
胞外电子传递机制及其环境效应研究进展
2020-04-29 05:54王红武
四川环境 2020年2期
金 辉, 王红武
(同济大学环境科学与工程学院,上海 200092)
引 言
微生物能通过呼吸作用氧化有机物为自身供给能量,呼吸作用对于微生物的自身生长以及环境中物质的循环都有着十分重要的意义。经典呼吸链理论认为细胞膜是生物电子传递、能量代谢的主要场所。近年来,关于胞外电子传递的研究将经典呼吸链从细胞内延伸到了细胞外,基于此类呼吸链的能量代谢方式即为胞外呼吸[1]。胞外呼吸是指微生物在厌氧条件下在胞内彻底氧化有机物并释放电子,经胞内呼吸链传递到胞外电子受体使其还原并产生能量维持微生物自身生长的过程[2]。可以进行胞外呼吸的微生物被称为胞外呼吸菌,其广泛分布于土壤、河流、海底沉积物中,可分为异化金属还原菌、腐殖质还原菌和产电菌3大类,多数为革兰氏阴性菌,极少数为革兰氏阳性菌[3]。胞外电子传递最早是在20世纪80年代由Lovely等人提出的[4],此后胞外电子传递受到了众多科研工作者的关注,其作为一种新型的细胞呼吸方式,在废水处理、环境修复、能量回收方面都有着较大的应用前景,因此对其传递机理、影响因素和应用前景的研究具有重要现实意义[5-6]。
1 胞外电子传递机理
微生物胞外电子传递主要包括两个过程。第一个过程是电子产生后从内膜经过周质传递到外膜,底物在微生物作用下发生氧化磷酸化,失去的电子通过胞内的NADH脱氢酶、辅酶Q、泛醌等传递至细胞内膜,再通过一系列细胞色素蛋白(Cytochrome,Cyt)或其它功能蛋白传递至细胞外膜[7]。细胞色素蛋白是一类以血红素(heme)为辅基的电子传递蛋白的总称,几乎存在于所有的生物体内。根据吸收光谱的不同,细胞色素可以分为a、b、c三类。第二个过程是电子从细胞外膜传递到最终电子受体,主要有直接接触传递,纳米导线传递机制、电子穿梭机制传递和应电运动四种传递机制[8]。
1.1 电子由内膜传递到外膜
不同产电菌将电子从内膜传递到外膜的方式不同,但细胞色素蛋白尤其是细胞色素c都起着举足轻重的作用。希瓦氏菌(ShewanellaoneidensisMR-1)和地杆菌(Geobactersulfurreducens)是目前对胞外电子传递机制研究最完善的两种胞外呼吸菌。
S.oneidensisMR-1的电子传递机理如图1所示,其细胞膜及周质空间中分布着由多种细胞色素蛋白所构建的电子传递网络[9-10]。CymA氧化位于内膜上的醌类中间体,并将释放的电子传递至位于周质中的细胞色素c。位于外膜上的MtrA、MtrB、MtrC和MtrC的同族蛋白OmcA形成跨外膜蛋白复合物Mtr途径,将电子从周质蛋白传递到细菌外膜表面。最后,由外膜细胞色素MtrC/OmcA完成对胞外电子受体的还原,即直接将电子传递到胞外电子受体上[11]。在这个过程中,CymA起着重要的枢纽作用,SirC和SirD组成的SirCD复合体可作为其替代物[12]。同样的,MtrABC也存在相应的同源物MtrFDE,这两个Mtr同系物模块具有相似的功能。其中,MtrA/D位于周质蛋白中,不能与外膜蛋白直接接触,MtrB/E起到了连接周质和外膜的作用,MtrC/F和OmcA/SirCD按1∶2的比例形成蛋白复合体,可与胞外电子受体直接接触并传递电子[13]。
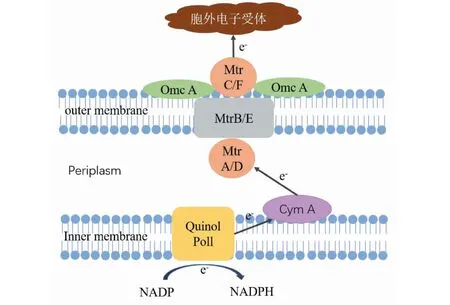
图1 希瓦氏菌(S.oneidensis MR-1)胞外电子传递示意图Fig.1 Schematic diagram of S.oneidensis MR-1 extracellular electron transfer
Geobacter的胞外电子传递模式与S.oneidensisMR-1类似,但Geobacter内各菌株内具有更多的细胞色素c ,且种间差异较大,因而其传递模式更为复杂。G.sulfurreducens位于内膜表面与细胞质相连的MacA对于将电子传递至外膜蛋白至关重要,它可以从醌池中获得电子[14],并还原位于周质中的三亚铁血红素PpcA[15],并将电子传递至外膜蛋白的OMCs(包括OmcB、OmcE、OmcS及OmcZ)[16]。与S.oneidensisMR-1不同的是G.sulfurreducens必须通过导电菌毛将电子传递给胞外电子受体。S.oneidensisMR-1的Mtr金属还原通道中的 MtrABC 蛋白复合体与G.sulfurreducens的孔蛋白-色素蛋白复合体没有同源性,说明它们相似的跨膜电子传递功能是通过独立进化而来的[17]。
1.2 电子从外膜传递到胞外电子受体
胞外呼吸的最终电子受体多为固体或大分子物质,无法进入细胞膜,因此电子需要从细胞膜传递到最终电子受体,由于这个过程发生在细胞外,因此目前的研究多集中于此,其机理也较为清晰。电子由外膜到最终电子受体主要有以下4种机制:直接接触机制、纳米导线机制、电子穿梭机制以及应电运动机制(如图2)[8,17]。
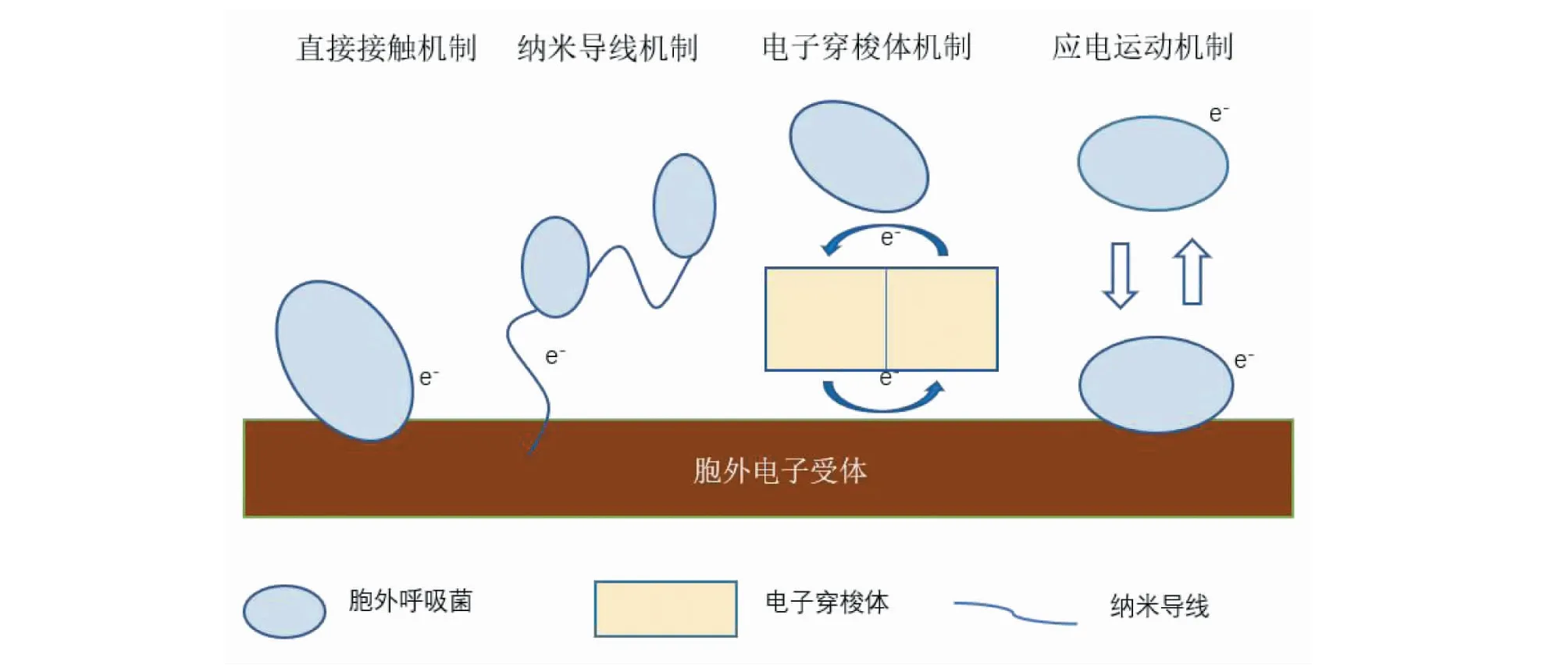
图2 4种胞外电子传递机制示意图Fig.2 Schematic diagram of four kinds of extracellular electron transfer mechanism
直接接触传递是指胞外呼吸菌与电子受体直接接触,通过分布细胞膜外侧的Cyt c将电子直接传递到胞外电子受体表面[18],直接接触传递是最直接的电子传递方式,但是该途径需要胞外呼吸菌紧靠胞外电子受体,限制了其应用且电子传递效率受到细菌浓度以及接触面积的影响[19-20]。
纳米导线最早是在2005年由Reguera等[21]提出的,他们发现胞外呼吸菌G.sulfurreducensDL1的菌毛具有导电性,并把这种生长在细胞周围的类似纤毛且具有导电性能的聚合蛋白微丝命名为“微生物纳米导线”。在电子传递过程中,纳米导线如同电子导管一样,将细胞膜与电子受体连接起来。由于纳米导线的存在,胞外呼吸菌可以在不接触受体的情况下,实现电子远距离转移,并且大大提高电子传递效率[14,22]。
微生物可以利用外源介体(如环境中存在的腐殖质、植物根系分泌物等)或内生介体(自身分泌的代谢产物,如核黄素等),将电子转移至电子受体,这种通过中介体进行胞外电子传递的方式即为电子穿梭机制,作为中介体的外源或内生介体叫做电子穿梭体(electron shuttle,ES),也称为氧化还原介体(redox mediate,RM)[23]。电子穿梭体是一种特殊的电子载体,可以可逆地参与氧化还原反应。电子穿梭体可以穿过细胞膜,在细胞内被还原并将电子传递到胞外电子受体,在受体表面被氧化,如此循环往复,实现电子的跨膜转移。
应电运动机制是指某些胞外呼吸菌可以将产生的电子储存在细胞膜表面,然后利用鞭毛运动撞击受体表面,在撞击瞬间释放电子,完成电子转移。在接触释放电子后,从胞外电子受体表面脱离,参与下一个循环[24]。
2 胞外电子传递的影响因素
胞外电子传递发展潜力好,随着对其传递机理研究的不断深入,人们开始关注影响胞外电子传递效率的因素,以及可以促进电子传递的措施。深入研究胞外电子传递机理以及促进措施对胞外电子传递在各个领域的应用有着十分重要的意义。
2.1 细胞色素c对胞外电子传递的影响
细胞色素c对胞外电子传递起到至关重要的作用。细胞色素c通常含有多个血红素(heme),电子可以在相邻血红素的铁原子之间进行转移,使得细胞色素c可以传递电子[25]。理论计算发现,细胞色素c中血红素的距离和排列方式会影响电子传递的能力和效率[26],因此,细胞色素c对电子的传递能力与其分子结构密切相关。韩蕊[9]对S.oneidensisMR-1野生型和各种突变体对水铁矿的还原及生物矿化作用进行了研究,结果表明MR-1主要通过胞外电子传递将水铁矿还原呈Fe2+,并且敲除MtrA、CymA等关键系胞色素c后,MR-1的电子传输能力大幅降低,对水铁矿的还原速率和转化程度也降低,说明MtrA、CymA是S.oneidensisMR-1胞外电子传递链的重要影响因素。丁德武[27]通过构建基因组尺度的S.oneidensisMR-1细胞色素c网络,发现细胞色素c网络的独特结构可能衍生出S.oneidensisMR-1产生电力的能力,并且可以从该细胞色素c网络中识别潜在的EET途径。Li Hui等人[28]将含氧官能团引入微生物燃料电池(Microbial fuel cells,MFCs)阳极后发现该MFCs的最大电荷密度增加了27.7%,阳极电阻则降低了21.8%,研究表明,含氧基团的存在并没有改变MFCs中的微生物群落,但其可以进行胞外电子传递的电化学活性菌(Electrochemically Active Bacteria,EAB)细胞表面的细胞色素c比对照组增加了4倍,这说明细胞色素c含量的增加可以增强EAB和电极之间的胞外电子转移。
2.2 解偶联剂对胞外电子传递的影响
胞外电子传递过程十分复杂,底物在氧化的过程中会在细胞质膜两侧产生质子推动力(proton motive force,PMF),为ADP磷酸化形成ATP提供能量。底物的氧化会生成大量电子和质子,并在膜外侧累积。在一定浓度范围,利用解偶联剂分离底物氧化过程和ATP合成过程,膜外侧质子的累积及产物抑制被破坏,包括S.oneidensisMR-1在内的六种纯种微生物胞外电子传递过程得到促进[29]。
虞盛松[30]通过考察不同浓度的解偶联剂3,3’,4’,5-四氯水杨酰苯胺(tetrachlorosalicylanilide, TCS)对Shewanella胞外电子传递的调控发现低浓度(50 g/L)的TCS可以在促进MR-1对乳酸的消耗同时提高Shewanella的产电能力;而高浓度(400 g/L)的TCS会通过降低ATP合成酶的活性来抑制Shewanella的代谢和产电。这些结果表明,适当添加解偶联剂是一种简便的胞外电子传递调控手段。
2.3 铁氧化物对胞外电子传递的影响
铁氧化物表面与微生物胞外电子传递链上的细胞色素c之间的相互作用决定了铁氧化物-微生物界面电子转移的效率。理论计算结果表明胞外电子传递过程中不同蛋白之间的电子传递速率远远大于外膜蛋白和铁矿物界面的电子传递速率,因此微生物胞外电子传递的决速步骤是外膜蛋白与铁矿物表面之间的电子转移过程[31]。磁铁矿能够促进铁还原微生物的胞外电子传递,在水田沉积物中加入磁铁矿能在Geobacter富足的情况下加速电子传递到电极上的过程[32]。也有研究发现,磁铁矿能够取代Geobacter胞外电子传递过程中菌毛的作用,促进地杆菌的长距离电子传递[33]。纳米磁铁矿能像细胞色素c一样附着在Geobacter的菌毛上,从而促进胞外电子传递,且可以代替OmcS完成胞外电子传递[34]。近期天然赤铁矿也被验证能够有效参与并促进微生物胞外电子传递[35]。
2.4 电子穿梭体对胞外电子传递的影响
在间接胞外电子传递过程中,微生物可以依靠自身分泌的内生介体或外界介体作为电子穿梭体,完成电子的跨膜传递。外加电子穿梭体(如腐殖质、黄素等)的存在对胞外电子传递具有促进作用。在ShewanelladecolorationisS12和AeromonashydrophilaHS01还原针铁矿(α-FeOOH)的过程中,添加AQC可使针铁矿的还原速率提高3~4倍的[36-37];在G.sulfurreducens还原水铁矿的过程中,添加溶解态腐殖质可使水铁矿还原速率提升6倍[38];在利用醌类物质2-羟基-1,4-萘酯LQ还原Se(Ⅳ)、Te(Ⅳ)的过程中,腐殖质类物质的加入能够有效增强二者的还原效果[39];在Bacillussp.BT1还原Cr(Ⅵ) 的过程中,AQS的存在可以提高Cr(Ⅵ)的还原效率[40];在利用S.oneidensisMR-1还原Fe(Ⅲ)氧化物的研究中,加入10μM/L的黄素类物质,Fe(Ⅲ)氧化物的还原速率可提供18倍[23]。
3 胞外电子传递的应用
胞外电子传递由于其电子受体的存在于细胞外的这一特殊性,因而在污水处理、污染物转化降解和生物能源的清洁生产中有着广阔的应用前景。
3.1 基于微生物燃料电池的应用
微生物燃料电池(MFCs),是以微生物为催化剂将可生物利用有机质中化学能直接转化为电能的燃料电池系统。MFCs以胞外电子传递链为基础,利用胞外电极接收胞外呼吸菌产生的电子,转化为电能。在MFCs中,微生物在阳极催化氧化有机污染物并产生质子和电子,质子通过基质和质子交换膜(Proton exchange membrane ,PEM)传递到阴极,与氧气和电子结合生成水;微生物通过呼吸作用产生的电子首先通过胞内电子传递链传递到细胞膜,然后通过纳米导线或电子穿梭体等直接或间接的方式传递到阳极表面再通过外电路传递到阴极,在阴极与电子受体发生还原反应,完成整个氧化还原反应,同时电子不断流过外电路而产生电能[41]MFC系统组成示意图如图3所示。自1991年MFCs第一次被用于处理污水,微生物燃料电池电池掀起了胞外电子传递的应用热潮,也是目前胞外电子传递研究最多,应用最广泛的领域[42]。
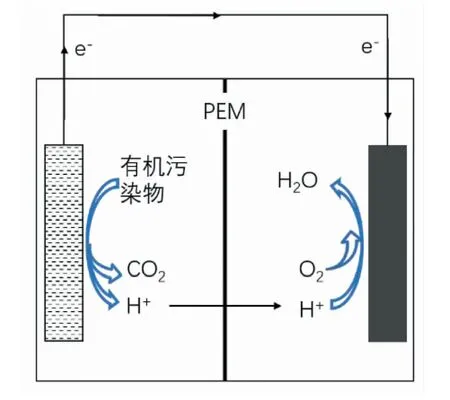
图3 MFC示意图Fig.3 Schematic diagram of microbial fuel cell
目前MFCs在微生物产电,易降解有机物、难降解有机物降解,生物脱氮、生物脱硫等领域有较多研究[41,43~47]。
废水中含有大量的有机物,这些物质可以作为碳源和电子供体被阳极微生物利用,在去除污染物的同时还可以进行能量回收。Li[48]利用单室空气阴极MFC系统处理某食堂废水,研究表明在进水COD为2 700±20mg/L时,MFC的最大功率密度可达5.6W/m3,平均输出电压为0.51V,并且COD去除率可高达90.3%。Guo[49]利用双室MFC处理炼油废水时发现COD和含油量分别为450±50mg/L和50±5mg/L时,可分别去除64±4%的COD和84±3的含油量,此时MFC的最大功率密度为0.33 W/m3。Miran[45]处理某含铜量为20mg/L的废水时发现,双室MFC对Cu2+的还原效率可超过98%,同时TOC的去除率也可超过90%。
3.2 基于污染物降解的应用
胞外电子传递也可应用于环境修复,在降解有机污染物、氧化还原有毒有害重金属污染物等方面起到了重要作用。微生物利用胞外电子传递降解污染物主要通过两种机理:一是以有机污染物为电子供体,降解有机物;二是以污染物为电子受体,将有毒有害的的重金属污染物转化为低毒或无毒状态[5]。
人类活动产生了大量有机污染物,有机物电子供体的异化作用会把 O2作为电子受体而迅速消耗掉,从而形成厌氧环境,为 Fe(Ⅲ)氧化物作为电子受体参与有机物的氧化提供了主要场所,使得微生物通过胞外电子传递降解有机物成为可能。少数地杆菌可以在还原Fe(Ⅲ)氧化物的同时氧化芳香族化合物并降解一些微生物无法直接利用的共存污染物,这对修复芳烃污染的土壤有重大意义[50]。
以金属为最终电子受体的金属还原菌,可以还原污水中的有价金属污染物,将有毒金属污染物转化为无毒或低毒物质。此外对于Cr(Ⅵ),Se(Ⅳ),Se(Ⅵ),Tc(Ⅶ)和U(Ⅵ)等以溶解态存在金属元素,以及Cr(Ⅲ),Se(0),Tc(Ⅳ)和U(Ⅵ)等不溶态存在的金属元素也可以利用金属还原菌进行修复[51]。如脱硫弧菌(Desulfovibriospp.)可以以H2为电子供体利用细胞色素c还原Cr(Ⅵ)[52]。在生物浸出方面,氧化亚硫杆菌在低品位的铜矿、金矿、铀矿等领域取得了良好的浸出效果[53]。生物浸出同样是利用胞外电子传递,将非溶解态有价金属转变为溶解态离子,并加以回收,在矿产资源的回用中应用广泛。除了金属离子污染物,铁还原菌可以降解水中的有机污染物[54]。对于一些难降解有机污染物,如石油烃、多氯联苯和硝基苯等污染物的去除效果也较好[55]。在生物脱盐领域,以胞外电子传递为基础的微生物脱盐电池(Microbial desalination cell,MDC),能够利用产电菌去除溶液中的部分阴、阳离子,从而起到较好的脱盐效果[56]。
微生物胞外电子传递可以产生含胞外电子受体的纳米材料,对Fe(Ⅲ),Pd(Ⅱ),Se(Ⅳ),Se(Ⅵ)等的胞外还原可以形成含Fe(Ⅱ),Pd(0),Se(0)等的纳米材料。这些纳米材料在催化,修复环境等方面都有着重要作用[4,57]。
4 结语与展望
胞外呼吸丰富了微生物代谢的多样性,在微生物燃料电池、污水处理、环境修复和纳米材料合成等环境领域有着良好的应用前景,因此,微生物胞外电子传递的研究将受到持续的关注。目前主要针对两种产电菌(S.oneidensis MR-1和G.sulfurreducens)开展了电子从胞内传递到细胞外膜的研究,对其他产电菌的研究很少;而随着部分微生物胞外电子传递过程中至关重要的细胞色素蛋白得到确定,电子传递途径逐渐清晰;人们已发现通过调控电子从外膜到胞外电子受体的过程可以影响电子传递的效率。但是,目前胞外电子传递的研究还有很大的局限性,大多数的研究针对单一菌种,如集中在S.oneidensis和G.sulfurreducens这两种产电菌上,亟待对其他菌种以及混合菌群开展研究,且胞外电子传递的分子学机制需要进一步完善。此外,在研究传递机理的同时,也亟需开展胞外电子传递的影响因素以及促进措施的研究,以提高产电效率及处理污染物的效果等。”
猜你喜欢
娃娃乐园·综合智能(2022年9期)2022-08-16
当代化工研究(2022年9期)2022-05-28
昆明医科大学学报(2021年4期)2021-07-23
科学大众(2021年9期)2021-07-16
食品科学(2020年17期)2020-09-21
食品与发酵工业(2019年21期)2020-01-13
中国比较医学杂志(2019年9期)2019-01-08
心肺血管病杂志(2018年11期)2018-12-18
中国调味品(2017年2期)2017-03-20
中国病理生理杂志(2015年10期)2015-01-26
