
二倍体棉种组织培养研究进展
2020-04-17 11:40孙悦华金平
棉花学报 2020年2期
孙悦,华金平
(中国农业大学农学院棉花遗传育种与基因组研究实验室/作物遗传改良北京市重点实验室/杂种优势利用教育部重点实验室,北京100193)
棉花是重要的经济作物。棉属共有53 个种[1],除了栽培种陆地棉(Gossypium hirsutum)和海岛棉(G.barbadense)等7 个四倍体棉种外,其余的皆为二倍体棉种。46 个二倍体棉种染色体组分别属于 A、B、C、D、E、F、G 和 K 组。二倍体棉种为四倍体栽培种提供了亲本(祖先)种,在棉属进化与多倍化机制研究方面有着重要意义。 研究表明,异源四倍体棉种是由两个不同的二倍体种杂交、染色体加倍而来的[2-7]。
二倍体棉种 A、B、E、F 组起源于亚非大陆,D组起源于美洲大陆,C、G、K 组起源于澳洲等地。A 组草棉、亚洲棉是栽培种,其他棉种均为野生种。 不同棉种株型差异大,有草本、灌木到乔木等类型。 二倍体棉种遗传多样性丰富,是棉花遗传改良的重要种质资源。 一些棉种具有高产、抗病虫害、耐逆、优异纤维品质性状的基因资源[1,8];已发现亚洲棉、异常棉、哈克尼西棉、三裂棉胞质不育系,其中哈克尼西棉、三裂棉细胞质获得了完全雄性不育系,并实现了三系配套[9]。
棉花远缘杂交在二倍体棉种资源利用中取得了成功,美国学者创造ATH(亚洲棉、瑟伯氏棉、陆地棉)三元杂种,成功改良了陆地棉纤维品质。 近期分子生物学研究涉及更多,如构建了亚洲棉 cDNA 文库[10],其中 WRKY 转录因子的表达与耐旱有关;雷蒙德氏棉WRKY 基因,参与棉花低温胁迫应答反应[11]。
组织培养技术是现代生物技术的基础。 棉花组织培养研究难度大,但是在种质资源改良与保存、杂种优势的利用、体细胞融合、基因工程等过程中有着巨大潜力, 对于建立转基因技术体系、开展体细胞融合、 功能基因组研究具有重要意义。 相关研究主要涉及植株器官与细胞培养的再生途径、原生质体培养和体细胞培养与融合等。
2 二倍体棉种组织培养体系
20 世纪70 年代以来, 逐渐开始初步建立棉花体细胞培养、茎尖培养、原生质体培养体系的探索。 棉花植株的再生途径可分为两类:一类是体细胞胚胎发生途径,外植体经过体细胞发生形成胚状体,胚状体发育获得再生苗;一类是愈伤组织途径,外植体先诱导获得愈伤组织,继而诱导愈伤组织分化产生芽和根并获得再生植株。二倍体棉种的再生体系建立相对于四倍体棉种难度更大, 现有再生途径主要为体细胞胚胎发生[12-26]。
2.1 体细胞培养
棉花体细胞培养体系,是棉花植株再生应用最为广泛的技术。1979 年,Price 等首次成功从克劳茨基棉的悬浮培养中获得了胚状体[12];随后,获得了高频胚胎发生的陆地棉柯字棉再生植株,开创了棉花体细胞培养及植株再生研究[15-16,18]。
二倍体棉种体胚发生方式是以下胚轴、叶片等作为外植体,诱导并增殖愈伤组织,随后诱导分化出胚性愈伤组织,进而获得再生植株。
迄今,获得了亚洲棉、异常棉、斯特提棉、瑟伯氏棉、戴维逊氏棉、克劳茨基棉、旱地棉、雷蒙德氏棉、野生拟似棉、司笃克氏棉、灰白棉、比克氏棉、 澳洲棉、 奈尔逊氏棉等14 个种体细胞胚[12-24];获得再生植株的棉种有:克劳茨基棉、野生拟似棉、亚洲棉、戴维逊氏棉、雷蒙德氏棉、司笃克氏棉、奈尔逊氏棉[13-14,17-20,22](表1)。
2.1.1基因型。基因型是二倍体棉种体细胞胚胎发生和植株能否再生的决定性因素[25]。 不同基因型的棉种体细胞胚形成能力不同。 二倍体亚洲棉的体细胞胚发生的能力明显高于草棉[15];不同的野生棉在相同的培养条件下体胚发生能力相差较大[20]。总之,大多二倍体棉种体细胞胚胎发生和植株再生的能力较低、难以观察到体细胞胚胎发生,少数棉种能够形成体细胞胚胎但难以得到再生植株。
2.1.2外植体的选择。外植体的选择决定棉花再生的成败。 一般选择幼嫩的组织或器官作为外植体,常用的有下胚轴、子叶、幼茎等,其中无菌苗的下胚轴最佳。 二倍体棉种体细胞胚胎发生过程中,下胚轴产生的愈伤组织最多[21]且产生的速度快[13]、体胚的转化率较子叶等其他外植体高,且继代培养的时间相对较短,从而可减少体细胞无性系变异[26]。
2.1.3培养基与培养条件。不同棉种的体细胞胚胎发生与植株再生对植物激素等外界因素的反应各有不同,通过不同激素组合、盐离子添加、不同碳氮源组合等调控方式,可找到与棉种相适应的培养条件的组合,从而提高二倍体棉种体细胞胚的产量与萌发能力(表2)。 成功获得再生植株的二倍体棉种,对培养基尤其是激素配比用量要求较严,只有在严格、适宜的培养基和培养条件下才能成功。
棉花组织培养常用激素有2,4-D (2,4- 二氯苯 氧 乙 酸 ,2,4-Dichlorophenoxyacetic acid)、IBA(吲哚丁酸,Indole butyric acid)、NAA (萘乙酸,1-Naphthylacetic acid)、KT(激动素,Kinetin)等。一般来说,2,4-D 能有效促进愈伤组织生长,并且
提高体胚转化率,在野生棉胚性愈伤组织诱导过程中可减少愈伤组织的白化死亡。 其中,NAA 为诱导亚洲棉胚性愈伤组织的最佳激素。

表1 二倍体棉种组织培养案例*Table 1 A list of diploid cotton tissue culture*

献文考参ces eferen R 3][1 4][1 7][1表2二倍体棉种体细胞培养再生植株培养体系Table2Alist of diploidcottonregenerationviasomaticembryogenesis件条养培基养培体植外组因基种棉体倍二s ndition re co ltu u C ajor steps lved in m vo in edium M t lan xp E e om en G loid cotton ip D荧白冷,/10 D L d 14 perio oto h P期周光;℃1)±8(2 1-L ol·m μ 0.9+B S:M ctionindu lture genic cu bryo m E导诱胚体叶子、轴胚下D 3-k棉基茨劳克ight L强光,t ligh t rescen fluo ite h w ol o C灯光;.8;pH51 KT-·L ol m.32 μ 2+2,4-Dcotyls ypo H m ianu lotzsch.k G 1.-2·s-m l·o m μ 33 sity ten in ryo b d em n an liferatio re progenic cultu bryo m E育发胎胚与殖增胚体onandcotyled 1 KT+-·L l o m μ 0.93+1 2,4-D-·L l o m 5 μ 0.04+B:MS t en mdevelop 1 IBA;-L l·o m μ 2.46-D1 2,4-·L l o m 3 μ 0.2+in liquid B:MS re n cultu sio en usp S养培浮悬;1 KT-·L ol m μ 0.93+activated %0.5+B S:1/4 M re lantlets cultuegenerated p R养培苗生再n.carbo ight L强光:duction in s allu C导诱织组伤愈1-·L g m 1+1 IAA-·L g 2 m+:MSctionallus indu C导诱织组伤愈轴胚下D 6棉似拟生野,/9D 5 L 1 eriod otop h P期周光,lx 00 0 0 1 sity ten in;T K cotyls ypo H es ssypioid.go G;℃1)9±(2+:MS n inatio d germ an ction du ryo in b atic em m o S发萌与导诱胚体d n an uctio ryo ind b atic em om S发萌与导诱胚体.1 LH-·L g.5 01 2IP+-·L g m 3+A1 NA-·L g 0.1 m;0 lx 00 5 0~t intensity 4 00 igh L强光:n inatio germ.;28 ℃2 D /1 d 12 L perio oto h P期周光期周光,℃2): (28±re allus cultu C养培织组伤愈;pH1 KT-·L g 1 m+A1 NA-·L g 2 m+:MSallus initiation C织组伤愈轴胚下A 2棉洲亚0 7 sity inten ight L强光,/8 D L 16 eriod otop h P 5.8;cotyls ypo H reum .arbo G速转re:n cultuensiousp S养培浮-1。 悬2·s-m l·o m μ d an callus selection ryogenic b m E持保与择选织组伤愈性胚d 16 perio to ho P期周光-1,·min r 30 en speed 1 ak h S;.8;pH51 KT-·L g m 0.1+A1 NA-·L g 0.5 m+:MS ce an ainten m.8 ℃,2/8 D L in S lture: M cu ryo initiation b id em iqu L织组伤愈性胚养培浮悬rary po d tem 3程过水脱3 d行进前基养培胚体);O 3 NH 4 o N,nO 3 N K led(doub id liqu re efo b tissue callus f o esiccation d r o starvation +S:M ration atu m and ation in ryo germ b atic em m o S熟成和发萌胚体.2 0+A1 NA-·L ol m 2.6 m+:MS ia ed transfer to m 5.8.;pH1 sucrose-·L ol 4 m.0 0+A1 NA-·L ol m 0.3 m.1 KT-L ol·m 0.2 m1 glucose +-L ol·m
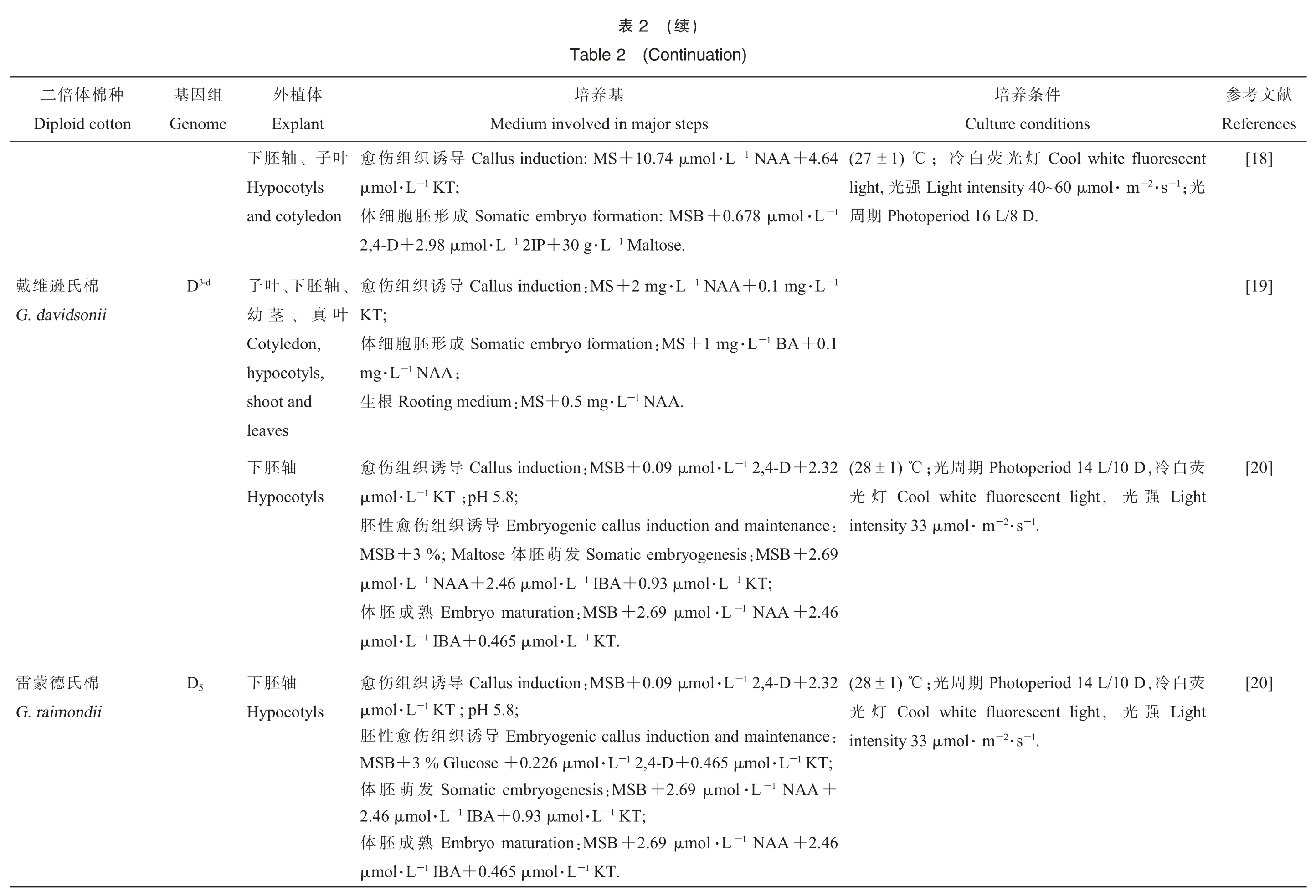
献文考参ces eferen R 8][1 9][1 0][2 0][2件条养培s ndition re co ltu u C rescent ite fluo h ol w o C灯光荧白冷;1) ℃±7(2光2-·s1;-m·ol m 0 μ 6 0~ight intensity 4 L强光t,ligh.D/8 L d 16 perio hoto P期周荧白冷,/10 D L d 14 perio oto h P期周光;℃1)±8(2 ight L强光,t ligh t rescen fluo ite h w ol o C灯光1.2-·s-m·l o m μ 33 sity ten in荧白冷,/10 D L d 14 perio oto h P期周光;℃1)±8(2 ight L强光,t ligh t rescen fluo ite h w ol o C灯光1.2-·s-m·l o m μ 33 sity ten in)续(2表)n tio a u tin n o(C 2 le b a T基养培体植外组因基种棉体倍二ajor steps lved in m vo in edium M t lan xp E e om en G loid cotton ip D 4.64A+1 NA-·L ol m μ 0.74 1+S M n:uctio allus ind C导诱织组伤愈叶子、轴胚下;1 KT-m o l·L μ cotyls ypoH 1-·L l o m 8 μ 0.67+B S: M ation ryo form b atic em om S成形胚胞细体on and cotyled altose.-1 M·L 30 g+1 2IP-·L ol m.98 μ 2+2,4-D 1-·L g 0.1 m+A1 NA-·L g 2 m+:MS n uctio allus ind C导诱织组伤愈、轴胚下、叶子3-d D棉氏逊维戴;T K叶真、茎幼nii avidso.d G 0.11 BA+-·L g m 1+:MSation form bryo atic em om S成形胚胞细体,on otyled C;A1 NA-·L g m cotyls,ypo h.A1 NA-·L g m 0.5+:MSm ediu g m ootin R根生d ot an sholeaves 2 2.3+-D1 2,4-·L l o m μ 0.09+B:MSctions indu allu C导诱织组伤愈轴胚下.8;;pH51 KT-m o l·L μ cotyls ypo H:ce aintenan m and ction du ryogenic callus in b m E导诱织组伤愈性胚9 2.6+B:MS esis gen bryo atic em m o S发萌胚体se alto M;%3+B S M;1 KT-·L ol m μ 0.93+1 IBA-·L l o m 6 μ 2.4+A1 NA-m o l·L μ 6 2.4+A1 NA-·L ol m.69 μ 2+B:MSration atu ryo m b m E熟成胚体.1 KT-m o l·L 65 μ0.4+1 IBA-m o l·L μ 2 2.3+-D1 2,4-·L l o m μ 0.09+B:MSduction allus in C导诱织组伤愈轴胚下D 5棉氏德蒙雷5.8;H p;1 KT-m o l·L μ cotyls ypo H dii on.raim G:ce aintenan m and ction du ryogenic callus in b m E导诱织组伤愈性胚;1 KT-·L ol m μ 65.4 01 2,4-D+-·L ol m μ.226 0 se +co lu G%3+B S M+A1 NA-·L ol m μ 2.69+B:MS genesis bryo atic em m o S发萌胚体;1 KT-L ol·m μ 0.93+1 IBA-L l·o m μ 2.46 6 2.4+A1 NA-·L ol m.69 μ 2+B:MSration atu ryo m b m E熟成胚体.1 KT-m o l·L 65 μ0.4+1 IBA-m o l·L μ

?
胚性愈伤组织诱导培养基的氮源去除NH4NO3(硝酸铵),同时 KNO3(硝酸钾)加倍可以成功诱导体胚发生[17,26];碳源一般采用葡萄糖。 但是,诱导克劳茨基棉体胚需要采用蔗糖和麦芽糖混合碳源,此时胚胎发生率要比单独使用葡萄糖高[20]。 麦芽糖是胚性愈伤组织增殖和体胚形成过程的最佳碳源之一[22]。
愈伤组织诱导继代培养过程中,选择悬浮培养的培养方式可有效增加体细胞数目,缩短体细胞胚胎发生时间[13],较固体培养基更为有效。 此外, 培养基加入一定的活性炭更利于体细胞萌发、生根。
2.2 茎尖培养
棉花茎尖培养使用的是茎尖完整的分生组织,可以规避二倍体棉种在体胚转化过程中基因型限制问题, 且再生植株所需时间也相对较短,有效减少因继代培养时间过长而引起的无性系变异。 目前茎尖培养研究主要集中于亚洲棉和比克氏棉[27-30],其中比克氏棉茎尖培养获得再生植株的时间[27]与体细胞培养[20]相比,可节省4 个月左右。 但通过茎尖培养建立再生系统的植株较少(表3)。
2.3 花药培养
花药培养培育单倍体植株,可用于探究胚胎发生的机理。 棉花花药培养难度较大,仍处于探索阶段。 棉花花药培养受基因型、花药培养的时期、培养条件等影响,进展不大,其中亚洲棉较易诱导产生愈伤组织,但难以从花药培养中获得再生植株;戴维逊氏棉较难诱导产生愈伤[31]。
2.4 原生质体培养
原生质体培养植株再生,需要经历原生质体分离、纯化培养、细胞壁再生、体胚发生等过程。原生质体培养和体细胞培养相比, 难度更大、培养过程更为复杂。 目前,仅有克劳茨基棉和戴维逊氏棉两个棉种培养成功的报道,其主要来源为胚性细胞悬浮系(表4)[19,32-36]。
二倍体棉种原生质体培养的周期较长,培养方式有平板培养法、液体浅层培养法、固液相结合培养法等,对培养基、黑暗等培养条件要求较为严格(表5)。 其中,使用改良KM8P 进行棉花二倍体种原生质体培养,成功得到了再生植株。
2.5 种间杂种
利用原生质体融合技术进行体细胞杂交,从而获得种间杂种。孙玉强[37]于2005 年首次成功通过来源于胚性愈伤组织和体细胞幼胚的原生质体融合得到棉花体细胞杂种植株,其中,陆地棉和克劳茨基棉、陆地棉和戴维逊氏棉、陆地棉和比克棉、陆地棉和司笃克氏棉组合的种间杂种植株的可育性较高,且种间杂种植株在表观上可观察到杂种优势。
3 二倍体棉种组织培养研究展望
棉花组织培养难度较大,存在较大的种间差异,其后续研究方向有以下几点:
3.1 胚性愈伤组织诱导
棉花组织培养的第一步通常为愈伤组织的诱导。 不同二倍体棉种产生愈伤组织的能力不同,其中以亚洲棉较为容易诱导愈伤组织。 同一棉种也可通过不同激素组合诱导愈伤组织[38],但培养得到的愈伤组织形态各有不同:早期愈伤组织颜色较浅、部分质地透明,排列疏松;晚期愈伤组织颜色变深,质地松软。 胚性愈伤组织为黄绿色,质地较脆[39]。 可针对不同的棉种,通过不同的激素等组合,诱导愈伤组织。
3.2 消除基因型限制
体细胞培养体系研究见于亚洲棉、克劳茨基棉、戴维逊氏棉等棉种。 基因型差异导致培养周期的延长,易造成体胚发育异常、体胚转化率降低等,也使得愈伤组织继代培养实验重复性差[23]。此外,亚洲棉[40]和雷蒙德氏棉[41]已完成全基因组测序,为研究关键功能基因提供了便利。
体细胞培养中愈伤组织分化为胚性愈伤组织是主要的瓶颈。 激活二倍体棉种控制胚胎发生能力的相关基因, 可能解决体细胞胚发生问题。体细胞胚胎发生受体激酶SERKs基因参与植物发育、激素感应及病理反应等。GhSERK1 基因在胚性愈伤组织时表达量达到高峰,过表达该基因能够加快愈伤组织形成胚性愈伤组织进程[42],缩短培养时间,而非本质上提高愈伤组织分化率。

株植生再养培尖茎种棉体倍二3表re ltu cu tip m te s ia v n tio ra e n e g re n tto o c id lo ip f d list o A 3 le b a T献文考参件条养培基养培体植外组因基种棉体倍二ces eferen R s dition re conultu C r steps ajo lved in m invoium ed M t plan x E e enom G n cotto id iplo D][28(L,light;/8 D 6 L toperiod 1ho P期周光;℃2)0±(3+A1 NA-·L ol m-10.7 μ.3 0+:MSctionoots indu h S养培芽发始起节叶子的叶子无A 2棉洲亚1;pH-2-·s·m ol m μ tensity 50 t in igh L强光);ark,d D;P1 BA-·L l o m r4.4 μ 2.2oary on otyled C.arboreum G 5.8..6~5 P1 BA-·L ol m μ.4 4+:MSoots proliferated h S养培殖增代继f o evoidodes dn1 glucose.-·L 20 g+:MSduction ts in oo R养培根生s on cotyled][29荧白冷;/8 D 6 L toperiod 1ho P期周光;℃1)5±(2养培芽发始起节叶子的叶子无t igh L强光,orescent light hite flu l w oo C灯光;P1 BA-·L g.5 or 2.0 m 1+:MS n uctio ts ind oo le sh ultip M ary on otyled C H5.8.p-1;2·s-m ol·m μ 50~tensity 40 in 1-L g·m.5 0+S:M ngated d elo oots harvested an h S养培殖增代继f o evoidodes d n;A 3 G s on cotyled.S M duction:1/2 ts in oo R养培根生][30期周光;0 lx tensity 2 00 t in igh L强光;℃3)2±(2·g m.0 1+:MS n.liferatio ro and p itiation ot in ho S养培殖增芽发始起尖茎叶子无./8 D 6 L 1 eriod otop h P;1 KT-L g·m 0.5+P1 BA-L ot tips ho S.A-1 IB·L g m 0.75uid +liq in:MSduction ts in oo R养培根生s on tyled out co ith w][27荧白冷,/8 D 6 L toperiod 1ho P期周光;℃1)8±(2 1-·L g m.1 0+P1 BA-·L g m 4+B:MSctionoots indu h S养培芽发始起节叶子的叶子无G 1棉氏克比t igh L强光,orescent light hite flu l w oo C灯光.85;;pH5Z D T ary on otyled C ickii .b G 1.2-·s-·m ol m μ tensity 33 in;1 GA3-·L g 5 m.0 0+B:1/2 MS lture bcu u S养培殖增代继evoid odes d n on.carb ctivated A%0.5+S M duction:1/2 ts in oo R养培根生s on of cotyled动激:T hthylacetic acid;K ap-N 1酸乙萘:A A;N in 5 vitam B andre system ltu cu oog k S&ige urash M物机5 有B+液母S:M B S;M re system ltu g cu oo k S&ige urash M液母S:M S:M注.iazuron hid T物生衍类脲基苯,)隆苯噻(灵叶脱:Z;TD 3 ibberellin A G素霉赤:3;GA rine opu in zylam en-B 6呤嘌基氨苄:6-P;BA utyric acid dole b In酸丁哚吲:;IBA etin in K素

养培体质生原种棉体倍二4表s ie c e p s n tto co id lo ip f d o re ltu u t c s la p to ro P 4 le b a T献文考参果结养培料材eferences R esult R aterial M 2][3 ass ell m C团胞细ther callus n A织组伤愈药花3][3 ass ell m C团胞细s ocotyl callu yp H织组伤愈轴胚下4][3 n plant eratio egen R株植生再atic cell ature som m d im n an sio en sp nic cell su bryo m E胞细胚体熟成未,液浮悬胞细胎胚5][3 ass ell m C团胞细allus C织组伤愈D 2-2essii arkn.h G棉西尼克哈9][1 ass ell m C团胞细allus C织组伤愈D 3-diiavidson.d G棉氏逊维戴6][3 n plant eratio egen R株植生再genic callus bryo m E织组伤愈性胚

件条养培株植生再养培体质生原种棉体倍二5表n tio ltiva u st c la p to ro p ia v n tio ra e n e g re n tto o c id lo ip r d l fo o c to ro p e h T 5 le b a T献文考参件条养培基养培料材组因基种棉体倍二eferences R s dition ulture con C s r step ajo m in involvedm iu ed M aterial M e om en G tton species loid co ip D 4][3荡振;ark D暗黑;:28 ℃n e solutionzym E解酶酶胶果%.5 1 ellulase +C酶素维纤:3%lution e so nzym E解酶浮悬胞细胎胚D 3-k棉基茨劳克);h 24~-1(14·min r g 40hakin S;9M W P C icellulase +em H酶素维纤半%0.5 ase +ectin P胚体熟成未,液m ianuklotzsch.G暗黑;:28℃lture layer cu id hin liqu T养培层浅1-·L l o m μ 0.452+8P:KMid layer cultureliqu hin T养培层浅体液ryonic b m E胞细ark;D.1 KT-·L ol m μ.465 0+2,4-Dsion spen cell su强光,bes t tu rescen aylight fluo D管光荧光日)+O 3 NH 4 o N(n B:MSction du s in bryo atic em m o S养培导诱胚体ature m and im 1.-2 s-·m l o m μ t intensity33 igh L 1-·L ol m 85 μ2.6+1 IBA-·L ol m 60 μ2.4+O 31 KN-·L ol m m 91.7 18atic cell som;A 3 G+1 KT-·L l o m 7 μ 0.69+A AN炭性活%0.05+B:MS lture lantlets cuegenerated p R养培苗生再c tive charcoal.A 6][3;ark D暗黑;℃0)3:(28~n e solutionzym E解酶酶胶果%.5 1 ellulase +C酶素维纤:2%lution e so nzym E解酶织组伤愈性胚D 3-d棉氏逊维戴);24 h1(14~-·min 0 r g 4 in ak h S荡振;9M W P C icellulase +em H酶素维纤半%2 ase +ectin P genic bryo m E nii so david.G;℃)1±lasts culture:(28 top ro P养培体质生原1 2,4-D+-L l·o m 5 μ 0.4+8P M re:K ltu plasts cu to ro P养培体质生原callus.Dark暗黑;1 KT-L ol·m μ 0.93+A1 NA-L l·o m μ2.68:5ediumover solid m id ith liqu re w ltu ble-layer cu ou d养培层双液固.8P M K+B3 MS cm 1-L ol·7 μm.3 15 I,-1 KL ol·m μ 10O,H 2 5·O 41 CuS-L l·o m μ 0.1,OH 2 7·O 4gS-1 ML ol·m 1.0 m,O 3 N-1 KL ol·m m.0 1,O 4 PH 2-1 KL ol·m m.2 0,O2H 2·l 21 CaC-L ol·m m:109M W P:C注;inetin K素动激:T;K ylacetic acid th aph 1-N酸乙萘:A A;N oxyacetic acid en rophichlo-D 2,4酸乙氧苯氯二:-D;2,4edium m 8P M K基养培8P M:K P 8 M 5.8;K H p l,ito ann m/v)(m%,9 S E M.3 berellin A ib G素霉赤:3 A utyric acid;Gdole b In酸丁哚吲:A IB
Wuschel(WUS)基因表达能够促进体细胞胚胎的发育,该基因在下胚轴与非胚性愈伤组织阶段表达微弱[38,43]。 过表达该基因,极大地提升了难分化棉种的愈伤组织分化率, 体胚发生率提高4倍[42]。
生长素的合成、运输、响应,在植物体细胞胚胎发生、 发育等全过程中起着重要的调节作用。亚洲棉ARF 家族是参与生物素信号调节的重要转录因子,影响生长素的运输和信号转导,参与根、叶的形成[44]。
3.3 原生质体融合
原生质体融合,可以克服植物远缘杂交不亲和性,创造体细胞杂种。 通过电融合法获得多种由陆地棉和二倍体棉种克劳茨基棉、 比克氏棉、司笃克氏棉的原生质体对称融合产生的杂种植株[37,45-47]。将陆地棉与克劳茨基棉原生质体非对称融合,获得了多种有利的田间材料[48]。 目前,对于棉花原生质体融合, 仍需要扩大基因型范围,丰富材料基础。
猜你喜欢
中国生殖健康(2020年7期)2021-01-18
中国生殖健康(2020年5期)2021-01-18
新农业(2020年22期)2020-12-18
今日农业(2020年24期)2020-12-15
天津农业科学(2018年12期)2018-01-15
北京航空航天大学学报(2017年6期)2017-11-23
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
科学中国人(2015年23期)2015-07-12
医学研究杂志(2015年12期)2015-06-10
