the figure are mean±SE. Different small letters above bars indicate significant difference (P<0.05, one-way ANOVA and LSD). 下图同The same below.
通过对西方蜜蜂不同放蜂密度下蜜蜂科、隧蜂科和胡蜂科3个优势类群的个体数量进行方差分析和多重比较,结果表明这3个传粉蜂优势类群的个体数量在西方蜜蜂高、中、低放蜂密度下存在显著差异,高放蜂密度下的蜜蜂科和胡蜂科个体数量均极显著低于中放蜂密度(P<0.01),高放蜂密度下的隧蜂科个体数量显著低于中放蜂密度(P<0.05),中放蜂密度下,蜜蜂科、隧蜂科和胡蜂科个体数均极显著低于低放蜂密度(P<0.01)(表4)。
3个优势类群个体数随西方蜜蜂放蜂密度变化显著,主要原因是西方蜜蜂放蜂密度对3个类群中的主要传粉蜂种个体数量影响较大。对西方蜜蜂不同放蜂密度下的6种主要传粉蜂的个体数量进行方差分析和多重比较,结果表明高放蜂密度下的东方蜜蜂个体数量极显著低于中放蜂密度(P<0.01),其余5种传粉蜂个体数量均显著低于中放蜂密度(P<0.05);中放蜂密度下东方蜜蜂和变侧异腹胡蜂个体数量显著低于低放蜂密度(P<0.05),其余4种传粉蜂个体数量均极显著低于低放蜂密度(P<0.01)(表5)。
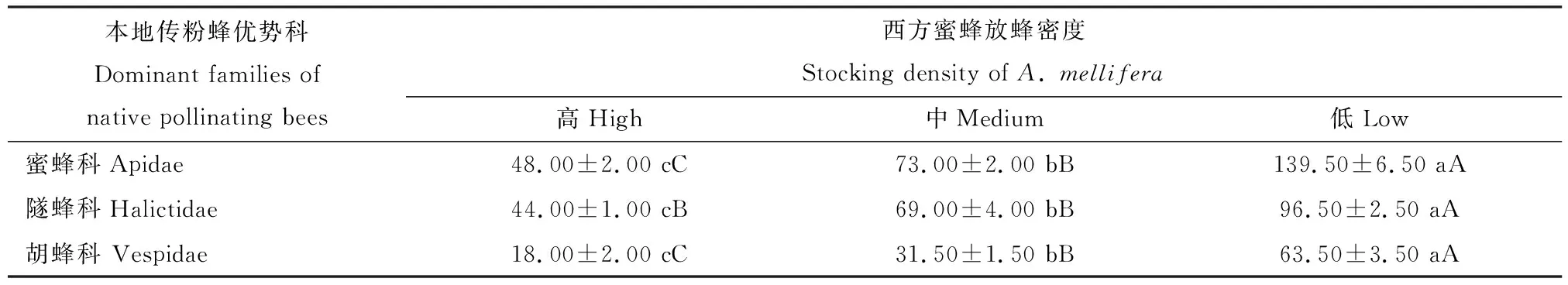
表4 西方蜜蜂不同放蜂密度下云南罗平本地传粉蜂3个优势类群个体数量比较Table 4 Comparison of individual numbers of three dominant groups of native pollinating bees under different stocking densities of Apis mellifera in Luoping, Yunnan
表中数据为平均值±标准误;同行数据后不同小写和大写字母分别表示差异显著(P<0.05)和极显著(P<0.01)(单因素方差分析和最小显著差数法)。Data in the table are mean±SE. Different small and capital letters following the data in the same row indicate significant differences (P<0.05) and extremely significant differences (P<0.01), respectively, by one-way ANOVA and LSD. 表5同The same for Table 5.

表5 西方蜜蜂不同放蜂密度下云南罗平本地主要传粉蜂种的个体数量比较Table 5 Comparison of individual numbers of major native pollinating bees under different stocking densities of Apis mellifera in Luoping, Yunnan
本地传粉蜂物种数量及个体数量不仅随西方蜜蜂放蜂密度的变化而变化,而且在同一放蜂密度下本地传粉蜂物种数量及个体数量还因生境类型(山林或农田)的不同而异。对西方蜜蜂同一放蜂密度下不同生境类型间本地传粉蜂物种数量和个体数量进行成对样本t检验,结果表明西方蜜蜂同一放蜂密度下,山林生境的本地传粉蜂物种数量高于农田生境,其中,高放蜂密度下,山林生境和农田生境的本地传粉蜂物种数量存在显著差异(t=23.00,P<0.05)(图2: A),中放蜂密度下也存在显著差异(t=15.00,P<0.05)(图2: B);但低放蜂密度下差异不显著(t=11.00,P=0.058)(图2: C)。西方蜜蜂同一放蜂密度下,山林生境的本地传粉蜂个体数量高于农田生境,且高放蜂密度(t=17.571,P<0.05)(图3: A)、中放蜂密度(t=52.00,P<0.05)(图3: B)和低放蜂密度(t=41.00,P<0.05)(图3: C)下两种生境本地传粉蜂个体数量均存在显著差异。
2.3 西方蜜蜂不同放蜂密度对罗平本地传粉蜂物种多样性的影响
计算西方蜜蜂不同放蜂密度下,本地传粉蜂类的多样性指数(H′)、均匀度指数(J′)及物种丰富度指数(Dm),并进行多重比较,结果表明西方蜜蜂低放蜂密度时本地传粉蜂多样性指数及丰富度指数均显著高于高密度和中密度放蜂时的情况(P<0.05),但高放蜂密度与中放蜂密度下的本地传粉蜂多样性指数(P=0.574)及丰富度指数(P=0.147)均无显著性差异(图4: A, B);但本地传粉蜂的均匀度指数(J′)却不受西方蜜蜂放蜂密度的影响(图4: C)。

图2 西方蜜蜂相同放蜂密度不同生境类型下云南罗平本地传粉蜂物种数量比较Fig. 2 Comparison of species number of native pollinating bees in different habitat types under the same stocking density of Apis mellifera in Luoping, YunnanA: 西方蜜蜂高放蜂密度试验点(试验点周围3 km范围内蜂群数量为2 176~2 750群)Test sites with high stocking density of A. mellifera (the number of A. mellifera colonies is 2 176-2 750 colonies within 3 km around the test site); B: 西方蜜蜂中放蜂密度试验点(试验点周围3 km范围内蜂群数量为1 534~1 594群)Test sites with medium stocking density of A. mellifera (the number of A. mellifera colonies is 1 534-1 594 colonies within 3 km around the test site); C: 西方蜜蜂低放蜂密度试验点(试验点周围3 km范围内蜂群数量为723~790群)Test sites with low stocking density of A. mellifera (the number of A. mellifera colonies is 723-790 colonies within 3 km around the test site). 图中数据为平均值±标准误;柱上不同字母示差异显著(P<0.05, t检验)。Data in the figure are mean±SE. Different letters above bars indicate significant difference (P<0.05, t-test). 图3和4同The same for Figs. 3 and 4.
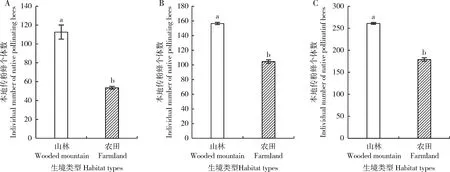
图3 西方蜜蜂相同放蜂密度不同生境类型下云南罗平本地传粉蜂个体数量比较Fig. 3 Comparison of individual number of native pollinating bees in different habitat types under the same stocking density of Apis mellifera in Luoping, Yunnan
相关性分析结果表明本地传粉蜂类多样性指数(H′)和丰富度指数(Dm)变化均与西方蜜蜂放蜂密度(蜂群数量)呈极显著负相关(H′:r=-0.890,P<0.01;Dm:r=-0.962,P<0.01)。

图4 西方蜜蜂不同放蜂密度下云南罗平本地传粉蜂多样性指数(A)、丰富度指数(B)和均匀度指数(C)比较Fig. 4 Comparison of diversity index (A), richness index (B) and evenness index (C) of native pollinating bees under different stocking densities of Apis mellifera in Luoping, Yunnan
3 讨论
本研究通过对罗平县西方蜜蜂不同放蜂密度下的本地传粉蜂进行连续采集,首次研究西方蜜蜂放蜂密度对本地传粉蜂群落结构及物种多样性的影响。经过两年来的持续调查,采集到本地传粉蜂共1 715头,经鉴定隶属于10科53种,西方蜜蜂不同放蜂密度下本地传粉蜂的种类和个体数量差异显著,随西方蜜蜂放蜂密度升高,传粉蜂物种数量、个体数量及物种多样性指数、丰富度指数均降低;同一放蜂密度下,山林生境的本地传粉蜂物种和个体数量均大于农田生境。为了较为准确地掌握罗平本地传粉蜂的种类,分析西方蜜蜂对本地传粉蜂群落结构的影响,本研究开展了连续两年的系统调查。通过对比分析发现,西方蜜蜂不同放蜂密度下本地传粉蜂物种数和个体数两年间数据整体变化不大,所得结论较为一致,本文因此将不同年份的数据合并分析,同时也便于从整体上说明西方蜜蜂放蜂密度对本地传粉蜂群落结构及多样性的影响。
每年油菜花期,全国各地大量西方蜜蜂蜂场涌入罗平县放蜂,且沿主要交通线路存在明显的高、中、低密度梯度变化。这一放蜂密度梯度主要反映了蜂场与县城距离的变化,多数养蜂人选择交通便利、离县城近的油菜蜜源区域放蜂。本研究从罗平县西方蜜蜂放蜂密度入手,调查不同放蜂密度下罗平本地传粉蜂的物种组成,分析西方蜜蜂放蜂密度对罗平本地传粉蜂类多样性的影响。在油菜大宗蜜源的大流蜜期,蜜蜂采集半径通常在2~3 km(Steffan-Dewenter and Tscharntke, 2000; 任晓晓等, 2016),本研究相邻试验点之间的距离大约保持为6 km。本研究原计划设立没有西方蜜蜂放蜂的对照试验点,但很难找到没有西方蜜蜂放蜂的试验点,除非到交通不便、距离较远、海拔较高的深山区,即使找到这样的试验点,由于与其他试验点在生态环境方面差异较大,将使得各试验点的本地传粉蜂的调查结果缺乏可比性。因此,本研究在罗平县选择了生态环境基本一致的6个试验点开展试验,根据6个试验点周围3 km范围内西方蜜蜂放蜂密度调查结果,为了便于对比分析,将西方蜜蜂放蜂密度划分为3个梯度,每个梯度包含2个试验点。总之,本研究尽可能考虑蜜蜂采集行为、传粉蜂的采样方法、试验点间数据可比性等因素的影响来设计研究方案,以实现本研究的目的。
从本地传粉蜂的种类数和个体数量看,西方蜜蜂不同放蜂密度梯度的样地间存在显著差异,但从传粉蜂群落结构看,西方蜜蜂放蜂密度对本地传粉蜂个体数量的影响主要表现在罗平传粉蜂的优势类群和优势种上,如蜜蜂科的东方蜜蜂、隧蜂科的切淡脉隧蜂,二者随着西方蜜蜂放蜂密度的升高,其个体数量显著降低。这说明西方蜜蜂与东方蜜蜂、切淡脉隧蜂生态位重叠可能更严重,竞争也愈发激烈。据报道切淡脉隧蜂国内主要分布在云南,国外分布在缅甸、锡金等,其雌性成虫均采食花粉,对其采访的植物具有传粉作用(吴燕如等, 1988)。罗平本地传粉蜂中还存在分舌蜂科和地蜂科这些数量极为稀少的稀有类群,但在西方蜜蜂高密度放蜂样地没有采集到,推测西方蜜蜂长期高密度放蜂可能会使这些稀有类群数量减少甚至灭绝,从而影响罗平本地传粉蜂的物种多样性和遗传资源多样性。
西方蜜蜂和东方蜜蜂对蜜粉源的要求及利用有所不同(余林生和孟祥金, 2001)。与多数本地传粉蜂类相比,西方蜜蜂的个体较大,它们对处于平缓地势的大宗蜜源具有较强的采集能力;而一些本地传粉蜂如东方蜜蜂个体较小,出巢采集勤奋且嗅觉灵敏,能够发现并善于利用分散的零星蜜源(余林生等, 2008)。当食物资源有限且外来竞争者存在优势时,竞争排斥可能会使处于竞争劣势的本地传粉蜂类放弃其原来的采集模式,避开强大竞争者的采集高峰期进行采集(Goulson, 2003)。Steffan-Dewenter和Tscharntke(2000)对意大利蜜蜂Apismelliferaligustica与德国本地野生传粉蜂类种间竞争的研究结果显示,因两蜂种之间绝大部分生态位重叠,食物竞争激烈,意大利蜜蜂迫使本地野生传粉蜂类改变其原有的采集偏好,本研究的结果也验证了这一结论。农田种植油菜较多,而西方蜜蜂蜂群也多数放置于农田,山林存在一些野生蜜源,在本地传粉蜂被迫舍弃农田大蜜源,转而采集山林中零星的回报较少的野生蜜粉源时,其在山林生境所受到的竞争压力相对小于农田,因此出现同一试验点山林生境中的传粉蜂种数量、个体数量及物种多样性等均比相应农田生境高。
蜜蜂种群数量及其生存范围的扩张与缩小是多种因素共同作用的生态过程,其机理也比较复杂(余林生等, 2006)。有研究表明,栖息环境的变化(如栖息地自然植被减少,人口密度增大,盲目垦殖,滥伐森林,滥施农药化肥)(程青芳等, 2002; 王鑫和何旭, 2012)、食物因素的制约(如植物种类单调,蜜粉源植物贫乏)(何旭和刘新宇, 2011)、蜜蜂病虫害的侵袭(王强等, 2010, 2013)、种间生存竞争(余林生等, 2008)与自然交配的干扰(李位三, 1991)等都是导致蜂类群体缩减的主要原因,并影响到其群体分布。杨冠煌(2005)的研究也表明,引入外来蜂种如意大利蜜蜂是导致本地传粉蜂种如中华蜜蜂数量和分布区域缩减的重要原因。在本研究中,当大量西方蜜蜂转入罗平放蜂,西方蜜蜂数量空前增多,与本地该季节出现的传粉蜂之间的食物竞争就非常激烈,由于西方蜜蜂处于竞争优势,使得本地传粉蜂种的数量大幅减少。本研究验证了外来物种对本地传粉蜂造成的威胁并导致传粉蜂的种类和数量的减少,也表明了罗平本地传粉蜂的物种多样性与西方蜜蜂放蜂密度呈负相关,放蜂密度越大,对本地传粉蜂造成的影响越大,即传粉蜂种类和数量越少,传粉蜂物种多样性越低。为了兼顾养蜂生产和本地传粉蜂的保护,根据本研究结果,建议罗平县西方蜜蜂放蜂密度控制在本研究中所定的低放蜂密度,即半径3 km范围内蜂群数量750群左右,按罗平县早春油菜有效蜜源面积(因该县山地面积占90%以上)计算,大约相当于2~3群西方蜜蜂/hm2。
年复一年的高密度放蜂,本地传粉蜂种类和数量的局部减少最终可能导致一些传粉蜂类的灭绝。面对这一亟待解决的问题,一方面需要合理控制西方蜜蜂的放蜂密度,减少对本地传粉蜂种干扰与竞争,另一方面也需要从食物竞争、病虫害跨种传播、生殖干扰等多角度进一步揭示外来蜂种对本地传粉蜂种多样性影响的机制。
猜你喜欢
河南农业(2024年3期)2024-03-05 02:20:52
中国科学探险(2022年8期)2022-12-31 07:27:54
Chinese Physics B(2022年6期)2022-06-29 08:53:46
奥秘(2022年5期)2022-05-31 02:22:41
中学生学习报(2022年1期)2022-03-21 02:01:29
疯狂英语·新悦读(2022年12期)2022-03-08 02:37:12
作文周刊·小学一年级版(2022年4期)2022-03-07 01:13:49
边疆文学(2020年5期)2020-11-12 02:29:46
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25 01:40:26
现代农业(2018年7期)2018-08-01 12:27:44
