
背角无齿蚌超氧化物歧化酶基因的克隆及多溴联苯醚-47和多溴联苯醚-209对其表达的影响
2020-04-08 02:22陈佳炜邵向阳黄天科李媛张龙慧董艳美王梦琪张科齐金旭夏西超邱茂林
生态毒理学报 2020年6期
陈佳炜,邵向阳,黄天科,李媛,张龙慧,董艳美,王梦琪,张科,齐金旭,夏西超,邱茂林
平顶山学院医学院,平顶山 476000
多溴联苯醚(PBDEs)是一类广泛应用于电子设备、塑料、纺织品和建筑材料中的阻燃剂,已被列入具有生态风险性的持久性有机污染物[1-2]。随其应用不断增加,PBDEs在环境中呈现持久性累积和生物体内大量蓄积的趋势,已成为威胁淡水生物的重要有机污染物[3]。PBDEs很难被环境中微生物所降解,容易在脂肪组织中累积下来,并且随整个食物链出现放大效应[3]。PBDEs在生物体中能够催化活性氧(ROS)生成,干扰线粒体呼吸作用,导致ROS过量生成,三磷酸腺苷(ATP)合成减少,最终导致细胞死亡[3]。在PBDEs家族中,PBDE-47和PBDE-209应用较为广泛,在鳗鲡(Anguillajaponica)、海鳟(Salmotrutta)和贻贝(Mytilusedulis)等多种水生生物中被广泛检测到[4]。
超氧化物歧化酶(SOD)是生物机体防御过氧化损伤系统的关键酶之一,将氧自由基快速歧化为普通分子氧和过氧化氢,是一类敏感的分子生态毒理学指标[5]。按其所含金属辅基不同,SOD可分为铜/锌SOD(Cu/ZnSOD)、锰SOD(MnSOD)、铁SOD(FeSOD)和镍SOD(NiSOD)[6]。Cu/ZnSOD和MnSOD主要存在于原核生物、甲壳动物、鱼类和哺乳动物,其中,Cu/ZnSOD与机体正常生理功能实现和疾病发生有密切关系,也是为揭示机体氧化应激过程而研究最多的一种酶[7]。Cu/ZnSODmRNA表达和酶活性常被用作环境污染潜在标志物,尤其对重金属、杀虫剂和持久性有机污染物等环境应激改变比较敏感[8-9]。
双壳类是软体动物一个重要类群,常年栖息在海洋、河流和湖泊底部,以滤食生活为主,是检测环境污染的重要生物标志物[10-11]。相对于海洋双壳类动物而言,淡水贝类用于环境检测和毒理学研究的步伐较为滞后。背角无齿蚌是双壳类软体动物主要成员之一,在多氯酚、重金属和多氟烷基化合物等检测中发挥积极作用,常作为淡水污染的指示性生物[12-13]。为了更好揭示PBDEs环境毒性,保护淡水资源和淡水生物,本研究以背角无齿蚌(Anodontawoodiana)为研究对象,克隆出AwSOD全基因序列,分析PBDE-47和PBDE-209对AwSOD时空表达的影响,为揭示PBDEs毒理效应提供理论参考。
1 材料与方法(Materials and methods)
1.1 材料
背角无齿蚌购自南阳市水产市场,PBDE-47、PBDE-209和二甲亚砜(dimethyl sulfoxide, DMSO)购自Sigma-Aldrich公司,PBDE-47和PBDE-209溶解于DMSO中以制备储备液。TRIzol试剂、M-MLV反转录酶、菌株DH5α、克隆载体pMDl9-T、连接酶、PCR产物回收纯化试剂盒和RACE试剂盒均购自TaKaRa公司,其余常规药品均为进口或国产分析纯。
背角无齿蚌壳长(6.5±0.5) cm,PBDE-47和PBDE-209染毒之前,动物置于实验室自动水循环系统中适应养殖2周,期间停止进食。随后,动物处理实验在长方形的塑料盒(40 cm×25 cm;10 cm高)中进行,每个盒子里面8只河蚌,饲养采用人工模拟池塘水(每1 L去离子水中含48 mg NaHCO3、33 mg CaCl2·2H2O、60 mg MgSO4·7H2O和0.5 mg KCl),喂食小球藻(Chlorellavulgaris)。为了确定AwSOD基因的组织分布,对来自同一塑料盒内5只动物进行解剖,取斧足、鳃、肝胰脏、闭壳肌、心脏、血淋巴和外套膜等组织。参考PBDE-47和PBDE-209对淡水生物水蚤毒性效应研究结果[14],动物处理实验过程中,动物分为对照组、PBDE-47处理组(6.25、12.5、25、50和100 μg·L-1)和PBDE-209处理组(10、20、40、80和160 μg·L-1),对照组用同体积DMSO处理,水中DMSO浓度不超过3‰[12]。分别在0、6、12、24和48 h每组中取出5只河蚌,解剖肝胰脏、鳃和血淋巴,液氮速冻,于-80 °C保存。
1.2 总RNA提取与cDNA模板制备
按照试剂盒说明书的要求,使用TRIzol法(Takara,大连)提取总RNA。用1.0%琼脂糖凝胶电泳检测RNA质量。用M-MLV第一链cDNA合成试剂盒(Takara,大连)合成第一链cDNA,作为PCR扩增反应的模板。
1.3 AwSOD全基因序列的克隆
根据双壳纲、腹足纲、昆虫纲、甲壳纲和脊椎动物在内其他物种的Cu/ZnSOD保守区域设计简并引物SOD1和SOD2,用于扩增AwSODcDNA片段,将PCR产物克隆到pMDT-19(Takara,大连)、采用双向测序,鉴定为Cu/ZnSOD部分序列。以部分cDNA序列中设计特异引物(表1),根据RACE试剂盒说明书,采用巢式PCR方法扩增AwSODcDNA的5’和3’区域。扩增产物克隆载体、双向测序、序列比对和拼接。
1.4 序列与系统发育的分析
在GenBank数据库(www.ncbi.nlm.nih.gov/blast)中用BLAST方法对AwSOD序列进行了比对和分析;采用DANMEN软件对AwSOD基因进行多序列比对;采用信号肽预测数据库(http://www.cbs.dtu.dk/services/SignalP)对AwSOD信号肽序列进行预测;采用SMART数据库(http://smart.embl-heidelberg.de/)对AwSOD蛋白质二级结构域进行预测;采用SWISS模型(http://swissmodel.expasy.org/)对AwSOD的三维结构进行预测;用MEGA 5.0邻位连接方法,构建AwSOD系统进化树。
1.5 real-time PCR定量检测 AwSOD的表达
为了确定AwSOD转录水平,采用SYBR Premix Ex TaqTM试剂盒并按照说明书的要求进行定量分析。根据已有贝类内参基因研究结果[15-16],选取β-actin作为内参基因,根据内参基因和AwSOD特异引物分别分离对应序列(表1),琼脂糖凝胶电泳仅检测出一个扩增条带,PCR产物回收、测序和鉴别;结合溶解曲线和扩增曲线结果确定内参基因和目的基因表达的特异性和高效性。使用ABI7500实时检测系统(Applied Biosystems,美国),采用两步法,进行real-time PCR,构建标准曲线,通过2-△△CT分析AwSOD表达水平。

表1 本研究中使用的引物的描述Table 1 Description of the primes used in this study
1.6 统计学处理
PBDE-47和PBDE-209处理后AwSOD的表达水平的显著性差异采用单向方差分析(analysis of variance, ANOVA),P<0.05为差异有统计学意义。
2 结果(Results)
2.1 背角无齿蚌 AwSOD分子结构
背角无齿蚌AwSODcDNA全长由949个核苷酸组成(GenBank No., KU363382),包含一个83 bp的5’非翻译区(UTR)、401 bp的3’UTR。开放阅读框由465 bp核苷酸组成,编码155个氨基酸的多肽链,分子量为15.78 kDa,理论等电点为30.22(图1)。终止信号(AATAAA)位于3’UTR的891~896处。AwSOD与其他的细胞质Cu/ZnSOD序列比对表明,AwSOD包含Cys-7、Cys-57和Cys-146这3个半胱氨酸,其中Cys-57和Cys-146在所有Cu/ZnSOD中都是保守的,证实与分子内二硫键的形成有关(图2)。在AwSOD中,铜结合保守氨基酸残基分别为His-46、His-48、His-64和His-120,锌结合保守氨基酸残基分别为His-64、His-72和His-81及Asp-8。AwSOD氨基酸序列中存在2个高度保守Cu/ZnSOD标签序列,分别为GKHGFHVHEFGDNT和GNAGARSACGVI(图2)。

图1 背角无齿蚌 AwSOD基因的cDNA序列和推导的氨基酸序列注:粗体标示起始(ATG)和终止(TAA)密码;波浪线标示终止信号“AATAAA”;下划线标示 AwSOD标签序列;灰色阴影标示Cu和Zn结合保守位点;方框标示半胱氨酸残基(Cys-57 and Cys-146)。Fig. 1 The cDNA nucleotide sequence and the deduced amino acid sequence of AwSOD gene of Anodonta woodianaNote: The start codon (ATG) and stop codon (TAA) are indicated in bold; putative polyadenylation signal “AATAAA” is showed with wavy line; the signal sequences of AwSOD are underlined; the binding residues of Cu and Zn are marked with shadow; two cysteines (Cys-57 and Cys-146) formed a disulphide bond are marked with boxs.

图2 背角无齿蚌AwSOD与其他物种SOD序列多重比对注:红色边框标示CuZnSOD标签序列;阴影标示Cu(His-46, His-48, His-64和His-120)和Zn(His-64, His-72, His-81和Asp-84)结合保守位点;下划线标示半胱氨酸残基(Cys-57和Cys-146)。Fig. 2 Multiple alignment of AwSOD of Anodonta woodiana with other species SODsNote: Two signature motifs sequences of CuZnSOD are marked as red box; the amino acids required for Cu (His-46, His-48, His-64, and His-120) and Zn (His-64, His-72 and His-81 and Asp-84) binding are shaded; two cysteines (Cys-57 and Cys-146) formed a disulphide bond are showed with double lines.
AwSOD二级结构和三级结构与Cu/ZnSOD具有较高的相似性,二级结构包含9个β-折叠和2个α-螺旋,其中有Cys-57和Cys-146形成二硫键(图3)。
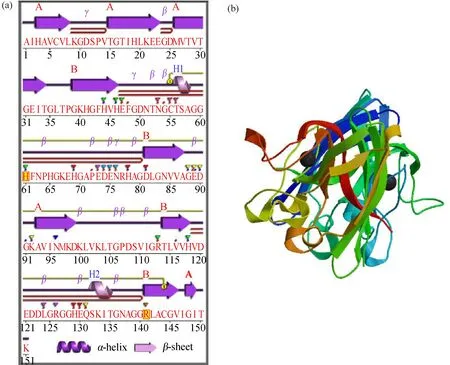
图3 背角无齿蚌AwSOD二级和三级结构预测注:(a) AwSOD二级结构;(b) AwSOD的三级结构。Fig. 3 Predicted secondary and tertiary structures of AwSOD deduced amino acids of Anodonta woodianaNote: (a) the secondary structure of AwSOD; (b) the tertiary structure of AwSOD.
2.2 AwSOD系统发育分析
BLAST分析表明,AwSOD氨基酸序列与Cu/ZnSOD具有较高的同源性,与褶纹冠蚌的同源性为97.42%,与海兔的同源性为61.39%,与斑马鱼的同源性为64.10%,与人的同源性为62.18%。为了研究AwSOD与其他物种Cu/ZnSOD之间亲缘关系,采用MEGA 5.0近邻连接法构建系统进化树,分别从不同脊椎动物和无脊椎动物中选出了多个Cu/ZnSOD序列,其中包括人、家鼠、非洲爪蟾、斑马鱼、凡纳滨对虾、果蝇、光滑双脐螺和牡蛎等。AwSOD与淡水贝类亲缘关系最近,其次海洋双壳类,最后是脊椎动物、腹足类和甲壳类(图4)。

图4 根据背角无齿蚌AwSOD氨基酸序列使用邻接法构建的系统进化树Fig. 4 Phylogenetic relationship of AwSOD amino acid sequence between Anodonta woodiana and other organisms according to neighbor-joining method
2.3 AwSOD的组织分布
Real-time PCR结果显示,背角无齿蚌AwSOD在斧足、鳃、肝胰脏、闭壳肌、外套膜和心脏壳中广泛表达(图5)。AwSOD在肝胰脏中的mRNA表达水平较高,在鳃和斧足为中等水平,在外套膜、闭壳肌和心脏中表达水平较低(图5)。

图5 背角无齿蚌不同组织中 AwSOD基因的表达(n=5)Fig. 5 Real-time PCR analysis of AwSOD transcript from different tissues of Anodonta woodiana (n=5)
2.4 PBDE-47和PBDE-209对肝胰腺 AwSOD表达的影响
在浓度6.25、12.5、25、50和100 μg·L-1的PBDE-47处理组中,肝胰腺中AwSODmRNA水平显著增加,这种上调效应在浓度6.25、12.5和25 μg·L-1PBDE-47处理组呈现时间和剂量依赖模式;与对照组相比,整个实验过程中,AwSODmRNA水平在6.25、12.5和25 μg·L-1PBDE-47处理组中分别增加了70.58%(P<0.05)、2.10倍(P<0.01)和2.61倍(P<0.01)以上(图6)。12.5 μg·L-1和25 μg·L-1的PBDE-47处理组中AwSODmRNA水平分别增加了3.57倍(P<0.01)和2.56倍(P<0.01)(图6)。
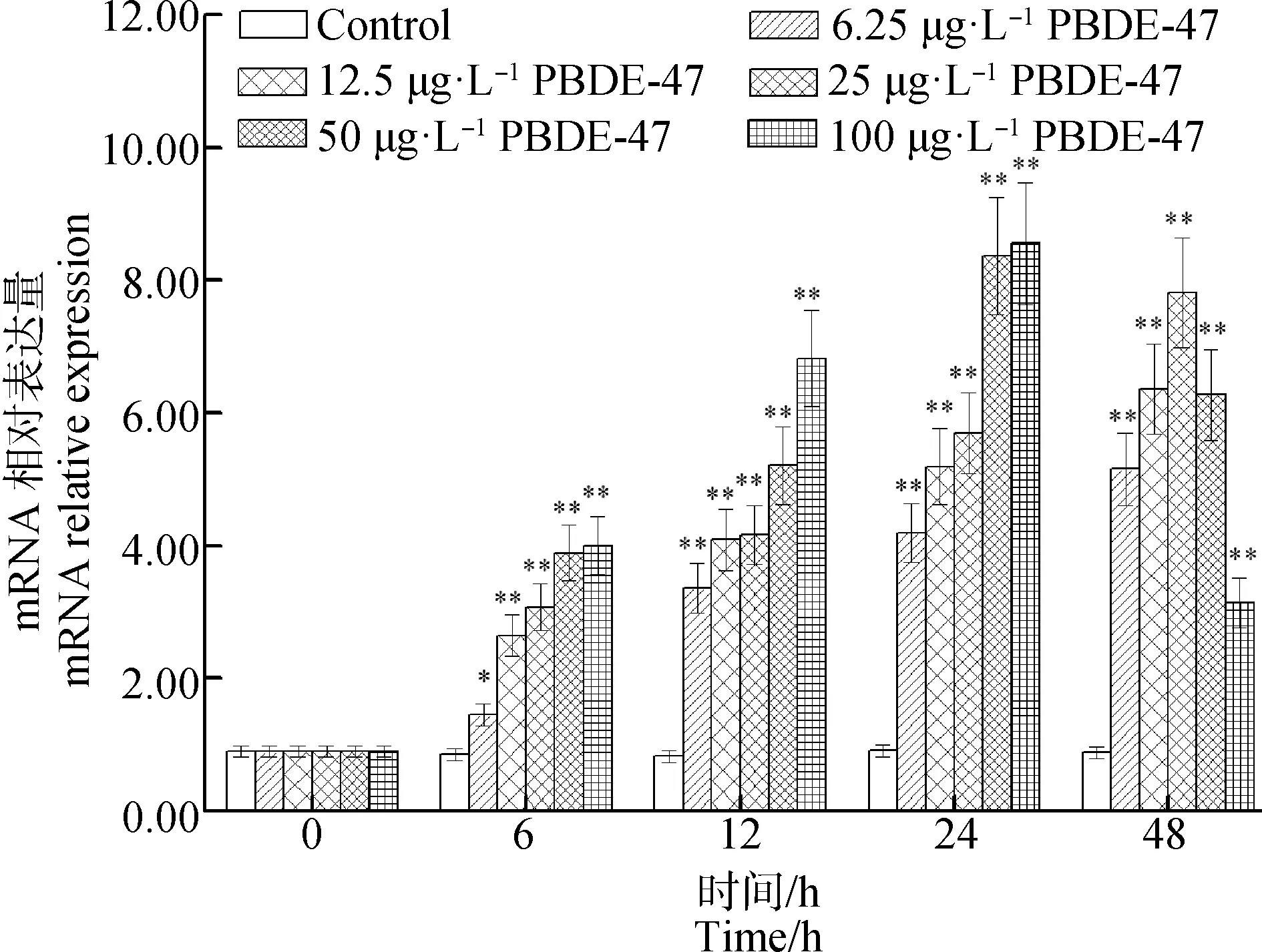
图6 PBDE-47对背角无齿蚌肝胰腺 AwSOD基因表达的影响注:每处理组的每个时间点n=5;*、**表示与相应对照组相比有显著差异( P<0.05、 P<0.01);下同。Fig. 6 Temporal expression of AwSOD in the hepatopancreas of Anodonta woodiana after PBDE-47 challenge as measured by quantitative real-time PCRNote: Data are expressed as means±SE; n=5 in each group at each time point; *, **represent P<0.05, P<0.01, compared with control group at the same time.
与对照组相比,PBDE-209处理后肝胰腺中AwSODmRNA水平随时间和剂量呈现增加趋势。在10、20、40、80和160 μg·L-1的处理组中,AwSOD表达水平分别增加了1.03%、77.08%(P<0.05)、91.66% (P<0.05)、1.33倍(P<0.05)和2.60倍(P<0.01) (图7)。

图7 PBDE-209对背角无齿蚌肝胰腺 AwSOD基因表达的影响Fig. 7 Temporal expression of AwSOD in the hepatopancreas of Anodonta woodiana after PBDE-209 challenge as measured by quantitative real-time PCR
2.5 PBDE-47和PBDE-209对鳃 AwSOD表达的影响
在浓度6.25 μg·L-1和12.5 μg·L-1的PBDE-47处理组中,鳃中AwSODmRNA水平显著增加;与对照组相比,整个实验过程中AwSODmRNA水平在6.25 μg·L-1和12.5 μg·L-1的PBDE-47处理组中分别增加了1.23倍(P<0.05)和1.64倍(P<0.05)以上(图8)。25 μg·L-1的PBDE-47处理组中,鳃中AwSODmRNA水平在24 h和48 h降至正常水平。25、50和100 μg·L-1的PBDE-47处理组中,鳃中AwSODmRNA水平在24 h和48 h低于正常水平,分别减少了46.96%和72.61%(P<0.05) (图8)。

图8 PBDE-47对背角无齿蚌鳃 AwSOD基因表达的影响Fig. 8 Temporal expression of AwSOD in the gill of Anodonta woodiana after PBDE-47 challenge as measured by quantitative real-time PCR
与对照组相比,PBDE-209处理组鳃中AwSODmRNA水平显著增加。在10、20、40、80和160 μg·L-1的处理组中,AwSOD表达水平分别增加了68.68%(P<0.05)、86.87%(P<0.05)、1.28倍(P<0.05)、1.54倍(P<0.01)和1.75倍(P<0.01)(图9)。

图9 PBDE-209对背角无齿蚌鳃 AwSOD基因表达的影响Fig. 9 Temporal expression of AwSOD in the gill of Anodonta woodiana after PBDE-209 challenge as measured by quantitative real-time PCR
3 讨论(Discussion)
本研究结果显示,AwSOD中未发现信号肽序列,提示AwSOD属于胞质Cu/ZnSOD家族成员,已经过加工并具备了成熟模式。AwSOD氨基酸序列与其他Cu/ZnSODs序列多重比对表明,2个半胱氨酸残基Cys-57和Cys-146与CuZnSODs氨基酸残基具有高度保守性,可能与二级结构中二硫键的形成有关[17-18]。在AwSOD和其他Cu/ZnSODs序列中,Cu和Zn结合氨基酸残基相对比较保守,这些保守的氨基酸有助于AwSOD结构稳定性和催化的作用[19]。同时,在相对恶劣环境条件下,这些氨基酸残基对稳定Cu/ZnSODs构象具有潜在作用,有助于Cu和Zn稳定Cu/ZnSOD三级结构[20]。
AwSOD在不同组织中呈现不同的表达模式,其中,在肝胰腺组织中AwSODmRNA的表达水平最高,这与肝胰脏作为主要代谢组织和主要防御器官有关。相同的现象在南美白对虾(Litopenaeusvannamei)和拟穴青蟹(Scyllaparamamosain)中也被发现[21-22]。相当于哺乳动物的肝脏和昆虫的脂肪体而言,水生无脊椎动物肝胰腺具有肝脏和胰腺双重功能,不仅是重要的消化器官,而且在非特异性免疫中发挥着重要的作用[21-22]。肝胰腺具有较高的代谢活性,水体中污染物进入机体后通过代谢过程会产生大量的ROS。AwSOD基因高表达提示其可作为一种重要的肝脏解毒酶。其他组织中AwSOD的广泛分布有助于将氧自由基快速转化为普通分子氧(O2)和过氧化氢(H2O2)。
研究发现,PBDE-47和PBDE-209处理后背角无齿蚌肝胰腺和鳃AwSOD表达水平显著升高,提示这可能与增强机体ROS清除能力和提高胁迫耐受性有关。伴随PBDE-47和PBDE-209在机体中的不断累积,细胞中ROS生成增加,诱发细胞氧化应激,线粒体出现呼吸爆发,细胞出现功能性受损。在胁迫条件下,生物本身SOD、过氧化氢酶、谷胱甘肽S转移酶和硫过氧化物酶表达水平提升,以提高机体清除ROS能力,维持机体稳态。长时间Cd暴露后,非洲爪蟾(Xenopuslaevis)和青鳉(Oryziasjavanicus)Cu/ZnSODmRNA表达水平显著上调[23]。白斑杆状病毒染毒后,日本囊对虾(Marsupenaeusjaponicus)鳃和血淋巴中Cu/ZnSOD表达水平显著升高[24]。乳球菌处理后,罗氏沼虾(Macrobrachiumrosenbergii)血淋巴中Cu/ZnSOD表达水平显著增加[25]。马拉硫磷、硫丹和二者混合物处理后,南美白对虾(L.vannamei)血淋巴中Cu/ZnSOD表达水平显著增加[26]。Cd2+胁迫后,褶纹冠蚌(Cristariaplicata)Cu/ZnSOD基因mRNA水平和酶活性迅速升高,并在72 h和48 h达到峰值,提示Cu/ZnSOD是用于监测早期水体重金属污染的重要靶分子[27]。由此可见,Cu/ZnSOD在维持机体氧自由基代谢平衡、免受氧化损伤、ROS清除和机体保护性防御中起着重要作用。PBDE-47和PBDE-209处理后背角无齿蚌肝胰腺和鳃AwSOD表达水平显著升高与增强机体抗氧化能力和耐受性有关。
与对照组相比,不同浓度PBDE-47暴露后,肝胰腺中AwSOD在高剂量组出现先升高后降低的现象,而在鳃中,AwSOD表达水平在6 h显著上调,后期出现下调现象,提示这种表达模式与PBDE-47毒性效应和剂量效应有关。正常情况下,动物体内ROS保持相对较低的水平,ROS生成后将迅速被一系列抗氧化酶迅速清除,以维持ROS水平与抗氧化酶活性之间的平衡[28-29]。抗氧化酶是机体内稳态的重要标志之一。鳃作为贝类重要免疫器官,鳃丝直接接触PBDE-47和PBDE-209,并通过鳃丝呼吸作用迅速进入机体,产生大量ROS,相对于肝胰腺而言,PBDE-47对鳃氧化应激效应更为直接和明显。持续高剂量长时间的PBDE-47暴露,导致进入体内PBDE-47超过鳃的解毒极限,则鳃会受到损伤,大量鳃组织中细胞凋亡,处理后期出现表达水平下调现象[30]。相同现象在褶纹冠蚌中也被发现,Pb2+胁迫后,褶纹冠蚌Cu/ZnSODmRNA的表达先升高后降低,在48 h达到最大值[27]。在PBDE-209处理组中,AwSOD的表达呈时间和剂量依赖性,与所选择的浓度未达到临界值,动物在这种浓度可以产生足够的抗氧化能力,从而保持机体PBDE-209氧化和抗氧化酶还原作用之间的平衡;实验中选定浓度未检测到PBDE-209半数致死浓度进一步证实了这一现象。
研究显示,地表水中PBDE-47含量为0.25 ng·L-1,PBDE-209含量为1.0 ng·L-1[31]。软体动物调查结果显示,PBDE-47、PBDE-99、PBDE-154和PBDE-153含量为0.39~3.65 ng·g-1,PBDE-209作为主要持久性有机污染物含量为10.2~284 ng·g-1[32]。PBDE-47和PBDE-209对淡水资源的负面效应和水生生物的累积效应已成为不可忽视的问题。PBDE-47和PBDE-209对背角无齿蚌机体氧化应激作用是一个综合的效应,其产生氧化应激后抗氧化系统启动,涉及多种抗氧化酶的参与。从AwSOD组织特异性表达及PBDE-47和PBDE-209对其表达的诱导作用,背角无齿蚌是用于淡水污染监测的重要生物之一。
猜你喜欢
生物化学与生物物理进展(2022年7期)2022-07-25
生物化学与生物物理进展(2022年6期)2022-07-21
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19
中成药(2018年12期)2018-12-29
中国医学影像学杂志(2018年9期)2018-10-17
西南国防医药(2016年6期)2016-12-01
中国病理生理杂志(2015年8期)2015-12-21
天津科技大学学报(2015年2期)2015-08-09
中国应用生理学杂志(2014年3期)2014-01-22
中国民族民间医药·下半月(2011年10期)2011-12-27
