
播期对棉铃对位叶蔗糖代谢及棉铃产量性状、纤维品质的影响
2020-04-17 11:40:16陈功彭金剑罗海华高欣袁长凯殷梦瑶向春玲王培培涂青华汤飞宇
棉花学报 2020年2期
陈功,彭金剑,罗海华,高欣,袁长凯,殷梦瑶,向春玲,王培培,涂青华,汤飞宇*
(1.江西农业大学农学院/ 作物生理生态与遗传育种教育部重点实验室,南昌330045;2.江西省农业综合行政执法总队,南昌330046)
江西省鄱阳湖植棉区通常于4 月上中旬播种。 由于此时该地区经常出现潮湿、寒冷的天气,不利于种子发芽和幼苗早期的生长发育,因此在棉花栽培中通常采取育苗移栽的方法为其出苗和早期幼苗生长提供较为适宜的微环境。 随着城镇化的发展和农村劳动力的转移,棉农迫切希望采用轻简化的种植技术,从传统精耕细作下的育苗移栽过渡到机械化直播,必然导致播种期的推迟;此外,南方多熟制棉田耕作制度的改革,如油(麦)后棉的推广也必然导致播种期的推迟[1]。 播种期的不同会引起棉铃生长中环境因子的变化,其中温度是对棉铃发育影响最为关键的环境因子[2]。 由于棉铃对位叶是棉铃生长发育所需碳同化物的主要来源[3],棉铃发育的质量直接影响到铃重、衣分等产量因素及纤维品质的形成。 蔗糖代谢调控棉铃及其对位叶碳水化合物的合成、分配与转运,从而影响棉铃产量性状及纤维品质的形成[4-6]。 因此研究播期对棉铃对位叶蔗糖代谢的影响,对于提高棉花产量和品质有重要意义。
蔗糖和淀粉均是棉铃对位叶光合作用的主要产物,其中蔗糖是碳水化合物长距离运输的主要形式,淀粉是碳水化合物贮藏的主要形式[7]。蔗糖代谢关键酶主要包括液泡酸性转化酶(Vacuolar acid invertas,VIN)、细胞壁酸性转化酶(Cell wall acid invertase,CWIN)、 蔗糖磷酸合成酶(Sucrose phosphate synthase,SPS)和蔗糖合成酶 (Sucrose synthase,SS)。 液泡酸性转化酶(VIN)和细胞壁酸性转化酶(CWIN)催化蔗糖不可逆分解为葡萄糖和果糖,分别位于细胞液泡内及细胞壁质外体空间, 催化作用适宜的pH 为3.5~5.5,VIN 为可溶性酶,CWIN 为不可溶性,以离子键结合于细胞壁[8];蔗糖磷酸合成酶(SPS)催化尿苷二磷酸葡萄糖 (Uridine diphosphate glucose,UDPG)和 6- 磷酸果糖合成蔗糖[8];蔗糖合成酶(SS)催化蔗糖可逆分解为尿苷二磷酸葡萄糖(UDPG)和果糖[8]。蔗糖代谢关键酶影响棉花叶片中碳水化合物的种类、含量及比例[8-9],也影响到碳水化合物的外运水平[10]。 蔗糖代谢关键酶的活性受低温的影响,但在不同器官中对低温的响应不同。 叶片SPS 和SS 在低温条件下活性下降[6]。 纤维SS、酸性转化酶和碱性转化酶在低温条件下活性上升,但SPS 活性下降[11-12]。目前对于棉铃对位叶中VIN 和CWIN 酶活性的研究鲜见报道。 同时,由于低温处理方式以及试验材料的不同,研究结果存在差异,对于棉铃对位叶蔗糖代谢对低温的响应机制尚不明确。 本文以不同播种期创造棉铃发育期间的温度差异,研究其对棉铃产量性状具有明显差异的2 个品系棉铃对位叶蔗糖代谢的影响,以及对棉铃产量性状及纤维品质的调节作用,为棉花生产上适宜播种期的安排提供参考依据。
1 材料与方法
1.1 材料与设计
试验材料为江西农业大学棉花课题组自育的陆地棉高品质系A201 和A705, 长期自交保纯,遗传性状稳定。 其中A201 为低衣分、高籽指类型,A705 为高衣分、低籽指类型。 试验于2016―2017 年在江西农业大学农业科技园进行,供试土壤为红壤旱地,呈微酸性(pH 6.0)。 前茬空闲,地力均匀,肥力中上,排灌方便,四周无高大建筑物和树木。 田间试验采用随机区组排列,设3 次重复,4 行区,行长7.7 m,行距1.0 m,株距0.3 m,小区面积30.8 m2,四周设保护行。 早播实为正常播种时间,采用营养钵育苗移栽;晚播采用大田直播。早播 2016 年 4 月 12 日播种,5 月 14 日移栽,2017 年 4 月 15 日播种,5 月 14 日移栽; 晚播分别 于 2016 年 5 月 6 日和 2017 年 5 月 28 日播种。 2 年施肥量相同, 全生育期施纯 N:276 kg·hm-2、P2O5:105 kg·hm-2、K2O:247.5 kg·hm-2。 磷肥和钾肥作基肥和花铃肥分2 次施用, 比例为1∶1;氮肥作基肥、花铃肥和盖顶肥分3 次施用,比例为1∶2∶1。 全生育期未进行化控,不整枝,于8 月中旬根据长势进行打顶。 苗期和蕾期各中耕除草1 次,花铃期根据天气情况灌溉1~2 次,根据田间需要进行病虫害和杂草控制。 2 年大田管理情况基本一致。
开花结铃期挂牌标记植株中部果枝(7~9 果枝)、内围果节当日白花,分别取5 DPA(开花后时间,Days post anthesis)、10 DPA、17 DPA、24 DPA、31 DPA、38 DPA、45 DPA 共 7 个时期的发育正常且无病虫为害的棉铃及其对位叶。 棉铃对位叶分成两部分:一部分作鲜样液氮速冻后及时保存于-80 ℃的超低温冰箱中,用于测定蔗糖代谢关键酶: 液泡转化酶 (VIN)、 细胞壁转化酶(CWIN)、蔗糖磷酸合成酶(SPS)和蔗糖合成酶(SS)的活性;一部分经 105 ℃杀青 30 min 后于60 ℃下烘干至恒重,烘干后的叶片用微型植物粉碎机磨碎并过32 目筛网,用于己糖(葡萄糖、果糖)、蔗糖和淀粉浓度的测定。
1.2 测定内容与方法
蔗糖、果糖、葡萄糖和淀粉的测定。 可溶性碳水化合物的提取:用万分之一电子天平称叶片干样50 mg,加入80%乙醇溶液3 mL(叶片材料先加入10 mg 活性炭粉末,用于去除色素)。 于混匀器混匀后,在80 ℃的水浴锅中提取30 min,取出后冷却至室温。 置 4 000 r·min-1离心机中离心10 min,转移上清液,剩余的残渣再次加入80%乙醇溶液3 mL,重复提取残渣2 次,合并3 次提取的上清液并用80%乙醇溶液定容至10 mL,用于葡萄糖、果糖、蔗糖浓度的测定。
淀粉的提取。 将上述用乙醇溶液提取后的残渣,放在60 ℃烘箱内烘至恒重。 加入2 mL 的蒸馏水,混匀器混匀,放入沸水中糊化15 min,冷却至室温。加入 9.2 mol·L-1高氯酸 2 mL,于震荡仪震荡 15 min, 然后加 4 mL 的蒸馏水,4 000 r·min-1下离心 10 min, 将上清液转移至 50 mL 的容量瓶中。 残渣再加入 4.6 mol·L-1的高氯酸2 mL,震荡 15 min,加蒸馏水 5 mL,混匀后在4 000 r·min-1下离心 10 min,合并上清液。 用5 mL 的蒸馏水洗涤残渣1 次,离心10 min,合并上清液,最后用蒸馏水定容至50 mL,用于淀粉浓度的测定。葡萄糖浓度的测定采用酶比色法[13],蔗糖、果糖浓度的测定采用间苯二酚法[14],淀粉浓度的测定采用蒽酮法[14]。
VIN、CWIN、SPS、SS 的提取和测定。 提取液的配制。 VIN、SPS 和 SS 提取液(pH 7.5)的组分包括 :50 mmol·L-1Hepes-NaOH,2 mmol·L-1Na2-EDTA,2.5 mmol·L-1DTT,10 mmol·L-1Mg-Cl2,0.05%Triton X-100 (V/V),1%(W/V) 难溶性PVP,10%甘油(V/V),0.3%β-巯基乙醇。 CWIN 的提取液另外再加入 0.8 mol·L-1NaCl(终浓度)即可。
粗酶液的提取。 称取0.4 g 鲜样放入预冷的研钵中,加入1 mL 提取液(预冷)和适量石英砂进行充分研磨,转移至10 mL PE 离心管,再加入1 mL 提取液清洗研钵1 次,合并混匀。 冰浴条件下提取30 min,每隔5 min 涡旋混匀1 次,以保证抽提完全。4 ℃ 4 500 r·min-1离心机中离心 20 min,将上清液置4 ℃冰箱中待测。剩余沉淀加入2 mL CWIN 的提取液,提取步骤同上。 SS 和 SPS 酶活性采用 UDPG 和果糖比色法[14];VIN 和 CWIN 酶活性测定参照Wang 等的DNS 比色法[15]。
产量性状及纤维品质的考察与检测。 棉铃成熟吐絮时,分别采摘每个小区棉株挂牌标记的剩余棉铃(无僵瓣花)30 个,晒干后称取铃重,经小型皮辊轧花机轧花后测定其纤维重、 种子重,从而计算衣分和籽指等产量性状。各小区取20 g 纤维送原农业部棉花纤维品质监督检验测试中心HVI900 系列测定, 检测指标包括纤维上半部平均长度(mm)、整齐度(%)、马克隆值、伸长率(%)、断裂比强度(cN·tex-1)共 5 个指标。
1.3 气象数据
气象数据来自国家气象中心南昌气象观测站(28°6' N,115°91' E),该站靠近试验地点(28°09'N,115°27'E)。 表1 列出了试验期间棉铃开花至吐絮期间日平均温度 (MDT), 日均最高温度(MDTmax),日均最低温度(MDTmin)、日均昼夜温差(MDTdif)和棉铃发育期间的总降水量。
1.4 数据分析
以小区为单位统计处理平均数, 采用SPSS20.0 统计分析软件包的一般线性模型(GLM)对实验数据进行方差分析,独立样本t测验分离处理平均数。 按基因型和年份对不同取样时期的碳水化合物浓度和蔗糖代谢酶活性取平均值,采用配对样本t测验比较早晚播期的差异。采用Origin8.5 作图。
2 结果与分析
2.1 早晚播棉铃发育的时长与温度差异
2 年晚播播种时间的差异是由于2016 年早晚播棉铃开花期相近,为创造铃期温度更大的差异,故进一步推迟2017 年晚播的播种日期。 2 年晚播铃期均长于早播,2016 年晚播铃期 (棉铃开花至吐絮)延长3 d,2017 年延长8 d;晚播铃期日平均温度、日均最高温度、日均最低温度、平均昼夜温差均低于早播(表1)。

表1 2016-2017 年棉铃开花到吐絮期日均温度、日均最高温度、日均最低温度、日均昼夜温差和总降水量Table 1 Mean daily temperature, mean daily maximum temperature, mean daily minimum temperature, mean diurnal temperature difference, and total precipitation during flowering to boll opening in 2016 and 2017

表2 播期对铃重、衣分和棉铃产量性状的影响Table 2 Effects of planting dates on boll size, lint percentage and within-boll yield components
2.2 播期对棉铃铃内产量性状的影响
由表2 可知, 2016 年和 2017 年 A201,及2016 年A705 晚播的铃重、单铃种子质量均显著高于早播;2 年试验A201 晚播的籽指均显著高于早播;2016 年A201 的单铃纤维质量表现为晚播显著大于早播;2 年试验A201 的衣分表现为晚播显著低于早播。 以上结果表明晚播相对低温条件下棉铃发育好于早播。
2.3 播期对棉铃纤维品质的影响
由表3 可知,2 年试验中晚播纤维上半部平均长度高于早播, 其中 2 年 A705 和 2016年A201 均达到显著水平;晚播纤维整齐度指数、伸长率和断裂比强度略高于早播,但差异均不显著; 马克隆值早播高于晚播,2017 年2品系均表现显著差异。 表明晚播纤维品质略好于早播。

表3 播期对纤维品质的影响Table 3 Effects of planting dates on fiber quality parameters
2.4 播期对棉铃对位叶碳水化合物浓度的影响
棉铃对位叶蔗糖水平整体趋势变化平缓,除2017 年开花后 5 d 外,2 年间 A705 和 A201 棉铃对位叶蔗糖浓度晚播均高于早播 (图1a)。 2016年晚播棉铃对位叶己糖浓度各时期均表现高于早播,2017 年早晚播的差异因取样时期而异,开花后5~10 d,早播己糖浓度大于晚播;开花后45 d,晚播高于早播(图 1b)。 A201 除 2016 年24 DPA 和 2017 年 10 DPA 外, 其他各取样时期淀粉浓度均表现为早播高于晚播,2017 年A705除10 DPA 外, 其他各时期淀粉浓度均表现早播高于晚播(图 1c)。

图1 不同播期条件下棉铃对位叶碳水化合物浓度的变化Fig.1 Dynamics of carbohydrate components concentration in boll-subtending leaf at two planting dates

表4 播期对棉铃对位叶己糖、蔗糖和淀粉的平均值和最大值的影响(2016―2017)Table 4 Effects of planting date on the mean, maximum contents of hexose, sucrose and starch in the leaf subtending to cotton boll(2016-2017)
由表 4 可知,2 年 A705 和 A201 棉铃对位叶各取样时期的蔗糖平均浓度及最大值晚播均显著高于早播。 2016 年A705 和A201 棉铃对位叶各取样时期的己糖平均浓度表现为晚播显著高于早播;2017 年A201 己糖浓度最大值表现为早播显著高于晚播。 不同播期棉铃对位叶的淀粉平均浓度和最大值的差异因基因型和年份而异。 综上所述,表明晚播相对低温条件下,棉铃对位叶蔗糖浓度高于早播(图1a,表4)。
2.5 播期对棉铃对位叶蔗糖代谢关键酶活性的影响
2 年棉铃对位叶VIN 酶活性表现为随开花后天数增加基本呈下降趋势(图2a),各取样时期的酶活性平均值除2016 年A201 外 (差异不显著),均表现为早播高于晚播,其中2016 年A705和2017 年A201 均达到显著水平。VIN 酶活性峰值均表现为早播高于晚播,2017 年 A705 和A201 达到显著水平 (表 5)。 2 年棉铃对位叶CWIN 酶活性除 2017 年 A201 的 24 DPA 和 45 DPA 外,其他各时期均表现为早播棉铃对位叶的CWIN 酶活性高于晚播(图2b),酶活性平均值2年均表现为早播显著高于晚播, 且酶活性峰值也表现为早播高于晚播,除2017 年A201 外均达到显著(表 5)。 2 年未发现 2 个品系的 SPS 酶活性对早晚播的响应具有一致性(图3a,表5)。 SS 酶活性 2016 年 2 品系除 A201 的 45 DPA 外,各时期早播均高于晚播。 2017 年A705 除 31DPA 和45 DPA 外,各时期均表现早播高于晚播,2017 年A201 在棉铃对位叶生长初期 (5 DPA) 和后期(31~45 DPA) 均表现早播高于晚播 (图3b)。2016 年 A705 和A201 的 SS 平均值和峰值均表现为早播显著大于晚播,2017 年 A705 的 SS 峰值表现为早播显著大于晚播,其余未见显著差异(表5)。 综上所述,表明晚播相对低温条件下,棉铃对位叶CWIN 和SS 酶活性受到抑制, 早晚播对VIN 和SPS 酶活性的影响因年份、 基因型和取样时期而异。
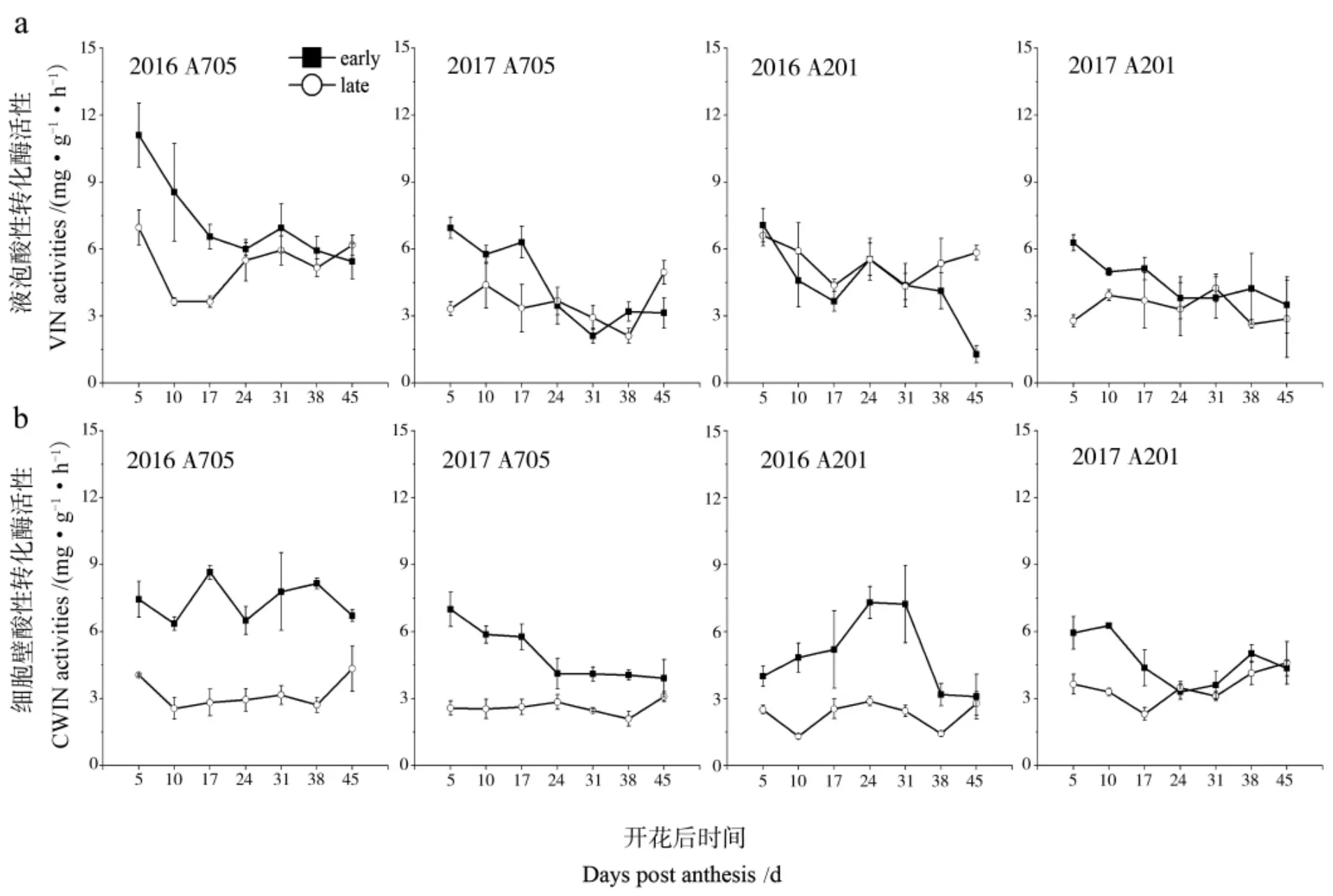
图2 不同播期条件下棉铃对位叶VIN 和CWIN 酶活性的变化Fig.2 Dynamics of VIN and CWIN activities in the leaf subtending to cotton boll at two planting dates

图3 不同播期条件下棉铃对位叶SPS 和SS 酶活性的变化Fig.3 Dynamics of SPS and SS activities in the boll-subtending leaf at two planting dates
3 讨论
3.1 不同播期条件下棉铃铃内产量性状及纤维品质的差异
棉铃发育受多种因素影响,温度被认为是对棉铃发育影响最大的环境因素。 Liakatas 等[16]报道低温(26 ℃/16.5 ℃ 昼温 /夜温)较高温(30 ℃/20 ℃昼温/夜温)提高了铃重和衣分。铃重、单铃纤维质量、 单铃种子质量和籽指随温度从18 ℃升高到25.5 ℃而增加,但在更高的温度(29.5 ℃)时快速下降[17]。 在本试验中,2 年 A705 和 A201铃重、单铃纤维质量、单铃种子质量和籽指均为晚播相对低温条件下表现更高,这可能是由于本试验晚播条件下铃期日均温 (2016 年25.2 ℃,2017 年 23.1 ℃)较早播(2016 年 26.8 ℃,2017 年28.3 ℃)更适宜棉铃生长发育。 且晚播条件下,棉铃具有更长的发育期,这可能是晚播棉铃铃重更高的原因之一。 Yeates 等[18]认为花铃期低温和低太阳幅射共同造成棉株生长速率下降,但棉铃充实阶段延长,从而达到高产。 棉纤维品质形成受温度影响,纤维正常发育的温度是20~30 ℃,日均温22 ℃被认为是纤维伸长的最适温度[17]。 这与本试验晚播所处的温度23.1~25.2 ℃更接近,这可能是A705 和A201 晚播纤维上半部平均长度更高的主要原因。 纤维长度是由伸长速率及快速伸长时间综合协调决定,研究发现高温增加伸长速率,缩短快速伸长时间,高温增加的伸长速率并不能补偿伸长时间缩短对纤维伸长造成的不利影响[19],而晚播低温条件下伸长时间的延长可补偿降低的伸长率[20]。 同样,晚播条件下纤维比强度增加,这与王友华等[21]和Bauer 等[22]人研究一致;但与Liakatas 等[16]的研究不同,他们认为在 30 ℃/20 ℃(昼 / 夜) 时的纤维比强度高于26 ℃/16.5 ℃(昼 / 夜)。 Lokhande 等[17]报道在气温18.01~29.5 ℃范围内, 纤维比强度与日平均温度呈线性正相关。 纤维强度的形成受纤维素沉积速率和时间决定,Wang 等[23]发现纤维强度与纤维素沉积速率呈负相关,而与纤维素沉积时间呈正相关。 可能在低温情况下,延长的纤维素沉积时间弥补了纤维素沉积速率下降的影响,这可能是晚播条件下纤维比强度增加的原因。 冯国艺等[24]报道在滨海盐碱地杂交棉品种(冀杂2 号)随播期推迟纤维品质提高,原因可能在于播期推迟有助于缓解对棉花生长发育的盐碱胁迫。 这与本研究适当推迟播期有助缓解生育前期低温的胁迫类似。 综上所述,晚播相对低温条件下,纤维品质略好于早播。
3.2 不同播期条件下棉铃对位叶蔗糖代谢的差异
棉花通过叶片光合作用产生同化物,棉铃对位叶为棉铃提供了其生长发育所需能量的80%以上[25]。 温度过高或过低均会导致棉铃对位叶碳水化合物合成及转运受到抑制,从而降低铃重[3,26]。本研究发现,2 年间晚播相对低温条件下A705和A201 棉铃对位叶蔗糖浓度均表现高于早播,即晚播条件下棉铃对位叶有潜力供给棉铃更多的蔗糖,从而提高铃重。 这与Liu 等[6]的研究结果相反,他们认为晚播蔗糖浓度较高的原因是低温阻碍了棉铃对位叶内蔗糖向外转运,导致铃重降低,可能是由于其研究中晚播所处温度过低。 在其他植物中也发现蔗糖随温度降低而累积[27-28]。本研究发现2 年在晚播条件下SS 酶活性降低,这与Liu 等[6]的研究一致。 晚播低温抑制SS 酶活性,植物中的SS 优先催化蔗糖的分解[8,29],晚播蔗糖分解减少,可能是晚播棉铃对位叶中蔗糖积累较多的原因之一。 此外,本研究还发现晚播棉铃对位叶中CWIN 酶活性也降低,其主要功能是在植物韧皮部的源库两端影响蔗糖的装运与卸载,从而调节碳的分配[30],这可能是晚播棉铃对位叶蔗糖浓度较高的又一原因。 综上所述,晚播相对低温下抑制棉铃对位叶SS 和CWIN 酶活性,因而减少了蔗糖的分解,使晚播条件下棉铃对位叶有更多的蔗糖向棉铃转运,这可能是晚播铃重增大和纤维品质提高的主要原因。

表5 播期对棉铃对位叶VIN,CWIN,SPS 和SS 活性的平均值和最大值的影响(2016―2017)Table 5 Effects of planting date on the mean and maximum of VIN, CWIN, SPS and SS activities in the leaf subtending to cotton boll(2016―2017)
4 结论
晚播相对低温条件下, 棉铃发育时间延长,铃重、单铃种子质量和籽指提高,衣分下降;晚播纤维长度增加,马克隆值下降;晚播低温降低了棉铃对位叶CWIN 和SS 酶活性,蔗糖浓度升高,晚播棉铃对位叶有更多的蔗糖向棉铃转运,因而有利于铃重增大和纤维品质提高。 上述研究可以为生产上合理调节播种期,提高纤维品质提供参考依据。
猜你喜欢
天津音乐学院学报(2022年2期)2022-07-28 02:32:12
河南科技学院学报(自然科学版)(2021年6期)2021-11-15 05:43:06
棉花学报(2021年1期)2021-02-05 09:37:16
广西糖业(2020年3期)2020-09-25 08:32:44
中国棉花(2019年3期)2019-03-27 06:56:14
浙江工业大学学报(2017年5期)2018-01-22 02:03:33
中国糖料(2016年1期)2016-12-01 06:49:06
印制电路信息(2015年6期)2015-12-30 12:57:52
当代音乐(下旬刊)(2015年5期)2015-05-30 05:21:05
中国果业信息(2015年11期)2015-01-23 19:19:58
