
西南桦幼林冠层光合特征及其对造林密度的响应
2020-04-26 13:15:44
中南林业科技大学学报 2020年4期
(中国林业科学研究院 热带林业研究所,广东 广州 510520)
关键字:西南桦;冠层;光合特征;造林密度
光合作用是植物进行物质和能量交换的主要途径,是森林生态系统能量流动和物质循环的基础,其决定着林分生产力,亦是反应竞争力大小的重要指标。树冠是林木与外界环境最主要、最直接和最活跃的界面,同时,也是光合作用的物质基础及主要场所[1-3]。因此,林木生长与树冠结构等密切相关,且不同冠层因其所处光环境等的异质性[4,5],致使其光合能力存在明显差异。目前国内外已有香椿Cedrela sinensis[6]、油橄榄Olea europaea[7]、毛白杨Populus tomentosa[8]、欧 美 杨Populus×canadensis Moench[9]、格 木Erythrophleum fordii[1]、刺 槐Robinia pseudoacacia[5]等多个树种开展过不同冠层光合能力研究,均发现树冠上层和外层的饱和光合速率、光饱和点、光补偿点等高于树冠下层和内层。然而关于造林密度对林木冠层光合生理特征的影响目前还尚不清楚。
造林密度是人工林营建前需要确定的重要因子,其不仅决定着前期造林成本,亦决定了林分的竞争强度,进而影响枝条发育及树冠结构,最终影响到林木的生长速度及木材质量[10]。对于造林密度与树冠结构的关系,已得到国内外学者的广泛关注[11-13]。造林密度、树冠结构分布、冠层光环境和光合能力四者紧密相关、相互影响,因此对于人工林而言,明确不同造林密度条件下林木各冠层光合能力的差异,对于确定合理造林密度、最佳修枝强度等经营措施均至关重要。
西南桦Betula alnoides是我国热带、亚热带地区一个珍贵用材树种,其树干通直,材质优良,生长较迅速,对环境适应性强,适宜培育大径材[14]。近十余年来,随着国家退耕还林等政策的实施,我国西南桦人工林发展迅速。至2015年底,其种植面积已超过15万hm2[15]。近年来,随着市场对于西南桦优质大径级无节材需求的增加,前期对西南桦造林密度与林木生长、枝条发育、冠层结构以及修枝与木材质量等方面进行了一系列探讨[10,16-19],本研究则以6年生西南桦幼林为对象,开展6种造林密度条件下西南桦人工林不同冠层光合特征研究,从光合生理的角度为西南桦造林密度和修枝强度的合理配置提供理论依据,最终服务于西南桦大径级无节材高效培育。
1 研究地概况
西南桦密度试验林位于广东省郁南林场镇龙庙工区(111°53′E,23°05′N),海拔440 m。该地属于南亚热带季风气候,年均气温21.6℃,年均降水量1 400~1 700 mm,降雨主要集中在4-9月份,年平均湿度80%。土壤为赤红壤。试验林于2012年5月采用无性系组培苗营建,设置 2 500株·hm-2(2 m×2 m)、1 667株·hm-2(2 m×3 m)、1 250株·hm-2(2 m×4 m)、1 111株·hm-2(3 m×3 m)、833株·hm-2(3 m×4 m)、625株·hm-2(4 m×4 m) 6个密度处理,3次重复,每个小区面积约 0.33 hm2,试验林总面积约6.67 hm2。造林后3 a 内,每年砍草1次,施复合肥1次。
2 研究方法
2.1 试验铺设
2017年3月,在西南桦密度试验林每个小区内各设置一个30 m×30 m 样地,进行每木检尺,调查胸径、树高、枝下高、冠幅,结果显示,林分平均胸径6.46~6.93 cm,平均树高5.00~5.69 m,平均枝下高1.40~1.94 m,平均冠幅2.43~3.33 m。基于上述调查结果,于每个样地选择优势木3株,其生长表现见表1。对于每株优势木,从树高6 m处以下,按每段1 m 分冠层进行光响应曲线测定。
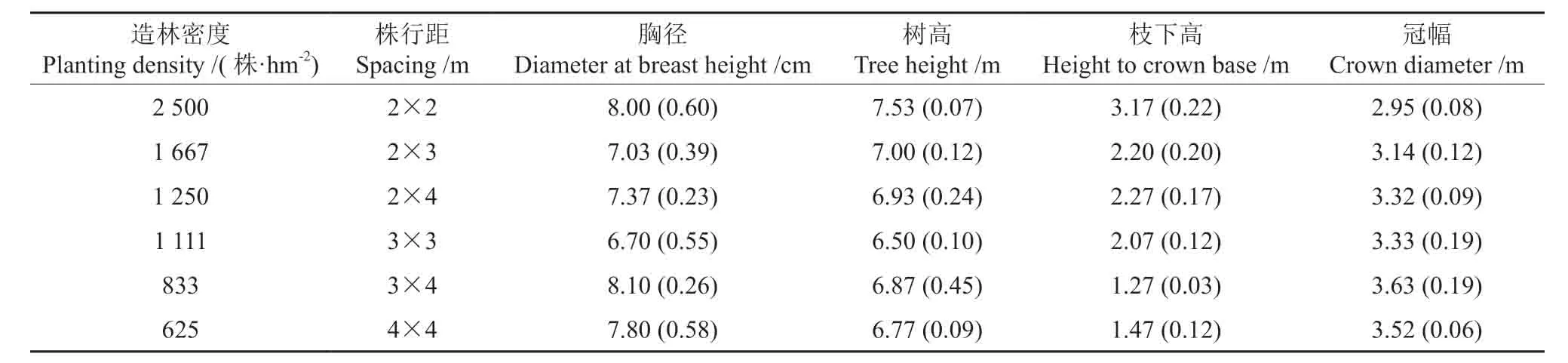
表1 幼龄西南桦优势木的生长表现Table1 Growth performance of sampled dominant trees in young plantation of Betula alnoides†
2018年5月,于上述每个样地3株优势木中随机选择一株进行光合测定。选择天气晴朗的上午,于所选西南桦优势木每个冠层(2~3 m、3~4 m、4~5 m、5~6 m)阳面中间位置选择一个平均枝,用高枝剪剪下立即插入水中,然后在水下剪除枝条基部,于枝条近顶部选择4 片发育完满叶,于9:30-11:30 期间进行光响应曲线测定[6,8]。测定时,将叶室温度设置为25℃,二氧化碳浓度控制采用二氧化碳注入系统,设定流量为400 μmol·m-2s-1,用Li-6400XT(美国LI-COR公司)红蓝光源控制光强,光强依次设为2 500、2 000、1 500、1 000、500、200、150、100、50、20和0 μmol·m-2s-1,进行光合测定。测定前若光 强较弱,则采用1 200 μmol·m-2s-1光强对测量叶进行20~30 min的光诱导。
2.2 数据分析
采用光合计算软件(光合计算4.1.1,叶子飘)[20]中的非直角双曲线修正模型计算表观量子效率、最大光合速率、光饱和点、光补偿点、暗呼吸速率,应 用SPSS 21.0(IBM-SPSS Inc.,Chicago,USA)统计软件中线性混合效应模型(GLMM)对不同冠层间及相同冠层不同密度条件下的光合特征差异进行分析,采用LSD 法进行多重比较。
3 研究结果
3.1 最大光合速率
造林密度和冠层高度位置对于西南桦幼林各冠层最大光合速率均有显著影响(P<0.05)。对于每个造林密度处理,随着冠层高度的上升,其最大光合速率均显著增加,特别是造林密度625株·hm-2和833株·hm-2两个处理增幅最大,其最大光合速率分别从最下层(2~3 m)的5.18 μmol·m-2s-1和4.33 μmol·m-2s-1显著增至上层树冠(5~6 m)的12.17 μmol·m-2s-1和10.95 μmol·m-2s-1,增幅分别达134.9%和152.9%。对于同一冠层而言,西南桦幼林最大光合速率随造林密度的增加总体呈降低趋势,如,冠层3~4 m和5~6 m,低造林密度处理(625和833株·hm-2)的最大光合速率显著大于高造林密度处理,而对于冠层2~3 m和4~5 m,最大光合速率随造林密度的增加虽存在降低趋势,但造林密度处理间差异未达显著水平(P≥0.05)。相对于冠层而言,造林密度对西南桦幼林最大光合速率的影响要小(表2)。
3.2 光饱和点
与最大光合速率一致,冠层间及造林密度处理间光饱和点亦存在显著差异。光饱和点随着冠层升高显著增大,且在低造林密度(625株·hm-2和833株·hm-2)条件下,下部(2~3 m)与上部(5~6 m)冠层光饱和点差异更大,下层光饱和点仅为上层的50%~60%,除造林密度处理625株·hm-2的3~4 m与4~5 m 以及造林密度处理1 250株·hm-2的4~5 m与5~6 m 冠层外,各密度处理的所有冠层间,西南桦幼林光饱和点均存在显著差异。对于同一冠层而言,西南桦光饱和点随着造林密度的增加呈现下降趋势,但大多为最低造林密度处理显著高于高密度处理,而且对于2~3 m 冠层,造林密度处理间光饱和点无显著差异(表2)。

表2 不同造林密度西南桦幼林冠层间最大光合速率、光饱和点差异分析†Table2 Variance of the maximum photosynthetic rate and light saturation point between crown layers of young Betula alnoides plantations with different planting densities
3.3 表观量子效率
西南桦幼林树冠表观量子效率变化幅度为0.041 9~0.083 2。随着冠层升高,表观量子效率并未呈现出显著增加或降低趋势。然而对于同一冠层而言,表观量子效率随造林密度的增加总体上呈现显著降低趋势。对于冠层3~4 m,625、833和1 111株·hm-2密度处理的表观量子效率显著高于其它密度处理;对于冠层4~5 m,这3个密度处理表观量子效率亦显著高于1 667和2 500株·hm-2处理;5~6 m和2~3 m 冠层的表观量子效率在不同密度处理间差异均不显著(表3)。

表3 不同造林密度西南桦幼林冠层间表观量子效率、光补偿点、暗呼吸速率差异分析†Table3 Variance analysis of apparent quantum yield,light compensation point and dark respiration rate of crown layers under different planting densities for young B.alnoides
3.4 光补偿点
西南桦幼林冠层光补偿点随冠层升高呈显著增大趋势。对于各密度处理5~6 m 冠层的光补偿点均显著高于2~3 m和3~4 m 或4~5 m 冠层,其余冠层间差异不显著。造林密度对西南桦幼林冠层光补偿点无显著影响;从下部至上部4个冠层,光补偿点的变化幅度分别为13~15 μmol·m-2s-1、10~15 μmol·m-2s-1、14~18 μmol·m-2s-1、19~24 μmol·m-2s-1(表3)。
3.5 暗呼吸速率
暗呼吸速率指林木在光照强度为0时,单位时间、单位面积林木通过呼吸消耗氧气和有机物生成二氧化碳的量。本研究中,冠层高度和造林密度对于西南桦幼林树冠的暗呼吸速率均无显著影响,其变化范围为0.61~0.96 μmol·m-2s-1(表3)。
4 讨论与结论
植物光合作用过程中,最大光合速率作为指示光合作用与光强关系的上限临界值,是衡量植物光合能力的重要指标[21];而光饱和点是反应植物对光能的利用能力,其值越高说明植物对强光的适应性越强[22];两者均在不同程度上决定着植物的光合能力。本研究中,随冠层升高,西南桦幼林冠层最大光合速率和光饱和点显著增大,换言之,光合能力及对强光的利用能力随冠层升高而增大,此与前人对于栲树Castanopsis fargesii[23]、毛白杨Populus tomentosa[8]、格木Erythrophleum fordii[1]、辽东栎Quercus liaotungensis[2]等树种的研究结果一致。冠层光合能力的差异主要与其所处光环境有关,因为树冠上部枝叶光照条件优越,而下部由于树冠内枝叶分布格局的影响,只能获得部分透射光或者散射光,且强度较弱。由此可见,不同冠层叶片所处光照条件的差异[4,24],不仅影响到各冠层的蒸腾速率和水分利用效率,亦致使其在长期适应过程中逐渐改变内在结构和生理代谢,以提升整个树冠的光合能力[25]。本研究中,随造林密度的增加,冠层(3~4 m)和冠层(5~6 m)的最大光合速率以及光饱和点显著降低,此与张旺锋等对棉花Gossypium hirsutum[26]和吕丽华等对夏玉米Zea mays[27]等农作物的光合特性研究结果类似。因为不同密度条件下植物冠层结构、叶片形态和数量以及群体内和冠内光环境存在差异[24,28-29]。对于林木而言,通常随造林密度增加,冠幅减小,枝下高增大,造成枝叶分布更加集中,冠层结构不合理,林木间及冠层间竞争及遮挡更加严重[19,28],林分内散射辐射与直射辐射透过率及利用率低,进而影响到林木各冠层的光合能力。而对于冠层4~5 m,受造林密度和冠层的交互作用以及高造林密度西南桦树冠枝条形态分布影响[10],随造林密度增加光合能力虽有降低趋势,但是不同处理间差异并未达到显著水平。此外,不同造林密度的株行距配置差异、6 m 以上冠层长度以及密度对冠层养分分配的影响等亦可能是导致造林密度影响规律较为复杂的原因,因为叶片的光合能力与叶片氮含量密切相关[6,30-31]。
表观量子效率和光补偿点均被作为重要指标用以指示植物对弱光的利用能力[32]。本研究中,低造林密度处理西南桦下部树冠表观量子效率显著高于高造林密度处理,下部冠层的光补偿点亦显著低于上部冠层(5~6 m),表明低造林密度树冠以及下部冠层对于弱光的利用能力要分别高于高造林密度和上部冠层,此与胡耀升等[2]对辽东栎、李勇等[32]对油松Pinus tabuliformis、刘强等[33]对长白落叶松Larix olgensis等的研究结果相似。造成此现象的主要原因在于林木下部树冠对于所处弱光环境的一种适应,而对于低造林密度林分,由于具有较长的树冠及较大冠幅,上部多层枝叶的遮挡致使其透光性较差,因此其下部冠层光环境更弱,造成其表观量子效率显著高于高造林密度林分,此亦可从低造林密度林分树冠下部的表观量子效率更大得到验证。而对于上部冠层,由于其所处光环境受冠层及造林密度影响较小,所以其表观量子效率无显著差异。归根结底,由造林密度和树冠结构造成的冠层光环境的异质性是影响冠层光合生理特性差异的最重要因子。
从光合能力的角度来说,上部冠层总体表现出强阳性特征,其相对于下部冠层具有更大的光合能力,而且西南桦冠层自身垂直方向相互遮挡的影响要高于因密度造成的相邻木在水平方向上树冠间的相互影响。然而,适宜的造林密度可进一步提升林木整个树冠及林分的光合能力,因为密度过高,光能在纵向利用不够充分,且林木间竞争及遮挡严重,而造林密度过低,林分内光耗散亦高,不能充分利用林地内的光合有效辐射。因此研究西南桦不同冠层叶片光合特征及其对造林密度的响应,将有助于充分利用林地空间及光合有效辐射,避免过多的光耗散,从而构建高产林分。另外,从本研究中所得不同冠层光合能力的差异亦可初步判别出对生长影响较小的冠层,如低造林密度处理(625和833株·hm-2)的2~3 m 冠层,进而通过修枝优化树冠结构,改善林内环境,提高树冠的总净光合同化能力。本研究仅对同一时间、同一方位不同冠层中部的平均枝进行了研究,具有其局限性,因此未来对于不同方位、水平冠层深度枝条的光合及生理特征(水分利用效率、蒸腾速率等)进行测定,并结合叶片形态(大小、长宽比等)、数量及分布特征(叶倾角、方位角、比叶面积等)等指标开展研究,将进一步将修枝工作明确到枝,此亦是珍贵树种高效培育的一个重要方面。
猜你喜欢
中国农业信息(2022年1期)2022-05-25 13:31:46
农业机械学报(2021年11期)2021-12-07 05:36:44
大气科学(2021年1期)2021-04-16 07:34:18
金秋(2021年20期)2021-02-16 00:58:14
江苏农业科学(2020年12期)2020-07-20 03:26:38
西安科技大学学报(社会科学版)(2019年1期)2019-09-10 07:22:44
现代园艺(2018年2期)2018-03-15 08:00:09
农业环境科学学报(2017年2期)2017-03-20 14:57:37
福建农业科技(2015年1期)2015-02-27 10:20:38
新课程·中旬(2014年10期)2014-12-26 04:47:30
