
糯玉米叶夹角的遗传模型分析
2020-05-11 12:30王军李洪涛许瀚元李景芳祝庆迟铭
江苏农业科学 2020年5期
关键词:糯玉米
王军 李洪涛 许瀚元 李景芳 祝庆 迟铭


摘要: 为了研究控制玉米叶夹角的基因遗传规律,以株型存在显著差异的2个糯玉米自交系组配的P1、P2、F1、BC1、BC2、F2 6个世代为试验材料,运用主基因+多基因遗传模型分析方法,探明控制糯玉米叶夹角的遗传模型,并进行遗传参数估计。结果表明,F1叶夹角杂种优势表现为负向离中亲优势,无超亲优势;糯玉米穗上第一叶叶夹角与穗下第一叶叶夹角遗传模型相同,均受2对加性-显性-上位性主基因控制,穗上第一叶叶夹角BC1、BC2、F2主基因的遗传率分别为63.96%、72.90%、82.55%,穗下第一叶叶夹角BC1、BC2、F2主基因的遗传率分别63.41%、71.83%、82.01%;27.58%的表型变异是由环境因素决定,非加性效应大于加性效应,易晚代进行叶夹角性状选择。研究结果为糯玉米叶夹角性状的基因定位和株型育种选择奠定了理论基础。
关键词: 糯玉米;叶夹角;主基因+多基因;遗传模型
中图分类号:S513.032 文献标志码: A
文章编号:1002-1302(2020)05-0065-06
玉米是重要的饲料、工业原料和粮食作物,在国民经济发展中占有重要地位[1]。2017年我国玉米种植面积达3 813.33万hm2,总产2.16 t,仅次于美国[2]。现阶段通过扩大种植面积增加玉米产量的空间有限,与美国相比,我国的玉米种植密度和单位面积产量仍有上升空间[3-4]。然而单纯提高种植密度将造成群体遮阴综合症,影响玉米植株光合效率和生长模式[5]。因此,改良玉米株型,提高品种的耐密性,逐步成为玉米育种的主要目标和增加玉米产量的重要途径。玉米株型是指玉米植株在空间的分布状态,不同株型会直接影响群体冠层内的光分布状况和群体的光能利用率,最终影响产量的高低[6]。自Donald正式提出了作物理想株型的概念[7]以来,国内外学者从光合、生理、遗传、碳氮代谢等各自研究领域对玉米株型与产量关系进行了研究,提出了耐密理想株型的形态指标,包括叶夹角、叶向值、叶长、叶宽、株高、穗位高、茎节间距、雄穗大小等[8-15]。其中,叶夹角是衡量玉米植株紧凑程度和耐密株型育种的重要形态指标,与受光姿态和群体透光率密切相关,直接影响植株的光合能力,进而影响玉米产量[16-17]。根据叶夹角的差异可以把玉米株型分为紧凑型、中间型和平展型。多项研究结果表明,紧凑型玉米杂交种能够最大限度地提高种植密度,进而增加玉米单位面积的群体产量[18-20]。紧凑型品种由于叶夹角小,叶层间的间隙得到提高,穗上叶截光能力低,增加了群体通透性,利于群体叶片间捕获光能和CO2的合理分布,增强植物光合作用和呼吸作用,提高植株净光合速率和群体生产力,品种耐密性增强,进而提高产量[21-22]。叶夹角的遗传研究是进行株型改良的前提,它是一个复杂的数量性状,国内外关于叶夹角遗传研究多集中在穗上叶夹角的经典遗传学及QTL定位方面,对穗下叶夹角或遗传模型研究较少;而且多项研究结果表明,叶夹角的遗传特点因研究材料和环境的差异而使结果不甚一致[23-41]。因此,对玉米叶夹角的遗传规律仍须进一步研究。本研究拟采用主基因+多基因6个世代联合分析方法对糯玉米穗上第一叶叶夹角和穗下第一叶叶夹角进行遗传模型分析,试图探讨糯玉米叶夹角的遗传规律,旨在为糯玉米叶夹角的基因定位和株型育种改良提供参考依据。
1 材料与方法
1.1 试验材料
选用综合农艺性状较好的叶夹角差异较大的糯玉米自交系为亲本,分别为连玉糯1号母本LN12和广西地方种质环江糯,以LN12×环江糯这个杂交组合的P1、P2、BC1、BC2、F1、F2 6个世代为试验材料。
1.2 试验设计
2017年夏在江苏徐淮地区连云港农业科学研究所试验场(119.31858°E、34.56128°N),利用叶夹角差异较大的糯玉米自交系LN12(P1)和环江糯(P2)为亲本配置杂交组合F1(LN12×环江糯);2014冬在海南省三亚试验地(108.91148°E、18.49811°N)播种亲本及F1杂交种,并配置BC1(LN12/环江糯//LN12)、BC2(LN12/环江糯//环江糯)、F2(LN12×环江糯)。2018年夏在江苏徐淮地区连云港农业科学研究所试验场,6个世代群体同期播种,均采用随机区组设计,重复3次,不分离世代P1、P2和F1群体每个小区各播种3行,每个群体60株,分离世代BC1、BC2和F2群`体每个小区各播种15行,每个群体300株。小区行长4.5 m,行距0.6 m,株距0.22 m。
1.3 性状测定
于吐丝期选择生长状况良好的单株进行穗上第一叶叶夹角与穗下第一叶叶夹角调查。叶夹角为叶片与茎秆之间的锐角角度(用量角器测量);性状的测量均由一人执行,且每个群体的数据采集均在同一时间段內完成,确保数据准确性。P1、P2、BC1、BC2、F1、F2 6个群体分别考察50、50、50、150、150、200株。
1.4 统计分析
利用盖钧镒等提出的植物数量性状遗传体系主基因+多基因混合遗传模型分离分析方法(P1、P2、F1、F2、BC1、BC2 6个世代联合分析方法)[23-24]对糯玉米叶夹角进行主基因+多基因模型分析,根据所选最适模型估算玉米叶夹角相应的主基因和多基因的遗传参数。主基因+多基因6个世代联合分析软件包[25]由南京农业大学章元明教授提供。
2 结果与分析
2.1 糯玉米各世代群体的叶夹角
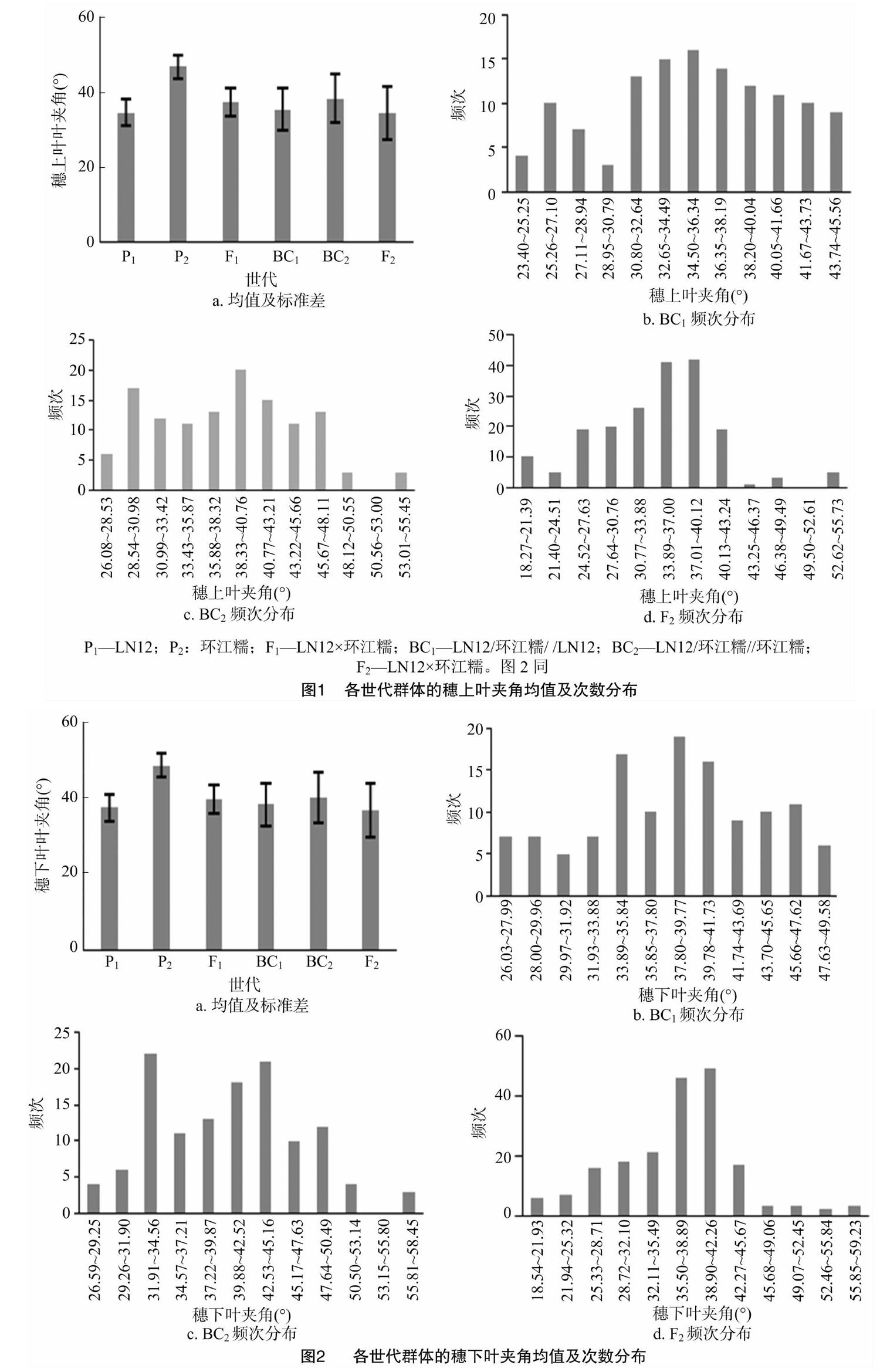
由图1、图2可以看出,各世代穗下第一叶叶夹角平均值均大于穗上第一叶叶夹角,且两者均表现为P1代叶夹角平均值低于P2,说明LN12的叶夹角小于环江糯;F1平均值介于P1与P2之间,且接近P1,说明杂种F1叶夹角杂种优势表现为负向离中亲优势,无超亲优势;3个分离世代群体BC1、BC2、F2叶夹角的次数分布呈双峰或多峰分布,表现为包含多个正态分布的混合分布,表明叶夹角受多个主基因控制,也可能存在多基因的互作影响。
2.2 糯玉米叶夹角遗传模型分析
AIC值表示观测值概率的估计分布与真实分布间的适合性程度。AIC值最小的模型为最佳可能模型。模型间AIC值差异不大时,可以有几个备选模型,再进行一组样本分布与模型所代表的理论分布间的适合性检验,选择参数显著差异最小的模型为最佳模型。由表1可知,穗上第一叶叶夹角的B-1、E-1、E-3模型和穗下第一叶叶夹角的B-1、E、E-3模型AIC值较小,故为叶夹角的备选遗传模型。根据表2的适合性检验选择统计量达到显著水平个数较少的模型作为最适模型,穗上第一叶叶夹角E-1、E-3模型分别有1、11个统计量达到显著或极显著差异,即分别有1、11个适合性检验统计量描述与分离群体的分布不一致;穗下第一叶叶夹角E、E-3模型分别有1、7个统计量达到显著或极显著差异,即分别有1、7个适合性检验统计量描述与分离群体的分布不一致,故穗上第一叶叶夹角和穗下第一叶叶夹角的最佳遗传模型均为B-1模型,即该性状表现为两对加性-显性-上位性主基因遗传模型。
2.3 糯玉米叶夹角遗传参数估计
由叶夹角最佳遗传模型的各成分分布参数估计出主基因和多基因的一阶遗传参数和二阶遗传参数。从表3一阶遗传参数可以看出,穗上第一叶叶夹角与穗下第一叶叶夹角遗传参数相近,均表现为非加性效应大于加性效应,推测两者可能是受相同基因控制。控制叶夹角的2对主基因的加性效应一正一负,负向效应较大;显性效应较大,且均为正向;2对主基因显性度绝对值均大于1,1对表现为正向超显性,1对表现为负向超显性,说明控制葉夹角的2对主基因的加性效应小于显性效应。加性×加性互作、加性×显性互作、显性×加性互作均为正值,显性×显性互作为负值,上位性效应累计为正向,说明2对等位基因在一起,无论处于纯合还是杂合状态,都将增加叶夹角。
从二阶遗传参数(表3)看,穗上第一叶叶夹角BC1、BC2、F2主基因的遗传率分别为63.96%、72.90%、82.55%,穗下第一叶叶夹角BC1、BC2、F2主基因的遗传率分别63.41%、71.83%、82.01%;F2世代的主基因的遗传率最高,在该世代选择具有较高的选择效率;BC1主基因遗传率小于BC2、F2主基因的遗传率,表明凡是以环江糯作为亲本的回交世代,其主基因遗传率就高于以LN12作为轮回亲本的回交世代的主基因遗传率,推测环江糯中增效主基因效应较大。主基因遗传决定了叶夹角表型变异的 72.42%,还有27.58%的表型变异是由环境因素决定的,说明玉米叶夹角主要受遗传控制,环境因素对其也存在一定影响。
3 讨论与结论
玉米叶夹角是耐密株型育种的重要形态指标,受遗传因素与环境因素的共同影响,属于复杂数量性状。经典数量遗传学多着眼于从群体水平上研究数量性状基因的总体效应,只能检测到性状的加性和显性以及上位性效应,难以从试验中检测出个别基因,本研究所采用主基因-多基因混合遗传模型分离分析方法体系将控制数量性状的基因分为主基因与微基因或多基因2种,打破了原有的数量遗传学的理论假设,通过对分离世代群体的分析为主基因与多基因2个方面的量化分析提供理论依据,更加准确地鉴别出(主)多基因的存在及估算其遗传效应和方差等遗传参数,进而为作物数量性状基因挖掘提供理论依据。
目前,国内外学者关于玉米叶夹角遗传规律作了大量研究,结果不尽相同。前人对叶夹角的经典数量遗传学研究比较深入,结果较为一致,认为加性基因效应最为重要,同时也受非加性基因效应影响[26-34]。邓武明研究表明,虽然不同组合不同部位叶夹角的遗传模型存在差异(穗上、穗位、穗下叶夹角遗传模型A组合为E-1、E-0、E-1,B组合为 B-1、E-1、B-1),各遗传效应也存在较大差异,但均属于2对加性-显性-上位性主基因遗传,总遗传率均在60%以上,不同的是多基因及其上位性效应存在与否[35]。丰光等研究表明,穗上叶夹角符合1对加-显主基因+加-显-上位性多基因遗传模型(即D-0),主基因与多基因的加性效应均大于显性效应,但以非加性效应为主,主基因遗传率为27.55%~52.60%,多基因遗传率为47.40%~72.45%,穗上叶夹角总遗传率较高[36]。马娟等研究指出,穗上叶夹角在春播、夏播不同环境下均符合2对加性-显性-上位性主基因+加性-显性多基因遗传模型(即E-1),而穗下叶夹角在春播、夏播条件下分别符合1对加性主基因+加性-显性多基因遗传模型(即D-2)和加性-显性-上位性多基因遗传模型(即C-0),不同的环境及部位遗传效应及遗传率差异较大,但叶夹角总遗传率均在80%以上[37]。张旷野等研究表明,不同组合穗上叶夹角的遗传模型存在差异(E-1、E-6),主基因遗传率较高[38]。本研究结果表明,叶夹角符合2对加性-显性-上位性主基因遗传模型(即B-1),总遗传率为72.42%,这与丰光等研究结论[36-37]不同,与邓武明的研究结果[35]较一致;本研究中控制叶夹角的非加性效应明显大于加性效应,充分说明了基因间互作效应在叶夹角遗传中发挥着重要作用,与经典数量遗传学研究结论不同。综上所述,不同学者关于叶夹角遗传模型、主基因对数、加性效应、显性效应、上位性效应、遗传率等遗传研究均存在一定的差异,推测可能与研究材料、研究部位、研究方法以及环境的不同有关,说明不同材料叶夹角的遗传机制不同,基因型与环境间存在明显互作效应,前人关于叶夹角不同的QTL定位结果[17,39-41]也说明了这一点。因此,在遗传育种实践中,应根据不同的育种材料,不断通过基因聚合的方法将控制叶夹角的不同基因聚合在一起,同时为避免主基因丢失过早和丢失有利的微效多基因,选育时在早代不宜进行过重的选择,这样可以充分发挥基因间互作效应,降低环境选择误差,准确高效地选育出株型紧凑的育种材料。
参考文献:
[1]于振文. 作物栽培学各论[M]. 北京:中国农业出版社,2003:92-93.
[2]赵久然,王 帅,李 明,等. 玉米育种行业创新现状与发展趋势[J]. 植物遗传资源学报,2018,19(3):435-446.
[3]仇焕广,张世煌,杨 军,等. 中国玉米产业的发展趋势、面临的挑战与政策建议[J]. 中国农业科技导报,2013,15(1):20-24.
[4]赵久然,王荣焕. 中国玉米生产发展历程、存在问题及对策[J]. 中国农业科技导报,2013,15(3):1-6.
[5]Tao Y,Ferrer J L,Ljung K,et al. Rapid synthesis of auxin via a new tryptophan-dependent pathway is required for shade avoidance in plants[J]. Cell,2008,133(1):164-176.
[6]徐庆章,王庆成,牛玉贞,等. 玉米株型与群体光合作用的关系研究[J]. 作物学报,1995,21(4):492-496.
[7]Donald C M. The breeding of crop ideotypes[J]. Euphytica,1968,17(3):385-403.
[8]Duncan W G,Hesketh J D. Net photosynthetic rates,relative leaf growth rates,and leaf numbers of 22 races of maize grown at eight temperatures[J]. Crop Science,1968,8(6):670-674.
[9]Mock J J,Pearce R B. An ideotype of maize[J]. Euphytica,1975,24(3):613-623.
[10]刘绍棣,程绍义,于翠芳. 紧凑型玉米株型及生理特性研究[J]. 华北农学报,1990,5(3):20-27.
[11]胡昌浩,董树亭,岳寿松,等. 高产夏玉米群体光合速率与产量关系的研究[J]. 作物学报,1993,19(1):63-69.
[12]张泽民,贾长柱. 玉米株型对遗传增益的影响[J]. 遗传,1997,19(2):31-34.
[13]彭 勃,张宝石,杨 一,等. 玉米株型育种及其主要性状遗传基础研究进展[J]. 河南农业科学,2007,36(3):14-19.
[14]吕丽华,陶洪斌,王 璞,等. 种植密度对夏玉米碳氮代谢和氮利用率的影响[J]. 作物学报,2008,34(4):718-723.
[15]郑芳英,翟 雨,李宗坤. 玉米株型性状与单株产量的遗传相关和通径分析[J]. 安徽农业科学,2012,40(25):12406-12407,12418.
[16]Vazin F,Hassanzadeh M,Madani A,et al. Modeling light interception and distribution in mixed canopy of common cocklebur(Xanthium stramarium)in competition with corn[J]. Planta Daninha,2010,28(3):455-462.
[17]Tian F,Bradbury P J,Brown P J,et al. Genome-wide association study of leaf architecture in the maize nested association mapping population[J]. Nature Genetics,2011,43(2):159-162.
[18]陳国平,赵久然,李伯航,等. 紧凑型玉米生长发育规律的研究[J]. 玉米科学,1993,1(3):33-38.
[19]李登海. 从事紧凑型玉米育种的回顾与展望[J]. 作物杂志,2000(5):1-5.
[20]宋 碧,吴盛黎,苏銮兵,等. 不同株型玉米高产群体的质量指标[J]. 山地农业生物学报,2001,20(1):1-8.
[21]Stewart D W,Costa C,Dwyer L M,et al. Canopy structure,light interception,and photosynthesis in maize[J]. Agronomy Journal,2003,95(6):1465-1474.
[22]Horton P. Prospects for crop improvement through the genetic manipulation of photosynthesis:morphological and biochemical aspects of light capture[J]. Journal of Experimental Botany,2000,51(增刊):475-485.
[23]盖钧镒,章元明,王建康. 植物数量性状遗传体系[M]. 北京:科学出版社,2003.[HJ2mm]
[24]盖钧镒. 植物数量性状遗传体系的分离分析方法研究[J]. 遗传,2005,27(1):130-136.
[25]曹锡文,刘 兵,章元明. 植物数量性状分离分析Windows软件包SEA的研制[J]. 南京农业大学学报,2013,36(6):1-6.
[26]赖仲铭,杨克诚,雷本鸣,等. 玉米几个自交系株型数量性状遗传的研究[J]. 中国农业科学,1981,14(4):28-36.
[27]王克胜,孔繁玲,杜曼·依马买地. 玉米株型性状的遗传表达和自交系与杂交种株型的聚类分析[J]. 北京农业大学学报,1993,19(3):19-28.
[28]苏书文,白琪林,郭新林. 玉米叶夹角性状配合力的研究[J]. 玉米科学,1993,1(1):1-4.
[29]陈 岭,崔绍平,徐 有,等. 玉米株型性状的遗传分析[J]. 华北农学报,1994,9(增刊1):11-15.
[30]李玉玲,张长江,杨铁柱,等. 玉米株型性状的基因效应研究[J]. 河南农业大学学报,1996,30(1):41-45.
[31]霍仕平,晏庆九,许明陆,等. 玉米主要株型数量性状的基因效应分析[J]. 玉米科学,2001,9(1):12-15.
[32]王秀全,陈光明,刘昌明,等. 玉米株型育种亲本选配的遗传规律研究[J]. 西南农业学报,2000,13(1):50-54.
[33]温海霞,蔡一林,王久光,等. 9个玉米自交系主要株型性状的配合力分析[J]. 西南农业大学学报,2002,24(3):223-225.
[34]蔡一林,王久光,孫海燕,等. 玉米几个株型性状的遗传模型及其与穗粒性状的典型相关分析[J]. 作物学报,2002,28(6):829-834.
[35]邓武明. 玉米株型性状基因效应研究[D]. 雅安:四川农业大学,2001.
[36]丰 光,景希强,李妍妍,等. 玉米穗上叶与主茎夹角性状的数量遗传研究[J]. 玉米科学,2012,20(1):53-56,66.
[37]马 娟,王铁固,张怀胜,等. 玉米叶夹角、叶向值主基因+多基因遗传模型分析[J]. 河南农业科学,2012,41(5):15-19.
[38]张旷野,孙铭泽,闫 伟,等. 玉米穗上叶叶夹角的遗传分析[J]. 作物杂志,2015(6):27-32.
[39]常立国,何坤辉,刘建超,等. 不同环境条件下玉米叶夹角的QTL定位[J]. 玉米科学,2016,24(4):49-55.
[40]张姿丽,刘鹏飞,蒋 锋,等. 基于四交群体的玉米叶夹角和叶向值QTL定位分析[J]. 中国农业大学学报,2014,19(4):7-16.
[41]孙 娇,赵美爱,潘顺祥,等. 玉米叶夹角的全基因组关联分析[J]. 华北农学报,2018,33(1):60-64.
收 稿日期:2019-01-29
基金项目:江苏省农业重大新品种创制项目(编号:PZCZ201711);江苏省农业三新工程(编号:SXGC[2017]097);江苏省连云港市财政专项(编号:QNJJ1807、QNJJ1707);江苏省连云港市“521工程”科研项目(编号:LYG52105-2018019);江苏省第五期“333工程”培养资金。
作者简介:王 军(1982—),男,安徽怀宁人,硕士,副研究员,主要从事玉米遗传育种和栽培技术研究。Tel:(0518)83081707;E-mail:58980883@qq.com。
猜你喜欢
农产品加工(2023年1期)2023-03-21
农村实用技术(2020年4期)2020-12-15
上海农业学报(2017年4期)2017-04-10
现代食品(2016年24期)2016-04-28
上海农业学报(2016年5期)2016-02-10
上海农业学报(2016年5期)2016-02-10
中国蔬菜(2014年11期)2014-02-01
食品科学(2013年17期)2013-03-11
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2010年3期)2010-11-16
