
马尾松PmPIN1基因的克隆及功能分析*
2020-04-28 07:10胡兴峰陈佩珍孙晓波季孔庶
林业科学 2020年3期
武 星 胡兴峰 陈佩珍 孙晓波 吴 帆 季孔庶
(南京林业大学 林木遗传与生物技术省部共建教育部重点实验室 南方现代林业协同创新中心 南京210037)
生长素作为植物体内一种重要的植物激素,对于植物生根有重要作用,其通过极性运输方式调控植物的生长发育。目前研究表明,生长素的极性运输在根中主要有2种表现形式: 一是沿中柱细胞向根尖的向顶式运输; 二是通过皮层细胞及表皮由根尖流向根基的向基式运输,随后,通过伸长区表皮层流回中柱细胞,形成循环回流(Kepinskietal., 2005; 李俊华等, 2006)。在拟南芥(Arabidopsisthaliana)中的研究验证,PIN-FORMED(PIN)蛋白家族是极性运输中重要的输出载体(Gälweileretal.,1998)。PIN1是研究较为广泛的成员之一,在生长素由茎尖流向根尖、维管束分化及不定根的形成过程中至关重要(Frimletal., 2003),分布于木质部薄壁细胞、维管束细胞和茎尖分生组织的细胞膜(Vietenetal., 2005; Paponovetal., 2005; Blilouetal., 2005)。在拟南芥的pin1突变体中,生长素极性运输效率下降(Sabatinietal.,1999)。pin1突变体有根去向重力性、侧根发生异常等表型(Frimletal., 2002a; Maheretal.,1980),与生长素极性运输抑制剂NPA处理后的植株表型一致(Frimletal., 2002b)。
作为我国南方特有的重要树种马尾松(Pinusmassoniana),其播种苗主根性极强而侧根数少且不发达,这在一定程度上影响了苗木的质量,最终会影响到造林成效; 同时其无性繁殖时生根难,造成其无性育林举步维艰,阻碍了良种选育获取更高增益的进程。可见为了从根本上解决马尾松的生根问题,有必要研究生长素相关的基因。本研究利用PCR与RACE技术成功克隆了马尾松PmPIN1基因cDNA全长,并通过在线软件对PmPIN1基因编码的蛋白进行生物信息学分析,利用实时荧光定量技术分析PmPIN1基因在马尾松不同组织中的表达差异; 其次,利用农杆菌(Agrobacteriumrhizogenes)介导法将PmPIN1基因导入至烟草(Nicotianatabacum)中,并对其进行功能分析。本研究将有助于在分子水平上揭示马尾松的生根机制和探寻有效的促根方法。
1 材料与方法
1.1 试验材料及试剂 试验材料为实验室30天生马尾松实生幼苗、南京林业大学校园内10年生马尾松及实验室30天生拟南芥实生幼苗。“RNAprep Pure多糖多酚植物总RNA提取试剂盒”及“新型植物基因组DNA提取试剂盒”均购自天根生物有限公司; 反转录试剂盒及同源重组试剂盒购自诺威赞生物有限公司; “Plasmid Maxi Kit”(质粒大提试剂盒)购自QIAGEN公司; “植物原生质体制备及转化试剂盒”购自中科瑞泰生物有限公司; pEASY-T1、pEASY-Blunt载体、大肠杆菌感受态购自全式金生物有限公司; 16318-hGFP载体购自西安淳风生物科技有限公司; 高保真GXL酶及NcoⅠ、SalⅠ、BamHⅠ限制性核酸内切酶、RACE试剂盒均购自Takara生物有限公司; EHA105农杆菌感受态购自上海唯地生物技术有限公司;pCAMBIA-1302真核表达载体由实验室保存; PEG4000购自Sigma公司; 纤维素酶、离析酶购自Yakult公司; PBS缓冲液购自上海捷瑞生物技术有限公司; DAPI、CM-DiI染色剂购自上海翊圣生物科技有限公司。
1.2 马尾松PmPIN1基因的克隆 在NCBI网站查询PIN1基因,并与实验室的马尾松转录组数据(National Omics Data Encyclopedia: OEZ004657)比对,得到马尾松PmPIN1基因核苷酸序列。利用“RNAprep Pure多糖多酚植物总RNA提取试剂盒”提取马尾松幼苗全株的RNA,提取步骤参照其说明书,琼脂糖凝胶电泳检测RNA完整性,NanoDrop超微量分光光度计测定RNA浓度与OD值,为保证RNA质量,试验器具均经高温处理,塑料器材为一次性无菌无酶,试验操作过程避免RNA酶对其影响。用反转录试剂盒反转得到cDNA模板。根据PmPIN1序列,利用Primer5.0设计特异性引物F1、R1、3′RACE Primer、5′RACE Primer,并以cDNA为模板,对马尾松PmPIN1基因进行PCR扩增,切胶回收目的条带,将目的片段连接至pEASY-T1或pEASY-Blunt载体上,转化至大肠杆菌感受态中,将阳性克隆送去测序,利用DNAMAN软件将测序片段进行拼接获得基因全长。引物序列见表1。利用在线软件TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)分析PmPIN1蛋白的亲水性,利用生物信息学软件MEGA5.1对不同物种的PIN1氨基酸序列构建系统进化树。
1.3PmPIN1基因的实时荧光定量分析 利用高枝剪剪取10年生马尾松1年生枝条、新叶、花,挖取其新生幼根,迅速清水冲洗干净,滤纸吸净表面水分后液氮冻存。利用“RNAprep Pure多糖多酚植物总RNA提取试剂盒”分别提取幼根、1年生枝条、新叶、花的RNA,用反转录试剂盒反转获得cDNA模板。以Actin2为内参基因,利用在线软件Primer3.0设计特异性引物F2、R2,Actin2-F、Actin2-R,并以 cDNA为模板,进行实时荧光定量PCR反应。生物学重复为3次,试验重复为5次; 根据公式ΔΔCt=ΔCt(实验)-ΔCt(对照)及RQ=2-ΔΔCt计算出PmPIN1在不同组织中的表达量,利用SPSS软件分析显著性差异,用Origin8.0软件作图。
1.4 真核表达载体的构建及转基因苗鉴定 利用在线软件CE-Design设计含NcoⅠ酶切位点的引物pCAMBIA-1302-PmPIN1F、pCAMBIA-1302-PmPIN1R,为使目的片段与GFP连接后表达顺利,设计引物时加保护碱基T,以cDNA为模板,克隆了含有酶切位点的目的片段; 并用NcoⅠ内切酶单酶切切割pCAMBIA-1302载体; 连接得到真核表达载体pCAMBIA-1302-PmPIN1,将其转化至农杆菌EHA105中,摇菌至OD≈0.6时,利用叶盘法侵染至继代培养的野生型烟草中,通过分化、壮芽、生根培养后,移栽至营养土中,用转pCAMBIA-1302空载烟草作为表型观察对照。利用“新型植物基因组DNA提取试剂盒”提取转基因与野生型烟草的DNA,具体步骤参照其说明书,设计特异性引物F3、R3进行分子水平上检测; 利用激光共聚焦显微镜观察转基因烟草根中PmPIN1-GFP蛋白的表达。
1.5 生长素含量测定 利用酶联免疫法测定转基因及野生型烟草不同部位的生长素(IAA)含量; 将等量不同浓度的NPA(1-N-萘基邻氨甲酰苯甲酸)(0、0.4、2、10 nmol·L-1)溶液注射在3 cm×3 cm大小脱脂棉上,将其包裹于转基因与野生型烟草的根茎结合处,另用保鲜膜缠绕,试验重复3次。10天后取下脱脂棉,利用酶联免疫法测定烟草根、根茎结合处、叶中的生长素含量。
1.6 亚细胞定位分析 将30天生拟南芥叶片切成1 mm宽小条,利用“植物原生质体制备及转化试剂盒”获得拟南芥原生质体,具体步骤参照其说明书。用SalⅠ、BamHⅠ内切酶线性化16318-hGFP; 利用CE-Design设计引物16318-hGFP-PmPIN1F、16318-hGFP-PmPIN1R(表1),以cDNA为模板,PCR获得含SalⅠ、BamHⅠ酶切位点的目的片段,连接获得16318-hGFP-PmPIN1重组载体(图1),转化至大肠杆菌(Escherichiacoli),经菌检、测序,从公司返还测序正确的大肠杆菌菌液,充分摇菌,利用“Plasmid Maxi Kit”获得高质量重组质粒,具体步骤参照其说明书。利用PEG介导法将16318-hGFP-PmPIN1重组载体转化至拟南芥原生质体,转化结束后孵育16 h,DAPI、CM-DiI染色剂侵染10 min,室温环境PBS清洗2次,每次5 min,然后用激光共聚焦显微镜观察荧光信号,进行亚细胞定位分析。为减少细胞破碎,转化过程尽量轻柔。
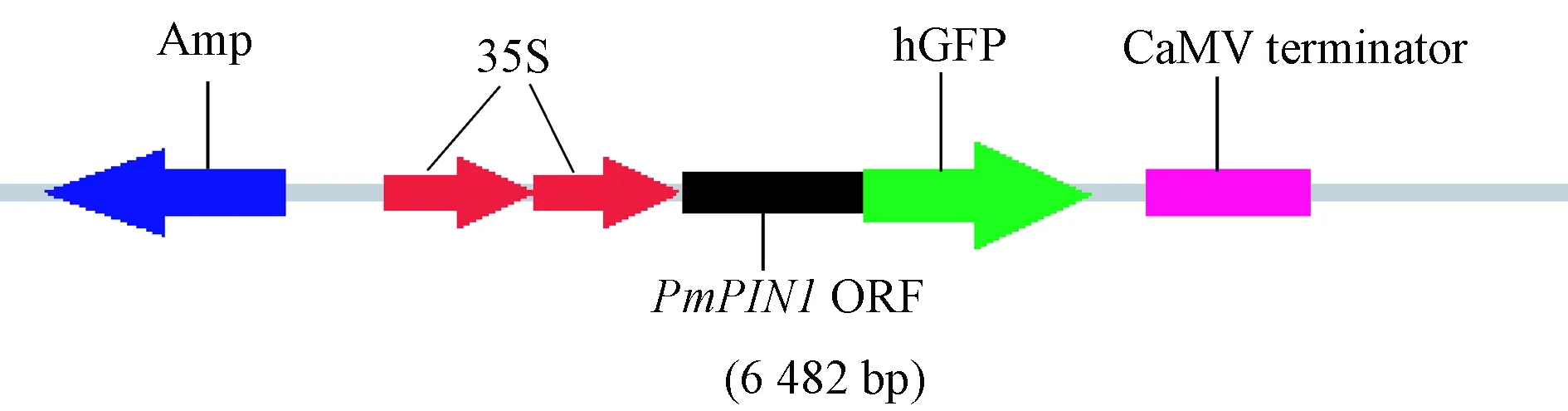
图1 16318-hGFP-PmPIN1融合表达载体示意Fig.1 Schematic diagram of 16318-hGFP-PmPIN1 fusion expression vector

表1 PCR所用引物Tab.1 Primer sequence used in PCR

图2 PmPIN1蛋白跨膜预测Fig.2 The prediction of transmembrane of PmPIN1 proteins峰值代表跨膜结构域。The red peak represents the transmembrane domain.
2 结果与分析
2.1 马尾松PmPIN1基因全长的克隆 利用PCR与RACE 技术从马尾松中克隆出PmPIN1基因全长cDNA,长度为2 914 bp,ORF为2 085 bp,其编码蛋白的分子量是76.32 kDa,等电点(pI)是9.27。在线软件TMHMM对PmPIN1蛋白的亲水性进行分析,该蛋白是亲疏性相间蛋白,中间以亲水性为主(图2),并且N端与C端均有膜运输蛋白,由此推测PmPIN1蛋白是膜蛋白。
2.2PmPIN1基因编码氨基酸序列分析 利用NCBI中的Blastp比对PmPIN1基因编码的氨基酸序列,结果显示,马尾松PmPIN1与油松(Pinustabulaeformis)相似度为99%,与花旗松(Pseudotsugamenziesii)、金松(Sciadopitysverticillata)的相似度分别为68%、66%。根据PmPIN1基因编码氨基酸序列的比对结果,选取多个物种的PIN1基因编码的氨基酸序列构建系统发育进化树(图3)。结果显示,马尾松与油松在同一支上,说明目前马尾松PmPIN1与油松PIN1亲缘关系最近,并且马尾松与水松(Glyptostrobuspensilis)、金松、罗汉柏(Thujopsisdolabrata)亲缘关系较近。PIN1基因在裸子植物与被子植物中均存在,进一步说明PIN1基因进化上的保守性。

图3 PmPIN1与其他物种PIN1基因系统发育树的分析Fig.3 The analysis of phylogenetic tree based on amino acid sequences of PmPIN1 gene from Pinus massoniana and other plants
2.3PmPIN1基因的实时荧光定量分析 2018年4月采取10年生马尾松幼根、1年生枝、新叶、花4个不同组织样品,并在4个组织中做PmPIN1基因的实时荧光定量分析试验(图4)。PmPIN1基因在幼根、1年生枝、新叶、花中的表达量差异达到极显著,在1年生枝中表达量最高,幼根中次之,约是在1年生枝中的0.75倍,在花中表达量最低。

图4 PmPIN1基因组织特异性表达Fig.4 The expression quantity of PmPIN1 gene in different tissues of Pinus massoniana不同小写字母表示显著差异(P﹤0.05)。Different lowercase letters indicate significant difference (P<0.05).
2.4pCAMBIA-1302-PmPIN1真核表达载体的构建与转基因烟草的检测 1)pCAMBIA-1302-PmPIN1真核表达载体的构建 选取pCAMBIA-1302载体上NocⅠ酶切位点,利用CE Design V1.04软件设计含有NocⅠ酶切位点的特异性引物,扩增含有NocⅠ酶切位点PmPIN1的ORF目的片段与线性化载体。切胶回收目的片段与单酶切载体,利用DNA连接酶连接构建出pCAMBIA-1302-PmPIN1真核表达载体(图5)。
2) 转基因烟草分子水平上检测 组培苗移栽至营养土中约30天左右时,提取转基因与野生型烟草的DNA,选取pCAMBIA-1302载体与PmPIN1的连接序列,利用Primer3.0设计特异性引物,PCR反应检测后,得到32株转基因植株,转基因烟草扩增出目的条带462 bp,野生型则没有(图6)。
3) 转基因与转pCAMBIA-1302空载烟草表型对照 分别移栽T0代转pCAMBIA-1302-PmPIN1烟草与转pCAMBIA-1302烟草组培苗,约20天时,观察二者的表型差异,转基因烟草较转空载植株高大,节间增长(图7); 转空载植株发根量较少,且根长不及转基因植株(图8)。
4) 转基因烟草GFP荧光检测 利用激光共聚焦荧光显微镜观察转基因及野生型烟草根部的PmPIN1-GFP荧光信号。发现野生型有微弱的自发荧光,而转基因烟草根部中间部位有明显的荧光富集现象,且在细胞周围表达明显(图9)。

图5 PmPIN1-GFP融合蛋白表达载体示意Fig.5 Schematic diagram of PmPIN1-GFP fusion protein expression vector

图6 转基因及野生型烟草基因组PCR检测Fig.6 Genome PCR identification of transgenic and wild tobacco 1-2: 野生型; 3-10: 转基因。1-2: Wild;3-10: Transgenic.

图7 转基因与转空质粒烟草20天的长势Fig.7 The growth of transgenic and transgenic plasmid tobacco after 20 days

图8 转基因与转空质粒烟草株高与根系对照Fig.8 Comparison between plant height and root system of transgenic and transgenic plasmids tobacco

图9 转化型与野生型烟草的GFP荧光观察Fig.9 GFP fluorescence detection of transgenic and wild tobaccoA: pCAMBIA-1302-PmPIN1; B: pCAMBIA-1302-PmPIN1根部分生区细胞 Cells in the root meristem zone.
2.5 生长素含量分析 1) 转基因与野生型烟草生长素含量测定 IAA酶联免疫测定转基因与野生型烟草根、根茎结合处、叶不同部位的生长素含量(表2)。结果表明,转基因烟草与野生型不同组织中生长素含量均未达到极显著差异,转基因烟草的根、根茎结合处的生长素含量高于野生型,叶中相反。
2) 不同浓度NPA处理后生长素含量变化 不同浓度NPA处理转基因与野生型烟草的根茎结合处,测定不同部位的生长素含量差异(表2)。测定结果显示,不同浓度NPA处理后,转基因烟草根、根茎结合处、叶中生长素含量变化均未达到极显著。而野生型烟草根中生长素含量变化达到极显著,且随NPA浓度增加,生长素含量减少; 处理浓度为2 nmol·L-1时,根茎结合处生长素含量最大; 处理浓度为10 nmol·L-1时,叶中生长素含量增加最多。
2.6 亚细胞定位分析 利用PEG介导法将16318-hGFP-PmPIN1重组载体转化至拟南芥原生质体中进行亚细胞定位分析(图10)。亚细胞定位分析表明PmPIN1蛋白主要分布在细胞膜上。
3 讨论
在马尾松人工林营建过程中,由于其主根性强、须根少,移栽时易伤根系,影响到造林成活率,前人通过截根的方式使主根变短且增加须根与菌根量,减少运输过程中的窝根现象,利于提高造林成活率(伍家荣等, 1994),但未在根本上解决问题。此外,马尾松无性繁殖可解决无性系育林问题,从而加快其育种进程,且可保留优良的遗传性状,最大程度地获取遗传增益。但生根困难一直制约了其进展,也影响到该树种遗传转化体系的构建,阻碍了其分子育种的开展。可见从根本上解决马尾松生根问题极其迫切。一般认为,最适激素种类和浓度对植物根系的发育起促进作用。在研究马尾松扦插生根试验中,采用自制的复配型生根调节剂,获得显著的生根效果(季孔庶等, 2001)。在组培快繁过程中用0.2 mg·L-1NAA处理马尾松无性系,其中GLM-80无性系生根效果最佳,且移栽成活率较高(姚瑞玲等, 2016)。分子水平的研究可从源头上分析根系发育过程,为从根本上解决马尾松实生苗根系不够发达问题,并攻克其组培不易生根及难以搭建转基因平台等难题提供理论基础。已有涉及马尾松生根相关基因的研究表明,PmTIR1在幼嫩马尾松根中表达量较高(苏江等, 2015),PmSHR、PmSCR、PmPIN2基因在成年马尾松根中表达量最高(朱沛煌, 2016; 武星等, 2018)。本研究发现PmPIN1基因在10年生马尾松1年生枝与幼根中均较高,PmPIN1-GFP荧光蛋白在转基因烟草根部中间部位有明显的荧光富集,PmPIN1转基因烟草呈现发根量增加的表型,说明PmPIN1基因可能参与根系发育过程。

表2 NPA对 PmPIN1转基因与野生型烟草生长素含量的影响①Tab.2 The effect of NPA on the content of auxin in PmPIN1 transformed and wild tobacco
①表中数据为3个重复的平均值±标准误,同列数据标有不同字母表示有显著差异(P<0.05)。The data in the table is mean value±standard error of 3 replicates, and the same column of data marked with different letters indicates significant difference (P<0.05).
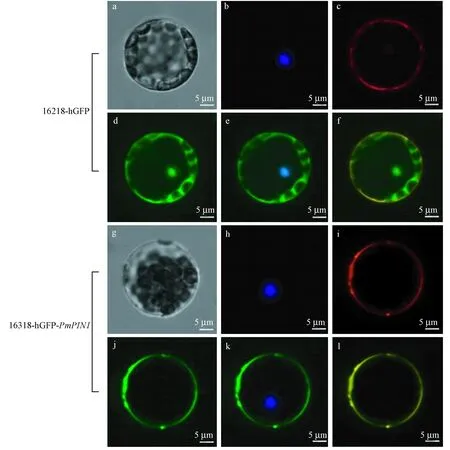
图10 PmPIN1蛋白亚细胞定位Fig.10 Subcellular localization map of PmPIN1 proteinsa,g: 明场; b,h: DAPI染核; c, i: CM-DiI染膜; d,j: GFP; e,k: GFP+DAPI; f,l: GFP+CM-DiI。a,g: Bright; b,h: DAPI stained cell nucleus; c,i: CM-DiI stained cell membrane; d,j: GFP; e,k: GFP+DAPI; f,l: GFP+CM-DiI.
生长素在植物根系形态建成过程中至关重要。早期的研究表明,生长素可通过增加侧根、不定根量影响根系发育(Woodwardetal., 2005)。生长素是一种具有极性运输性质的植物激素,生长素的极性运输是指其流动方向不受重力作用,从形态学上端流向下端(倪为民等, 2000)。PIN蛋白是一类公认的生长素输出载体,在生长素由细胞内向外运输的过程中至关重要。研究发现,pin突变体有生长素运输异常的表型(Gälweileretal.,1998; Chenetal., 1998); PIN蛋白的时空表达与生长素的运输方向一致(Peeretal., 2004); PIN蛋白运输形成的生长素浓度梯度在侧根发育过程中起关键作用(Benkováetal., 2003)。在生长素的极性运输过程中,NPA结合蛋白(NBP)受肌动蛋白的协助,影响输出载体PIN蛋白的分布与运输(Mudayetal., 2002),NPA是一种常见的生长素运输抑制剂,其通过与运输载体结合的方式改变蛋白质的空间结构,进而影响载体与生长素的结合位点。芽孢杆菌(Bacillus)侵染拟南芥,表现出主根伸长、侧根量增多,8 μmol·L-1的NPA处理可解除芽孢杆菌的促进作用(Pérez-Floresetal., 2017); NPA处理野生型水稻(Oryzasativa),根中生长素响应报告基因DR5∷GUS在根尖集中表达,在中柱没有蓝色信号,NPA处理OsPIN2超表达水稻时,蓝色信号分布没有发生变化(陈赢男, 2012)。本研究通过酶联免疫(IAA)测定发现转化PmPIN1烟草的根和根茎结合处的生长素含量高于叶; NPA处理野生型烟草,不同组织生长素含量变化明显,且根中出现极显著变化; NPA处理PmPIN1转基因烟草,不同组织生长素变化均不明显,说明PmPIN1转基因烟草可减少NPA对植物体内生长素分布的影响,表明PmPIN1调控生长素的极性运输,此结果与PIN家族的研究结果(陈赢男, 2012)一致。 PIN1蛋白结构研究表明,其由中间较长亲水区域与两端疏水区域构成,定位在细胞膜上(Zazímalováetal., 2007)。菊花(Chrysanthemummorifolium)CmPIN1蛋白的跨膜预测表明其N端与C端为跨膜区,中间为亲水区; 亚细胞定位表明其定位于细胞膜上,属于膜蛋白(吕素慧等, 2016)。水稻OsPIN1(Xu, 2005)与拟南芥AtPIN1(Paponovetal., 2005; Kreceketal., 2009)中均有相同结论。本研究预测PmPIN1蛋白的跨膜结构,并对其进行原生质体亚细胞定位分析,得出PmPIN1是膜蛋白,说明PIN蛋白在不同物种中具有保守性。
4 结论
本研究克隆了马尾松PmPIN1基因,基因全长2 914 bp,包含2 085 bp的ORF。PmPIN1蛋白N端与C端都存在膜运输蛋白,预测PmPIN1属于膜蛋白,与亚细胞定位结果一致。PmPIN1在被子植物与裸子植物中均存在,表明PmPIN1基因进化上的保守性。PmPIN1基因在幼根与1年生枝条中表达量较高,进一步验证PIN1蛋白参与维管束及侧根形成的过程。转基因烟草根、根茎结合处生长素含量高于野生型,叶中相反; 不同浓度NPA对烟草根茎结合处处理时,野生型烟草不同部位的生长素含量均随NPA的浓度变化而发生明显变化,转基因烟草无明显变化。转基因烟草生长势优于转pCAMBIA-1302空载植株,且发根量增加; PmPIN1-GFP绿色荧光蛋白富集在转基因烟草根部中间部位。综上可见,PmPIN1在根生长发育过程中起一定作用。其具体作用将有待进一步的深入分析。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
学与玩(2022年10期)2022-11-23
今日农业(2022年3期)2022-06-05
生物学通报(2021年4期)2021-03-16
科学大众(2020年12期)2020-08-13
绿色科技(2019年5期)2019-11-29
乡村科技(2019年9期)2019-06-26
数码世界(2018年7期)2018-12-24
中国粮油学报(2018年12期)2018-03-19
创新科技(2015年1期)2015-12-24
