
额尔齐斯河银鲫繁殖生物学研究
2020-04-06 05:10张志明刘成杰丁慧萍谢从新马徐发
水生生物学报 2020年2期
张志明 刘成杰 丁慧萍 谢从新 马徐发
(1. 水利部中国科学院水工程生态研究所, 水利部水工程生态效应与生态修复重点实验室, 武汉 430079; 2. 华中农业大学水产学院, 武汉 430070; 3. 中国科学院水生生物研究所, 武汉 430072)
繁殖是鱼类生活史的重要组成部分, 包括鱼类的两性系统、性腺发育、洄游、产卵、受精、孵化, 以及仔、稚鱼生长和发育等一系列过程[1], 它是鱼类自然种群补充的唯一方式, 也是鱼类应对自然死亡、捕捞死亡以及迁出等引起的种群减少和维持种群发展及延续的根本[2]。繁殖生物学研究包括性比、性腺发育、初次性成熟大小、繁殖周期和时间、繁殖力等方面, 其中, 初次性成熟大小和繁殖力评估, 还是鱼类种群动力学研究的基础[3]。鱼类繁殖策略则是指鱼类在长期的自然选择过程中形成的与其生存环境条件相适应的繁殖特性, 包括生殖洄游、聚群、亲体护幼、产卵类型等, 保证鱼类及其后代能够获得最大的存活率。对自然条件下鱼类繁殖特性进行研究是鱼类人工繁育及增养殖可持续发展的前提, 也是鱼类资源保护和相关渔业管理政策制定的重要依据[4]。
银鲫(Carassius auratus gibelio)隶属于鲤形目(Cypriniformes), 鲤科(Cyprinidae), 鲤亚科(Cyprininae), 鲫属(Carassius), 我国仅分布于额尔齐斯河及黑龙江水系[5]。作为额尔齐斯河流域重要的经济鱼类, 银鲫具有生长速度快, 个体大, 环境适应能力强, 且肉质细嫩、口感好等优点, 深受广大渔民及休闲渔业爱好者的喜爱。近年来, 由于过度捕捞、水电开发、农田灌溉及水质污染等原因, 导致额尔齐斯河银鲫种群资源量呈现逐年下降的趋势, 其野生种群急需保护[6]。
目前, 有关额尔齐斯河银鲫的生物学研究方面的报道仍较为有限, 主要集中在资源量调查[7]、食性分析[8]、年龄及生长[9]、形态学比较分析[10,11]等方面。而其繁殖特性方面, 仅见于任慕莲等[7]对额尔齐斯河及其附属水体银鲫的调查性研究。因此,本文通过对额尔齐斯河银鲫种群组成、性比、性腺发育、繁殖季节、初次性成熟大小及繁殖力等繁殖生物学特性进行研究, 从而进一步丰富银鲫生物学方面的研究资料, 并为额尔齐斯河银鲫的资源养护和可持续利用提供科学依据。
1 材料与方法
1.1 样本采集
2013年4—10月, 在新疆额尔齐斯河主河道中国段下游(47°96′—48°07′N; 85°51′—85°62′E), 采用流刺网(内层11 cm, 外层23 cm)、定置刺网(网目5 cm)、拉网(网目1 cm)等渔具采集银鲫样本546尾,每月不少于30尾。样本在新鲜状态下进行标准体长(Standard length,SL)、体重(Body weight,BW)、性腺重(Gonad weight,GW)、空壳重(Eviscerated weight,EW)等常规生物学测量, 长度精确到1 mm,重量精确到0.01 g。依据性腺的形态学特征目测鉴定性别, 取微耳石作为年龄鉴定材料。
1.2 性比
对采集银鲫样本的雌鱼和雄鱼(♀/♂)比例进行统计, 并与其他地理区域的结果进行比较分析。
1.3 性腺组织学
用多聚甲醛对性腺样品进行固定, 梯度酒精脱水,梯度二甲苯透明, 石蜡包埋切片, HE染色, 甘油封片,用Nikon Eclipse 80i显微拍照系统对性腺组织切片进行观察和拍照。参考Blazer[12]和Smith等[13]将卵母细胞发育分为6个时相, 并分别根据卵母细胞和精细胞发育及比例情况对卵巢[14,15]和精巢[16]进行发育分期。
1.4 繁殖季节和产卵类型
性腺宏观发育分期参考《鱼类学》中的描述[17]。银鲫的繁殖季节依据性腺发育和成熟系数(Gonadosomatic index,GSI, %)月变化情况确定, 成熟系数计算公式为:
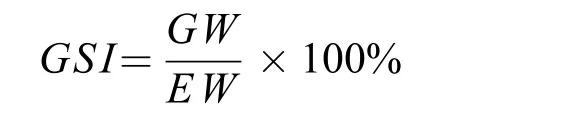
式中,GSI代表性体指数;GW代表性腺重;EW代表空壳重。
取Ⅳ期(目测法分期)卵巢部分样本, 10%甲醛溶液固定后带回实验室, 在Leica EZ4D解剖镜下对积累卵黄的卵粒进行拍照, 用Image Pro Plus v6.0软件测量卵径大小。根据卵巢组织学特征、卵径频率分布月变化情况判断产卵类型。
1.5 初次性成熟体长及年龄判定
分别按体长10 mm和年龄1龄分段, 计算各区段内的性成熟个体比例, 对性成熟个体比例和每个区段的中值进行Logistic回归[18], 公式如下:

式中,P为各体长区段的性成熟比例;r为常数;X为体长或年龄区段的中间值;b为初次性成熟体长或年龄。
1.6 繁殖测定
采用重量分析法对银鲫的繁殖力进行测定[19]。于繁殖季节前后(4—7月), 从98尾IV期(目测法分期)卵巢的前、中和后部随机称取1 g左右的子样,10%甲醛固定后带回实验室, 计数沉积卵黄的卵粒数, 采用下列公式计算获得绝对繁殖力(Absolute fecundity,AF)和相对繁殖力(Relative fecundity,RF):
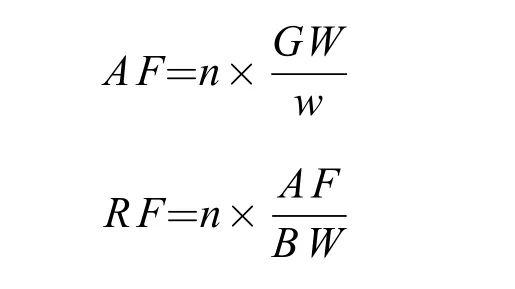
式中,AF代表绝对繁殖力;RF代表相对繁殖力;n代表计数的卵粒数;GW代表卵巢重;w代表卵巢子样重;BW代表体重。
采用回归分析法检验绝对繁殖力与体长、体重、年龄和卵巢重的关系[20]。
1.7 数据统计与分析
采用Photoshop CS2软件进行图像处理, 数据分析处理和绘图采用Microsoft Excel 2003、SPSS 19.0和OriginPro 8.5.1软件。数据采用平均数±标准差表示, 显著水平设为0.05。
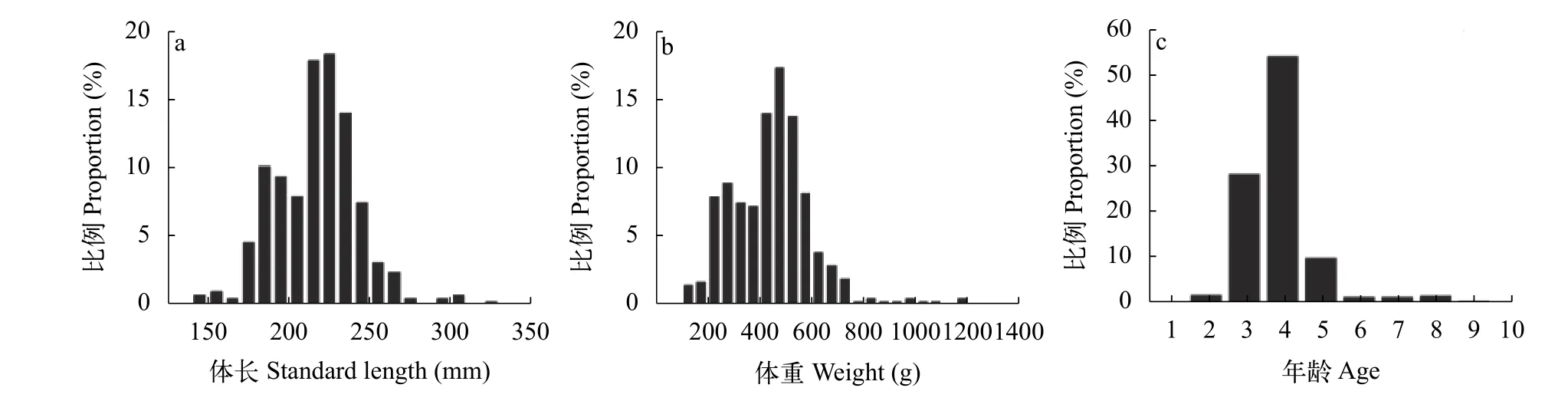
图 1 雌鱼的体长(a)、体重(b)及年龄(c)分布Fig. 1 The standard length (a), weight (b) and age structure of the female reproductive population
2 结果
2.1 群体结构及性比
据统计, 雌鱼体长分布范围为142—328 mm,优势体长组为220—240 mm, 占总体的50.3%(图 1a);体重范围为102.8—1171.6 g, 优势体重组为400—600 g, 占总体的60.8%(图 1b); 年龄范围为2—9龄,优势年龄组为3—4龄, 占总体的82.8%(图 1c)。
在546尾银鲫样本中, 雌性488尾, 雄性45尾, 性别未辩13尾, 性比(♀∶♂)为10.84∶1。
在繁殖期, 银鲫雌性个体腹部饱满膨大、柔软, 其个体明显大于雄性个体, 生殖孔圆形微凸, 呈红色; 雄性, 腹部偏硬, 个体小于雌性, 生殖孔凹陷, 略呈三角形。
2.2 卵母细胞发育分期
第1时相卵母细胞(图版Ⅰ-1 O1): 为卵原细胞向初级卵母细胞过渡的细胞。细胞质很少, 细胞核形态明显。
第2时相卵母细胞(图版Ⅰ-1 O2): 细胞进入小生长期, 细胞质和细胞核物质增多; 出现若干核仁,靠近核膜分布。
第3时相卵母细胞(图版Ⅰ-2、3): 在第2时相卵母细胞基础上, 细胞质继续增加, 该时相卵母细胞早期的特点是在靠近卵膜处出现一层液泡, 滤泡膜与卵母细胞膜之间出现辐射带(图版Ⅰ-2a); 细胞继续增大, 液泡层数增加且辐射带增厚, 此时为第3时相中期阶段(图版Ⅰ-2b); 到第3时相后期整个细胞质充满液泡, 细胞体积进一步增大, 辐射带明显增厚(图版Ⅰ-3)。
第4时相卵母细胞(图版Ⅰ-4—6), 此时的细胞变化特点在于卵黄积累程度和细胞核位置变化。早期细胞质中卵黄颗粒开始增加, 液泡层数减少,辐射带增厚(图版Ⅰ-4); 随后卵黄颗粒进一步增多,液泡变少集中于卵母细胞边缘 (图版Ⅰ-5); 晚期细胞核偏离细胞中央, 卵黄继续积累, 仍存在少量液泡(图版Ⅰ-6)。
第5时相卵母细胞(图版Ⅰ-7), 卵黄颗粒数量达到最大, 之后会逐渐融合成块状, 细胞质边缘仍存在少量液泡。
第6时相卵母细胞(图版Ⅰ-8), 主要存在于产后卵巢, 为退化吸收阶段的卵母细胞, 细胞核和卵黄等物质开始裂解。
2.3 性腺发育分期
卵巢组织学发育分期如下:
Ⅰ期: 卵巢中以进入初级卵母细胞小生长期的细胞为主, 细胞主要特点为细胞质较少, 细胞核较大(图版Ⅱ-1)。
Ⅱ期: 卵巢中出现进入大生长期的第3时相卵母细胞(O3), 卵母细胞体积增大, 细胞膜附近出现液泡和辐射带(图版Ⅱ-2)。
III期: 卵巢中的卵母细胞继续发育, 体积进一步增大, 第3时相卵母细胞数目逐渐增多, 液泡充满整个细胞质, 细胞辐射带明显增厚(图版Ⅱ-3)。
Ⅳ期: 卵巢中出现第4时相卵母细胞, 由于卵黄积累, 细胞体积增大, 同时还可观察到处于小生长期的卵母细胞(图版Ⅱ-4)。
Ⅴ期: 卵巢以处在发育晚期的第4时相卵母细胞(O4)为主, 细胞积累了大量卵黄颗粒, 逐渐过渡到成熟状态(图版Ⅱ-5)。
Ⅵ期: 产后卵巢, 有空滤泡(POF)存在, 未产出的成熟卵母细胞(O6)逐渐退化并被重吸收(图版Ⅱ-6)。
精巢组织学发育分期如下:
Ⅰ期: 精小叶以精原细胞为主, 中间充满结缔组织(图版Ⅲ-1)。
Ⅱ期: 精小叶出现空腔, 精原细胞发育为精母细胞, 有少量的精细胞出现(图版Ⅲ-2)。
Ⅲ期: 精小叶中精母细胞大量分裂生成精细胞, 生殖细胞聚集形成精小囊(图版Ⅲ-3)。
Ⅳ期: 精小叶空腔扩大, 腔中精细胞和精子数量占绝大比例(图版Ⅲ-4)。
Ⅴ期: 精小叶中精子数量继续增多, 精小叶空腔继续扩大, 精巢发育成熟(图版Ⅲ-5)。
Ⅵ期: 精子已大部分排出, 精巢中主要由精原细胞组成, 小叶中存在少量精子(图版Ⅲ-6)。
2.4 卵径频率
据统计, 卵子直径大于1.0 mm的月份主要在4月、5月、6月, 分别占总数的47.02%、51.37%和47.47%。7月份卵径主要集中在0.5—0.7 mm, 均值缩减至0.59 mm, 分布于该区间的卵子约占79.18%;9月和10月卵径均值分别为0.73和0.94 mm, 此时,银鲫卵巢经短暂的产后停滞期后开始不断发育, 其卵径较7月有所增大(图 2)。
2.5 繁殖季节
银鲫雌性性体指数(GSI)在4、5月均保持在较高水平(图 3), 分别为11.3%和14.1%, 且4月较5月偏低, 表明这一时期银鲫卵巢仍处在发育阶段或未进入繁殖高峰期; 6月份, 雌鱼性体指数出现明显的下降(7.64%), 直至7月接近最低值1.83%; 8—10月, 雌鱼性体指数(GSI)呈现缓慢上升的趋势。银鲫雄性的性体指数变化与雌鱼类似。从卵巢不同发育期比例月变化况来看(图 4), 4—6月Ⅳ期卵巢比例较大, 分别为76.36%、74.69%和51.65%; 7月则以Ⅵ期(产后期)为主, 占65.45%; 8—10月以Ⅱ期和Ⅲ期为主。根据以上分析, 额尔齐斯河银鲫繁殖期为5月持续到7月, 高峰期为6月。
2.6 初次性成熟体长及年龄
将性成熟个体比例对体长和年龄分别进行Lo-gistic回归, 获得方程如下(图 5):
体长, 雌性:P=[1+e-k(SL-161)]-1(n=488,R2=0.92);
雄性:P=[1+e-k(SL-135)]-1(n=44,R2=0.86)。
年龄, 雌性:P=[1+e-k(A-2.3)]-1(n=488,R2=0.99);
雄性:P=[1+e-k(A-1.9)]-1(n=44,R2=0.93)。
银鲫初次性成熟体长(SL50) 和年龄(A50)分别为雌鱼161 mm和2.3龄, 雄鱼135 mm和1.9龄, 渔获物中雌、雄鱼最小性成熟个体年龄均为2龄。
2.7 繁殖力
银鲫绝对繁殖力为11850—187419粒, 均值为(42453±28205)粒; 相对繁殖力为32.5—212.9粒/g,均值为(98.19±34.6)粒/g。
绝对繁殖力与体长(SL)呈线性相关性较差, 与体重(BW)呈幂指数相关性较体长优势明显, 但与年龄(Age)及卵巢重(GW)无显著相关性(图 6), 与体长和体重最佳拟合关系式如下:
绝对繁殖力与体长:F=766.24SL-131226(n=98,R2=0.564);
绝对繁殖力与体重:F=5.15W1.44(n=98,R2=0.745)。
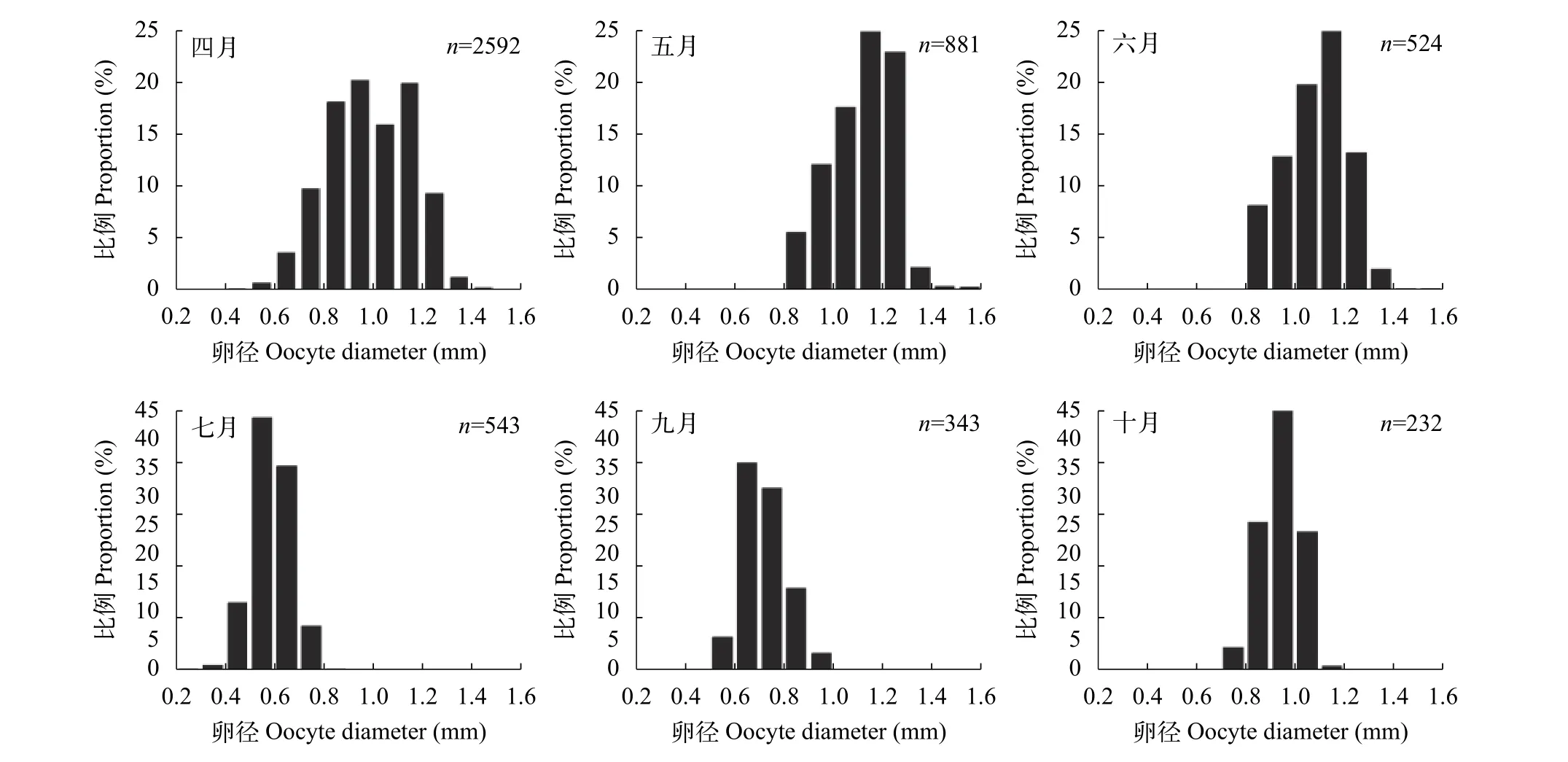
图 2 银鲫不同月份卵径频率分布图Fig. 2 Size frequency of oocytes in C. auratus gibelio in different months

图 3 银鲫各月份的性成熟系数变化趋势Fig. 3 Mean gonadosomatic index variation by month for C.auratus gibelio
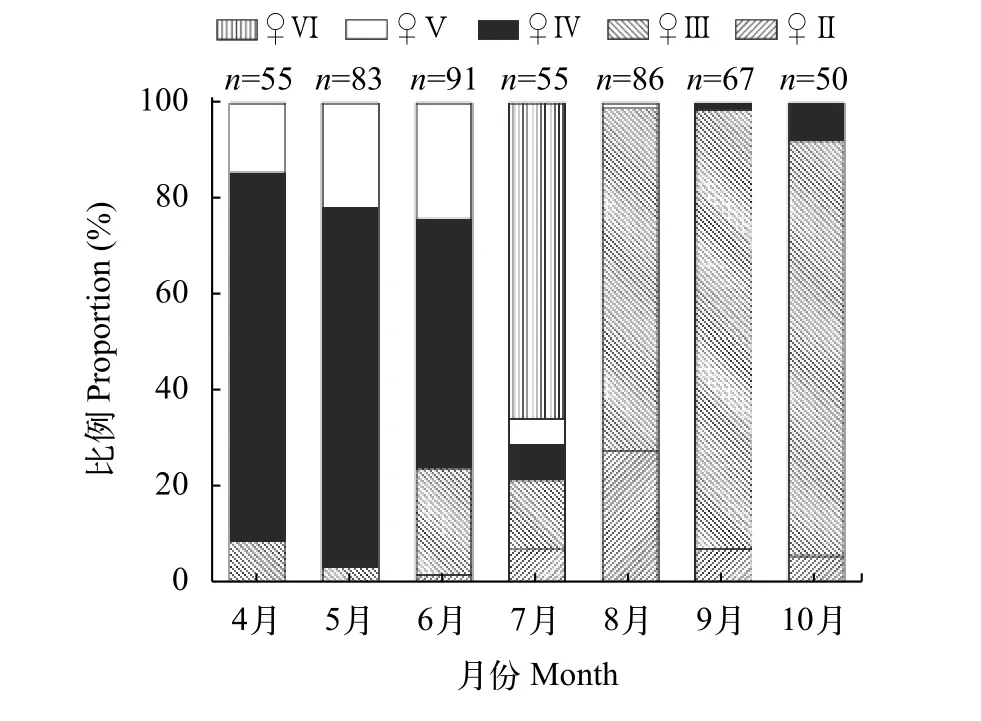
图 4 各月份银鲫卵巢各发育期比例分布Fig. 4 Variation in maturity stages of ovaries from April to October in C. auratus gibelio
3 讨论
3.1 性比
在本研究中, 额尔齐斯河银鲫雌雄性比为10.84∶1,与雅鲁藏布江鲫种群的雌雄性比(12.84∶1)相近[21],但明显小于Anzali Lagoon (141.86/1)[22]。桂建芳和周莉[23]的研究指出, 额尔齐斯河银鲫为三倍体, 具有独特的雌核发育生殖方式, 而人工繁育试验和分子标记分析又证实银鲫存在有性生殖[24], 因此, 银鲫雌性个体在种群中占比较高。李风波等[25]的研究结果表明, 额尔齐斯河水系银鲫群体中雄性个体可能占5%—10%。在本研究中, 额尔齐斯河银鲫雄性个体占比8.45%, 在该范围内。从另一方面讲, 雌性个体的数量关系到种群繁殖力的大小, 雌性个体数量上的优势使得种群具有较大的繁殖潜力, 同时特殊的繁殖方式使得银鲫能更好的适应环境变化,能够更好的使种群得以生长繁衍, 并确保在采取适当保护措施的情况下, 银鲫种群资源量可在较短时间内达到相对较高的水平。
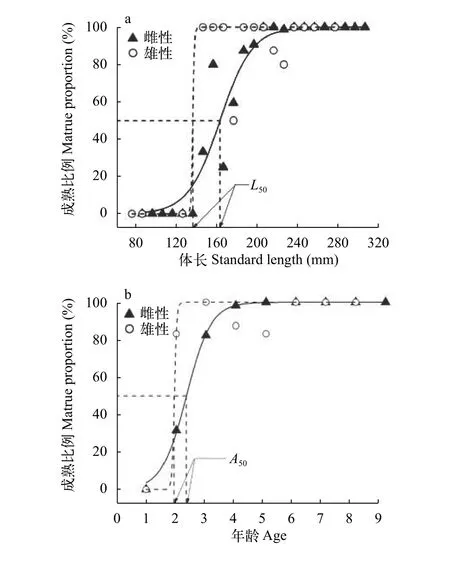
图 5 银鲫性成熟体长(a)和年龄(b)比例Logistic回归Fig. 5 Logistic functions fitted to proportion of sexually mature individuals in terms of length (a) and age (b) for C. auratus gibelio

图 6 银鲫绝对繁殖力与体长(a)、体重(b)、年龄(c)和卵巢重(d)的关系Fig. 6 Regressions between absolute fecundity and standard length (a), body weight (b), age (c), and ovary weight (d) in C. auratus gibelio
3.2 产卵类型分析
施瑔芳[26]根据卵巢中卵母细胞是否存在不同发育阶段, 将鱼类的产卵类型划分为三种: 分别为完全同步型、分批同步型和分批非同步型。在银鲫的Ⅳ期卵巢中, 有少量Ⅱ、Ⅲ时相与Ⅳ时相卵母细胞同时存在的现象(图版Ⅱ-4), 而在Ⅴ期卵巢中,卵母细胞大部分形态相似, 发育阶段基本同步(图版Ⅱ-5)。此外, 分批产卵的鱼类一般可发现其卵巢切片中产后滤泡和发育到Ⅳ期的卵母细胞会同时存在, 在银鲫的Ⅵ期卵巢中可以同时观察到产后滤泡和第Ⅲ、Ⅳ时相的卵母细胞(图版Ⅱ-6), 但是观察到Ⅳ时相的卵母细胞细胞膜破裂不规则, 处于产后退化状态, 无法产出; 第Ⅲ时相的卵母细胞到第Ⅳ时相和第Ⅴ时相需要积累大量的卵黄, 在短时间内也无法产出。谢从新[17]研究指出, 不分批产卵的鱼类性体指数(GSI)在产前达到最高峰, 产后迅速跌落, 而分批产卵的鱼类其性体指数(GSI)不会出现剧烈变化。银鲫性体指数(GSI)在5月达到高峰, 而在6—7月份又迅速跌落到最低值。不分批产卵鱼类卵径分布均匀, 呈单峰型, 而分批产卵鱼类卵径分布不均匀, 呈多峰型[27]。而在本研究结果中, 银鲫卵径频率分布(图 6)呈单峰型。因此, 综上分析, 推测额尔齐斯河银鲫为分批同步产卵类型。
3.3 繁殖时间
鱼类繁殖时间需要适宜的水温, 较低的水温会使繁殖开始的时间推迟[28]。额尔齐斯河银鲫繁殖季节要明显晚于太湖[29]、草海[30]、洞庭湖及洪湖[31]四个水体的鲫(表 1), 主要原因是额尔齐斯河处于冰封期从11月延续到翌年4月, 4月水体解冻水温上升, 导致额尔齐斯河银鲫繁殖时期比其他四个水域要晚。同时在4月水温开始上升, 水体中饵料丰度开始提高[6], 银鲫选择该季节(5—7月)进行繁殖, 能够使后代获得充足的食物来源和适宜的生长水温,从而提高后代的存活率。
3.4 不同种群初次性成熟年龄比较分析
估算的额尔齐斯河银鲫雌鱼和雄鱼初次性成熟年龄分别为2.3龄和1.9龄, 雌、雄鱼最小性成熟年龄为2龄, 与太湖[29]、草海[30]、洞庭湖、洪湖[31]、梁子湖[32]及衡水湖[33]的鲫种群相比性成熟较晚(表 1),分析其原因可能是额尔齐斯河流域纬度较高, 具有气温低、冬季漫长等气候特点, 适合鱼类生长的时间短, 而其他几个水体适合鱼类生长的适温时间长,因此比额尔齐斯河银鲫达到性成熟的年龄相对较小; Grover[34]研究发现, 野生群体中温度和性别成正相关。同时, 延迟性成熟时间, 也可以看作是银鲫对新疆恶劣环境适应的一种表现。

表 1 不同水域鲫属鱼类繁殖特性比较Tab. 1 Comparison of the reproductive biology characteristics ofCarassius fish from different water bodies
3.5 不同种群繁殖力比较分析
种群密度、个体营养条件和环境因子等因素会对鱼类繁殖力产生一定的影响[4]。对额尔齐斯河银鲫种群繁殖力与个体生物学指标相关性研究表明绝对繁殖力与个体的年龄相关性不显著, 而与体长、体重呈显著的正相关, 同时研究发现相同年龄,不同体长、体重个体之间的繁殖力差异较大, 在相同的水域环境条件下, 个体之间营养积累程度的差异会对个体繁殖力大小产生一定的影响。与其他水域的鲫种群相对繁殖力进行比较, 结果表明额尔齐斯河银鲫相对繁殖力较小(表 1)。相对繁殖力体现了个体的繁殖策略, 相对繁殖力低意味着鱼卵体积大, 卵黄积累的多, 从而使每个卵发育成为成体的机会增加[24]。额尔齐斯河银鲫的绝对繁殖力除比梁子湖[32]和达里湖[35]的鲫种群小以外, 比其他水域的要大(表 1), 体现出较大的繁殖潜力, 这是银鲫在额尔齐斯河寒温带气候条件和高纬度条件下保证的物种的延续和繁衍所选择的繁殖策略。

图版 Ⅰ 额尔齐斯河银鲫卵母细胞发育分期Plate Ⅰ Different developmental stages of oocytes in C. auratus gibelio from the Irtysh River
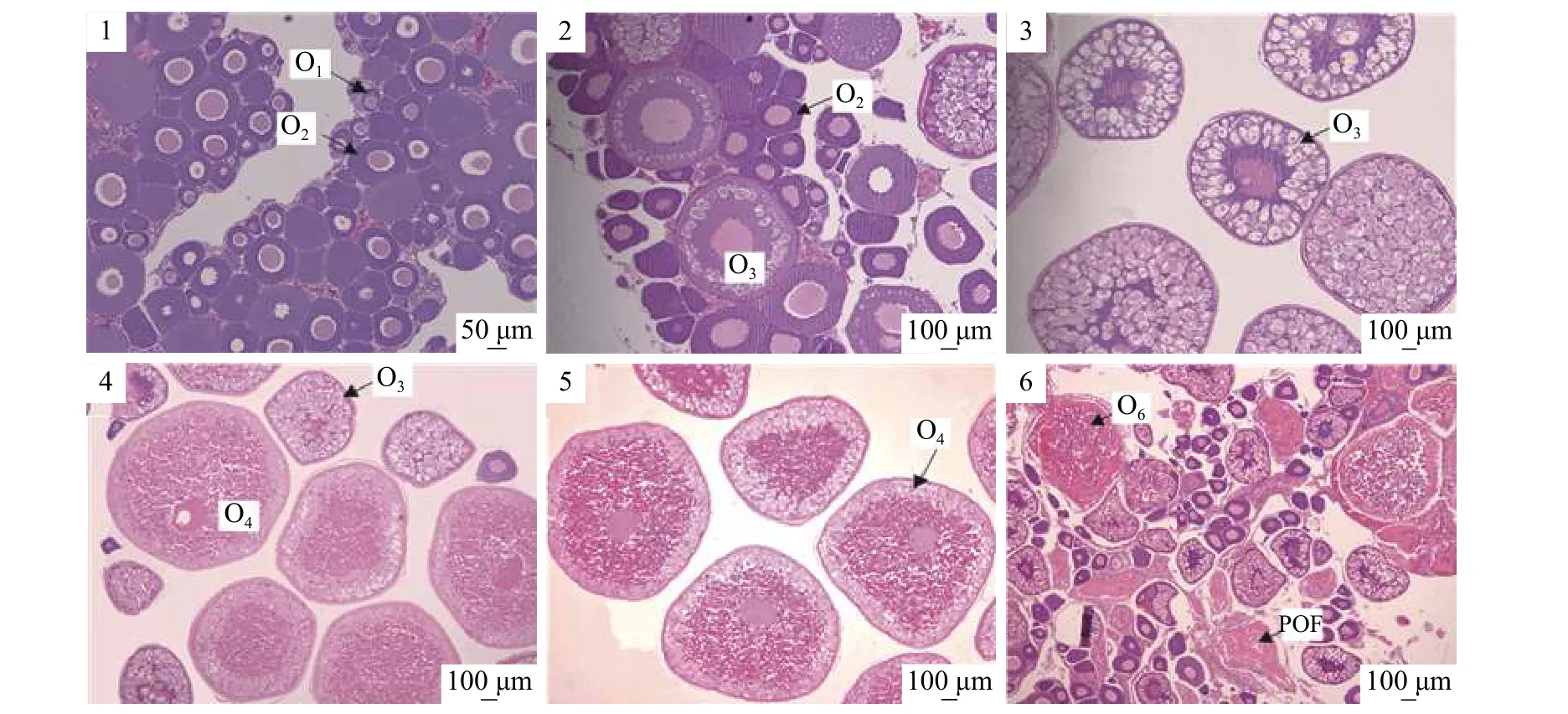
图版 Ⅱ 额尔齐斯河银鲫各时期卵巢Plate Ⅱ Different developmental stages of ovaries in C. auratus gibelio from the Irtysh River

图版 Ⅲ 额尔齐斯河银鲫各时期精巢Plate Ⅲ Different developmental stages of testes in C. auratus gibelio from the Irtysh River
猜你喜欢
黑龙江水产(2022年2期)2022-05-09
海洋渔业(2021年1期)2021-03-18
水生生物学报(2021年1期)2021-02-04
河北渔业(2019年5期)2019-07-08
农业与技术(2018年14期)2018-11-12
猪业科学(2018年4期)2018-05-19
湖北畜牧兽医(2018年1期)2018-01-26
中国三峡(2016年4期)2016-05-26
新疆人文地理(2016年7期)2016-05-14
中药与临床(2015年5期)2015-12-17
