
转运系统集成改造提升大肠杆菌胞外蛋氨酸积累
2020-05-19 02:03李由然丁重阳石贵阳
食品与生物技术学报 2020年3期
董 伟, 李由然, 张 梁, 丁重阳, 石贵阳*
(1. 江南大学 粮食发酵工艺与技术国家工程实验室,江苏 无锡 214122;2. 江南大学 生物工程学院,江苏 无锡214122)
蛋氨酸作为一种含硫氨基酸被广泛应用于饲料添加,食品制造,生物制药等领域[1-3],目前,蛋氨酸的生产仍然停留在利用化石燃料通过化工合成的办法满足市场需求, 对生态环境造成很大压力。微生物发酵法生产L-蛋氨酸因其低成本与环境友好型等特点,受到越来越多专家学者的亲赖。 通过随机诱变和代谢工程可以选育蛋氨酸高产菌株,但前者不确定性较强,工作量较大。 随着全基因组测序技术和系统代谢工程的日趋成熟,使理性改造菌株发酵生产蛋氨酸成为代谢合成研究的热点[4]。
目前,微生物代谢工程改造增强蛋氨酸合成主要通过加强蛋氨酸合成酶编码基因metB[5-6]、S-腺苷甲硫氨酸合成酶编码基因metK[7]、解除反馈抑制的高丝氨酸转琥珀酰基酶HTSFbr[8]和高丝氨酸琥珀酰基转移酶编码基因metA[9]的表达等方式来增强碳代谢途径,敲除MetJ 阻遏蛋白编码基因metJ、MetD 运输系统的编码基因metNIQ、赖氨酸合成途径编码基因lysA 和苏氨酸合成途径编码基因thrC 等方式阻断支路降解途径,同时也有关于硫代谢途径和葡萄糖运输途径修饰相关的研究报道[10,11]。微生物代谢合成网络结构复杂,为了避免代谢流流向一些未知的支路途径和减少胞内氨基酸的过量积累对菌体生长造成负担,氨基酸转运系统成为一些研究者关注的重点,如本研究室曾发现转运蛋白对苏氨酸代谢合成起到非常重要的作用[11]。大肠杆菌蛋氨酸MetD吸收转运系统编码基因metN、metI 和metQ 的全部敲除可以降低E.coli W3110 胞外蛋氨酸的吸收速率,YjeH 和YeaS 分泌系统可以加强蛋氨酸向胞外转运,但目前并没有将两者相结合进行胞外蛋氨酸积累影响的报道[12]。
本研究中以E.coli W3110 为出发菌株,通过比较metN、metI 和metQ 单基因敲除菌株与metN、metI 和metQ 的全部敲除菌株胞外蛋氨酸吸收速率, 选择胞外吸收速率最小的菌株进行YjeH 和YeaS 分泌系统研究,阐述弱化蛋氨酸吸收与增强蛋氨酸分泌集成改造策略对胞外蛋氨酸积累的影响。
1材料与方法
1.1 材料
1.1.1 菌株与质粒 本研究中所用的基座菌株E.coli W3110 为实验室保藏, 基因敲除所需要的质粒pKD13、pKD46、pCP20 购于美国耶鲁大学大肠杆菌菌 株 库 (CGSC,E.coli Genetic Stock Center,New Haven,USA)[12]。 实验室构建和保藏的质粒pETABY, 由pN25 启动子控制蛋氨酸操纵子编码框pET-ABY 的表达,metA、metB、malY 分别编码高丝氨酸琥珀酰转移酶、γ-胱硫醚聚合酶、β-胱硫醚酶。重 组 菌M10、M11、M12、M13、M14、M15、M16 为 本研究构建,各菌株及质粒见表1。
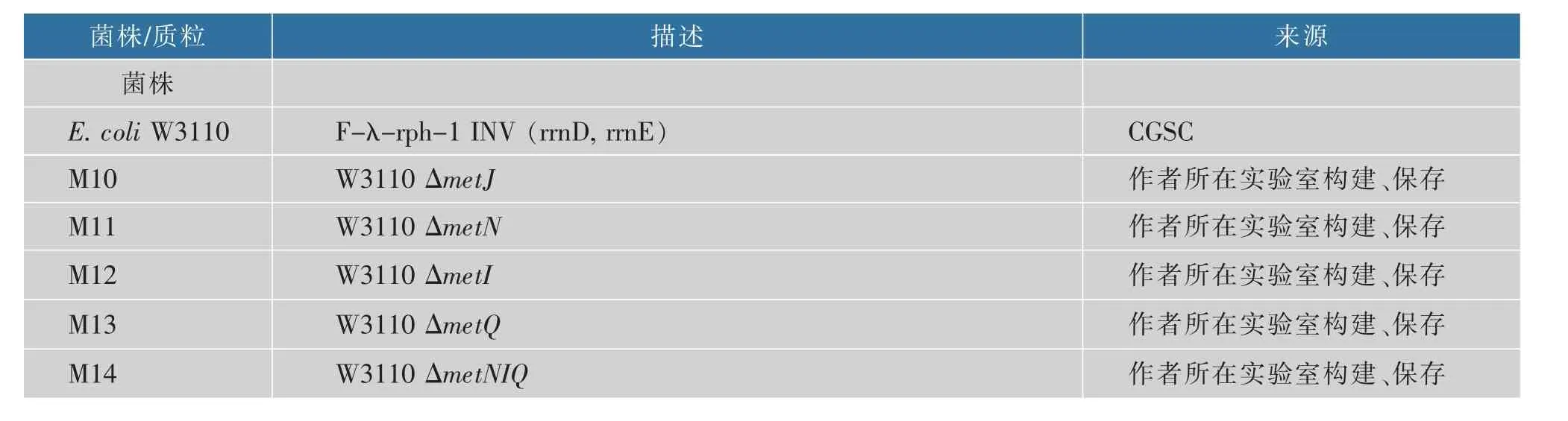
表1 本研究中主要的菌株和质粒Table 1 Main strains and plasmids used in this study
1.1.2 工具酶及试剂 实验所用的RNA 提取试剂盒、2×taq PCR master mix、 质粒DNA 提取试剂盒,购自杭州宝赛生物公司;胰蛋白胨、琼脂粉、酵母粉,购自英国Oxoid 公司;琼脂糖,购自Biowest 公司;氨苄青霉素、氯霉素、卡那霉素,购自Sigma 公司;其他试剂均为国产或进口分析纯。

续表1
1.1.3 培养基及培养条件 SOB 培养基(g/L):酵母粉5,KCl 0.186,MgCl20.95,NaCl 0.5,蛋白胨20;转接携带有质粒pKD46 的液体SOB 培养基需另加入7.5 g/L L-阿拉伯糖。
摇瓶发酵培养基为改进的TPM 培养基(g/L)[13]:维 生 素B120.01, 葡 萄 糖 20,MgSO4·7H2O 2,MnSO4·4H2O 0.5, 酵母粉4,KH2PO44,(NH4)2SO414,L-threonine 0.238,L-lysine 0.164,CaCO310,PLP 0.01,生物素0.01,微量元素母液5 mL。 微量元素母液成份(g/L):CuSO4·5H2O 1,CaCl21.35,Na2B4O7·10H2O 0.23,ZnSO4·7H2O 2.25,FeSO4·7H2O 10, (NH4)6Mo7O24·4H2O 0.106,35 g/dL HCl,10 mL, 调整最终pH 为7.0。接种体积分数2%,31°C、250 r/min 摇床培养48 h。
分析培养基(g/L):蛋氨酸0.5,H3BO31,MgSO42.05,MnSO420,(NH4)2SO40.75,NH4Cl 0.13,FeSO440,CaCl240,CoCl28,(NH4)2HPO42,ZnSO44,CuCl22,(NH4)2Mo2O72.8,Na2HPO42,K2HPO44,调整最终pH 6.7。
1.1.4 引物

表2 本实验所用引物Table 2 Oligonucleotides used in this study
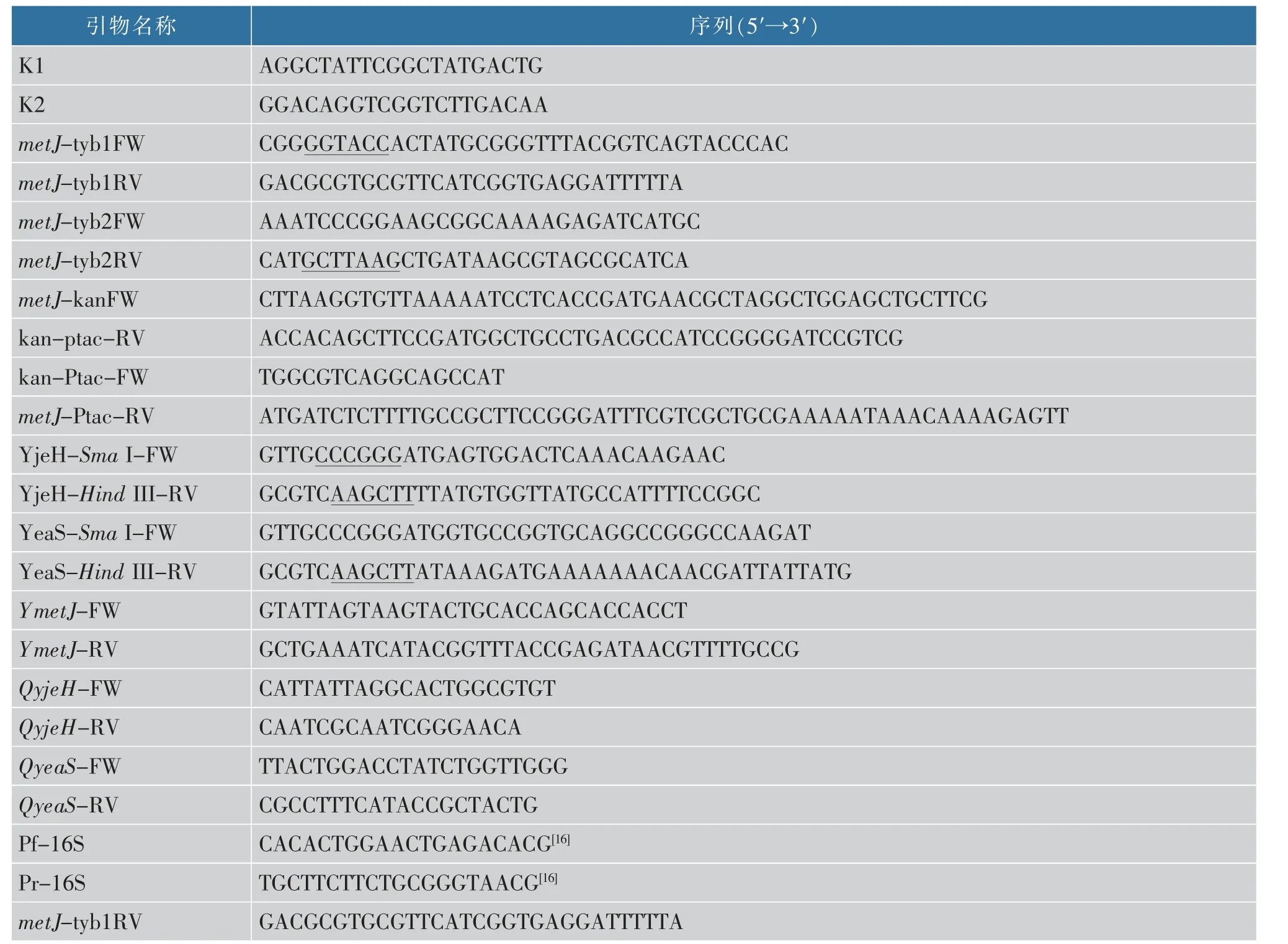
续表2
1.2 方法
1.2.1 基因敲除和基因组替换方法 利用Red 重组技术进行基因敲除[14]。1)PCR 打靶片段的制备:质粒pKD13 作为模版, 以InmetQ-FW/InmetQ-RV、InmetI-FW/InmetI-RV、InmetN-FW/InmetN-RV为引物,2×taq聚合酶扩增片段大小为1.4 kb 的敲除盒metQ、metI、metN。PCR 扩增体系10 μL,2×PCR master mix 5 μL,ddH2O 4.7 μL,模板0.1 μL,引物各0.1 μL。2)突变盒的纯化浓缩:PCR 反应产物凝胶电泳纯化后所得片段中加入1/50 倍体积5 mol/L NaCl和2 倍体积预冷的无水乙醇,于-70 ℃冰箱静置20 min,取出后4 ℃、12000 r/min 离心10 min,去除上清液后得到黏附于管壁的絮状沉淀,加入无水乙醇(预冷),4 ℃、12000 r/min 离心4 min, 用适量ddH2O振荡溶解。 3)基因敲除:制备含有质粒pKD46 的E.coli感受态细胞,电转体系为:感受态细胞,95 μL;突变盒片段,5 μL;电压,1800 V;后培养37 °C,1 h。以YmetQ-FW/YmetQ-RV、YmetI-FW/YmetI-RV、YmetN-FW/YmetN-RV 作为引物进行PCR 验证。4)卡那霉素抗性的消除:制备整合了敲除盒的E.coli菌株的感受态细胞,通过电激转化导入质粒pCP20,利用含有氯霉素抗性的SOB 固体平板进行筛选,挑取转化子进行PCR 验证。
基因组上整合替换,所用引物见表2。打靶片段的制备参见文献[15]。 利用Red 重组在E.coliW3110染色体上单拷贝整合yjeH、yeaS。
1.2.2 菌体生长曲线的测定 分别挑取E. coliW3110、M11、M12、M13 的单菌落于20 mL SOB 液体培养基,37 °C、200 r/min 过夜培养, 转接50 mL FM 培养基,37 ℃、200 r/min 培养,每隔2 h 取样检测分光光度计600 nm 下的菌体密度,测定生长曲线。
1.2.3 菌体生物量的测定 菌体浓度以分光光度计600 nm 下的吸光值OD600表示,菌体干重(DCW)以本实验室的经验公式换算获得:
DCW=OD600×0.3809-0.0048[12]
1.2.4 蛋氨酸的测定 HPLC 柱前邻苯二醛(OPA)自动衍生法测定蛋氨酸的浓度,Waters C18(250 mm×4.6 mm,0.5 μm) 反相柱分离,A、B 流动相梯度洗脱,洗脱程序如表3 所示,柱温40 ℃,紫外检测器检测,检测波长338 nm。
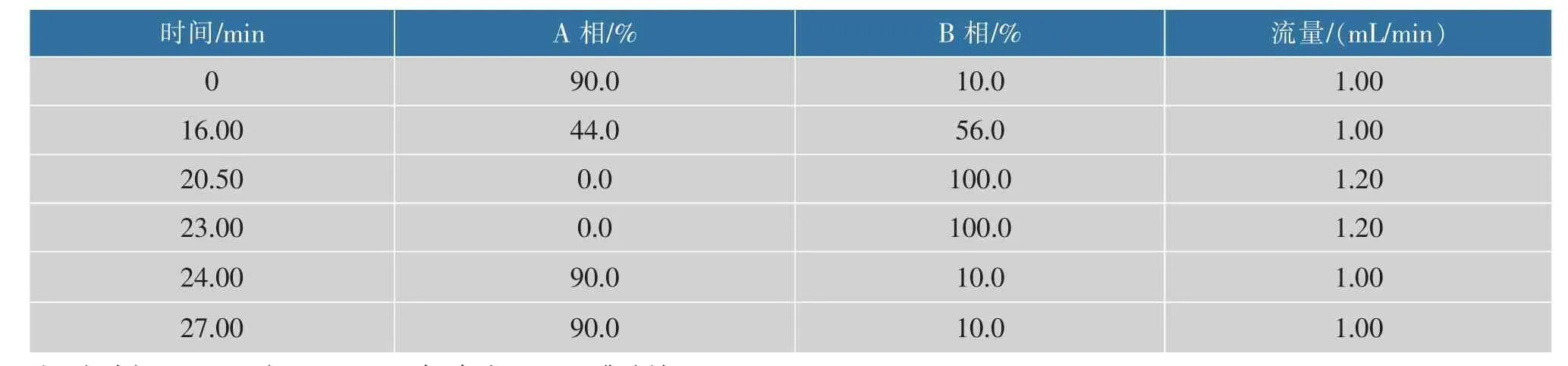
表3 HPLC 蛋氨酸分析洗脱程序Table 3 Gradient elution for analysis of L-methionine by HPLC
1.2.5 荧光定量PCR (RT-qPCR) 利用试剂盒提取大肠杆菌总的RNA,qPCR CT 值的校正以引物pf-16S,pr-16S 扩增16SrRNA 作为内参基因, 相对表达量以2-△△CT模型进行检测[16],所用引物见表2。
1.2.6 蛋氨酸胞内运输能力检测 SOB 培养基中添加0.5 g/L 蛋氨酸作为种子培养基, 过夜培养,诱导菌株表达蛋氨酸吸收载体。 在蛋氨酸分析培养基中添加1 g/L 的蛋氨酸作为碳源, 发酵过程中检测菌体生长过程中蛋氨酸的消耗情况,以蛋氨酸对菌体的比消耗速率表征大肠杆菌吸收蛋氨酸的能力。
2结果与分析
2.1 MetD 运输系统缺失菌株的构建
以E. coliW3110 为出发菌株, 按照1.2.1 的方法敲除metN、metI、metQ,构建菌株M11、M12、M13,结果如图1 所示。metN、metI、metQ的鉴定引物分别为YmetN-FW/YmetN-RV、YmetI-FW/YmetIRV、YmetQ-FW/YmetQ-RV,利用2 对引物与K1、K2 对敲除菌株进行菌落PCR 验证,得到990、740、1170 bp 和1170、1420、1090 bp 的PCR 片段,电泳结果与理论值相符,消除kan抗性后,得到850、880、950 bp 的DNA 片段,送生工测序结果正确,成功构建了metN、metI、metQ基因敲除菌株。
2.2 弱化蛋氨酸吸收基座菌株的筛选

图1 metN、metI、met 突变转化子的PCR 鉴定Fig. 1 PCR identification of metN, metI, metQ gene mutants
按照1.2.6 的方法对大肠杆菌W3110 和重组菌株M11、M12、M13 蛋氨酸吸收能力进行了检测。 图2 表示摇瓶培养10 h 菌体的生长以及培养基中蛋氨酸质量浓度的变化,表4 列出了重组菌株蛋氨酸的比消耗速率。 由图2 可知,在蛋氨酸分析培养基中重组菌与对照菌株均生长缓慢,但是对蛋氨酸的消耗速率显著差异。 由表4 可知, 摇瓶培养10 h,metN、metI、metQ 基因单敲除菌株M11、M12、M13 蛋氨酸平均比消耗速率分别为11.23、10.34、12.03 mg/(L·h),与对照菌W3110 相比蛋氨酸的平均比消耗速率分别降低了9.5%、16.7%、3.1%。 metI 基因的敲除显著降低了大肠杆菌对于胞外蛋氨酸的吸收效率,metN 次之;敲除基因metQ,蛋氨酸吸收能力降低幅度最小。 本研究室前期研究发现,metNIQ 三敲除菌株胞外蛋氨酸吸收速率降低了13.4%[12], 基因metI 单敲除更加有利于减少大肠杆菌对胞外蛋氨酸的重吸收。 因此,本研究将M12 作为弱化蛋氨酸吸收基座菌株与蛋氨酸分泌系统相结合进行集成改造,考察胞外蛋氨酸的积累。
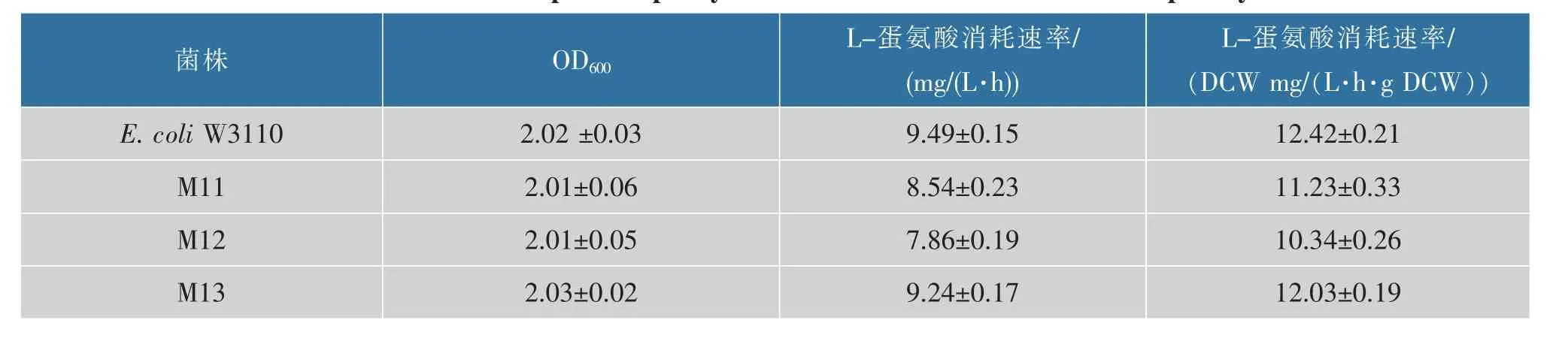
表4 重组菌株蛋氨酸吸收能力Table 4 L-methionine uptake capacity of strains with MetD modified transport system
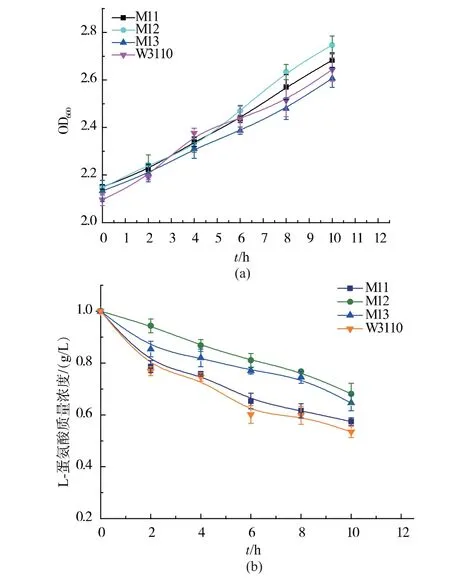
图2 蛋氨酸分析培养基中重组菌株生长状况Fig. 2 Growth status of recombinant strains in methionine assay medium
2.3 YjeH,YeaS 运输系统游离表达对菌株生长以及蛋氨酸胞外积累的影响
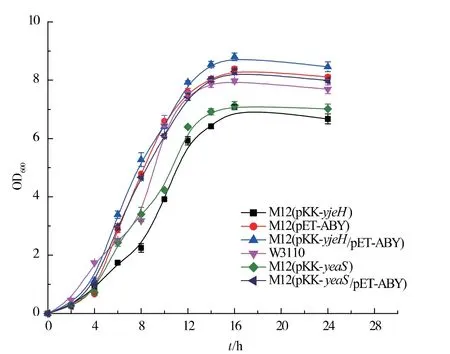
图3 重组菌株E. coli M12、M12(pET-ABY)、M12(pETABY /pKK-yjeH)、M12 (pKK-yjeH)、M12 (pKKyeaS)、M12(pET-ABY /pKK-yeaS)生长曲线Fig. 3 Growth curves of recombinant strains E.coli M12,M12(pET-ABY),M12(pET-ABY/pKK-yjeH),M12(pKK-yjeH),M12(pKK-yeaS) and M12(pET-ABY/pKK-yeaS)mutants
将实验室前期构建的yjeH、yeaS 基因过表达载体pKK-yjeH、pKK-yeaS 转染M12, 记录菌体生长以及胞外蛋氨酸积累情况。从图3 可以看出yjeH 与yeaS 的过表达在一定程度上均抑制了菌体的生长,本研究室前期构建用来表达解除了反馈抑制的高丝氨酸转琥珀酰基酶和增强蛋氨酸代谢过程中的γ-胱硫醚聚合酶以及β-胱硫醚酶[12]的质粒pETABY 的表达可以有效缓解生长抑制。 yjeH、yeaS 的过量表达可能使大肠杆菌胞内蛋氨酸质量浓度过低, 影响了大肠杆菌生长过程中所需要物质的合成,菌体的生长受到抑制,同时胞外蛋氨酸的质量浓度过高也会抑制菌体的生长, 质粒pET-ABY 的表达强化了蛋氨酸合成途径,在基因交互作用的影响下其生长抑制得到了有效缓解。 对比M12(pETABY)、M12 (pET-ABY/pKK-yjeH) 和M12 (pETABY/pKK-yeaS)发酵过程中胞内蛋氨酸的质量浓度变化,如图4 所示,YjeH、YeaS 运输系统的强化促进了大肠杆菌向胞外分泌L-蛋氨酸, 降低了胞内L-蛋氨酸的积累,YjeH 运输系统的效果更为显著。 我们对2 种转运系统游离表达菌株进行了发酵实验,结果如图5 所示,YjeH 与YeaS 运输系统的表达都能够在胞外积累一定量的蛋氨酸, 其最大积累量分别为0.21 g/L 和0.18 g/L,蛋氨酸产率分别为0.06 g/g DCW 和0.05 g/g DCW。
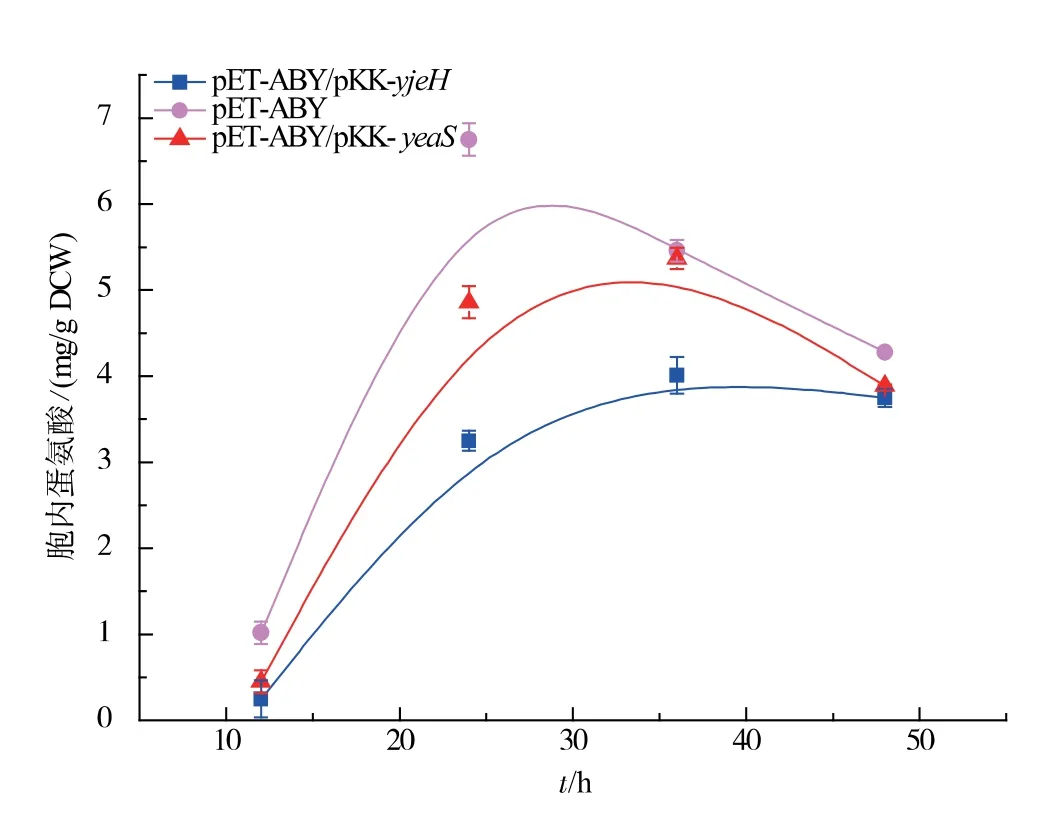
图4 重组菌株发酵过程中胞内蛋氨酸的质量分数Fig. 4 Intracellular methionine concentration during fermentation of recombinant strains
2.4 YjeH 和YeaS 在染色体上的整合表达
metJ为大肠杆菌蛋氨酸代谢过程中的关键阻遏基因, 其通过与代谢过程中关键基因metK、metQIN、metF、metC、metE、metBL和metR等启动子的Met-box 相结合来影响蛋氨酸代谢通路,为了降低SAM 和metJ的共阻遏效应, 我们通过敲除编码基因metJ构建解除阻遏调控的重组菌株[17-20]。 构建打靶质粒T-metJPtacM[15],通过电激转化的方法,在E. coliW3110 染色体上敲除metJ基因并单拷贝整合yjeH和yeaS基因, 一步实现metJ的敲除和yjeH、yeaS在染色体上的整合表达。 以W3110 染色体为模板,用引物YjeH-SmaI-FW/YjeH-HindIIIRV、YeaS-SmaI-FW/YeaS-HindIII-RV PCR 扩增yjeH,yeaS,将所得到的基因片段,与经过HindIII,SmaI 酶切处理过的载体T-metJPtacM 连接, 酶切得到大小分别为1257 bp/5059 bp 和639 bp/5059 bp的片段如图6(a)、6(c)所示,测序结果无碱基突变,说明载 体T-metJPtacMyjeH,T-metJPtacMyeaS构 建 成功。按照1.2.1 的方法将metJPtacMyjeH,metJPtacMyeaS整合到W3110 的染色体上,构建菌株M15、M16,鉴定结果如图6(b)、6(d)所示。
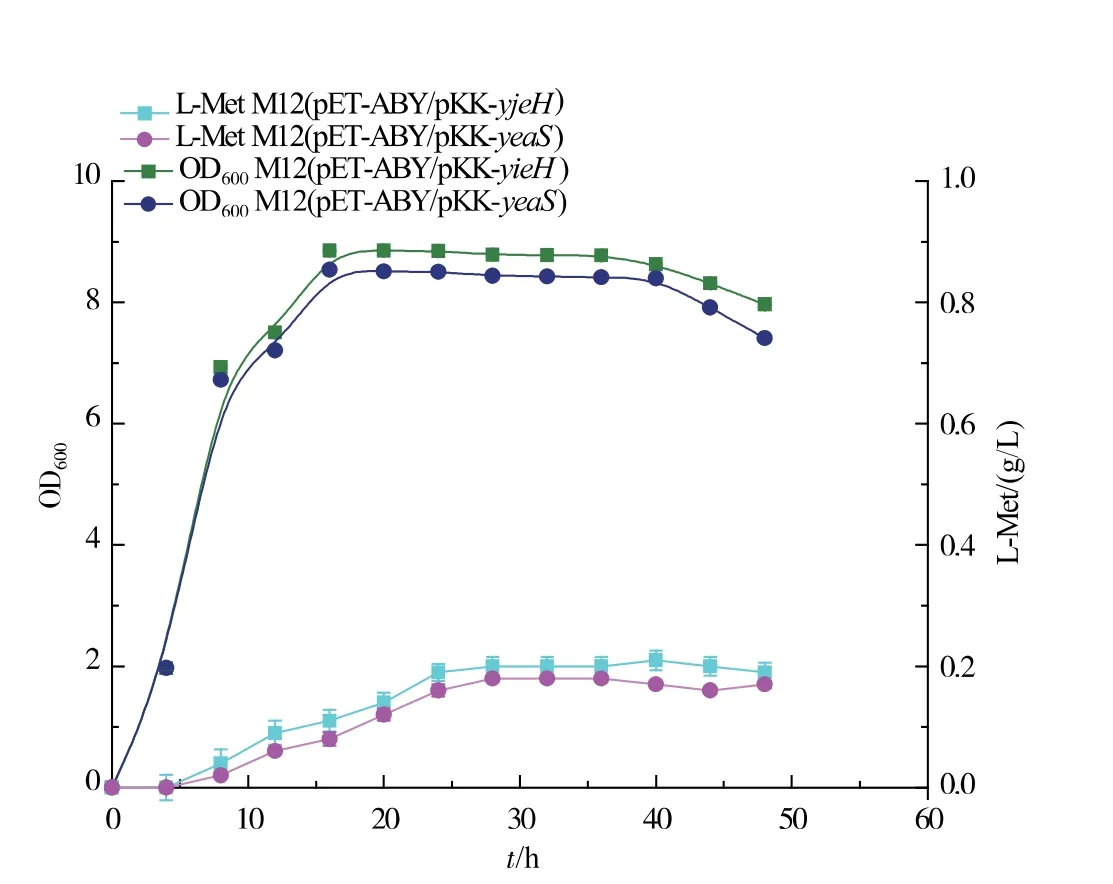
图5 重组菌M12 (pKK-yeaS/pET-ABY)、M12 (pKKyjeH/pET-ABY)摇瓶发酵生长代谢状况Fig. 5 Growth and metabolism of recombinant M12 (pKK-yeaS/pET-ABY) and M12 (pKKyjeH/pET-ABY) during shake flask fermentation
2.5 YjeH 运输系统整合表达对菌株生长以及蛋氨酸胞外积累的影响
利用RT-qPCR 比较了菌株M15 相对于野生E. coliW3110yjeH和yeaS基因的表达比率, 结果如图7 所示,与原始菌株相比,yjeH、yeaS基因的表达水平分别提高了1.65 倍和1.59 倍。对M15(pETABY)和M16(pET-ABY)进行了发酵实验验证基因yjeH、yeaS整合表达对蛋氨酸胞外分泌的影响,如图8 所示,M15(pET-ABY)和M16(pET-ABY)最大胞外蛋氨酸的积累量分别为0.48 g/L 和0.25 g/L,蛋氨酸产率分别为0.13 g/g DCW 和0.08 g/g DCW。与游离表达相比, 其产量分别提高了128%和38.9%,产率提高了85.7%和60%。

图6 菌株M15,M16 的PCR 鉴定电泳图Fig. 6 PCR identification of strain M15 and M16

图7 不同菌株yjeH 转录水平测定Fig. 7 Relative gene transcriptional levels of yjeH in the different strain

图8 重组菌M15(pET-ABY)、M16(pET-ABY)摇瓶发酵生长代谢状况Fig. 8 Growth and metabolism of recombinantM15 (pETABY) and M16 (pET-ABY) during shake flask fermentation
3结 语
随着对微生物氨基酸合成途径的深入了解,转运系统在氨基酸育种研究中的作用越来越受到人们的重视,近年来的研究表明,对氨基酸向胞内吸收和胞外运输途径进行改造,是提高氨基酸产量行之有效的方法。MetD 运输系统是蛋氨酸吸收的主要途径,与过表达代谢途径中促进蛋氨酸合成的关键酶相比,修饰蛋氨酸向胞内的运输系统并不能提高蛋氨酸生物合成代谢流,但能够有效减少组织对胞外蛋氨酸的吸收,降低胞内蛋氨酸的浓度,改善内部环境,从而减弱蛋氨酸对细胞的毒害作用以及降低无效循环,进而可以提高鲁棒性以及氨基酸合成的能力[12]。 本实验以E. coli W3110 为基座菌株,利用Red 重组技术分别获得E. coli W3110 中MetD运输系统的关键组分metN、metI 和metQ 单敲除菌株, 与菌株W3110 相比,metI 的缺失使胞外蛋氨酸吸收速率降低了16.7%。 为了避免多拷贝引起的转基因沉默以及基因表达水平过高抑制菌体生长,对M12 进行yjeH 和yeaS 基因单拷贝整合有效促进了胞外蛋氨酸的积累,同时缓解了游离表达造成的菌体生长抑制现象,M15 (pET-ABY) 和M16 (pETABY) 最大胞外蛋氨酸产量分别提高了128%和38.9%。 可见,在弱化蛋氨酸吸收基座菌株的基础上加强胞内蛋氨酸的分泌进行集成改造可以有效提高胞外蛋氨酸的积累。 尽管很多研究人员分别进行氨基酸转运分泌系统和吸收系统的研究,但将两者有机结合进行系统的分析改造是转运系统工程研究的重要趋势,在以后蛋氨酸以及其他氨基酸生产菌株的理性改造过程中必将发挥愈来愈重要的作用。
猜你喜欢
中国储运(2022年11期)2022-11-30
生物信息学(2022年3期)2022-11-12
安徽建筑大学学报(2022年4期)2022-09-27
物流技术与应用(2022年8期)2022-08-26
中国饲料(2022年5期)2022-04-26
西南农业学报(2021年10期)2021-12-14
国际太空(2021年8期)2021-11-05
昆明医科大学学报(2021年4期)2021-07-23
猪业科学(2020年12期)2021-01-09
农业环境科学学报(2020年9期)2020-10-09
