
青藏高原典型草地植被退化与土壤退化研究
2020-03-27 09:24郝爱华尤全刚段翰晨黄翠华董斯扬
生态学报 2020年3期
郝爱华,薛 娴,彭 飞,尤全刚,廖 杰,段翰晨,黄翠华,董斯扬
1 中国科学院西北生态环境资源研究院沙漠与沙漠化重点实验室, 兰州 730000 2 中国科学院大学, 北京 100049
作为地球“第三极”,青藏高原拥有世界上最大的高寒草地生态系统,草地面积约16.538×105km2[1],占中国草地面积的41.88%,世界草地面积的6%[2]。然而,近30年来青藏高原草地退化严重,退化面积约占高原草地总面积的38.8%[3]。随着草地退化,植被盖度降低[4],群落结构和物种组成发生改变[5],土壤有机碳和总氮严重流失[6],土壤持水能力下降[7],表层土壤干旱[8- 9], 生态系统稳定性明显降低。小尺度上景观异质性增强[10],尤其是高寒草甸“斑块状”退化已成事实[11]。草地退化已严重威胁青藏高原生态环境、生物多样性保护和畜牧业经济发展[12]。
高寒草原和高寒草甸是青藏高原最主要的两种植被类型,属于典型的“高原地带性”植被[13]。高寒草原是在寒冷、干旱的亚高山、高山地区,由冷旱生密丛型多年生禾草、青藏苔草等为建群种,植被稀疏,垂直分层结构明显的草原群落[14]。高寒草甸是草甸植被中适高寒生境的类型,以耐寒适寒的中生多年生草本植物为建群种,草群密集,草层低矮,无明显层次分化的群落类型;高寒草原主体分布在羌塘高原中部、东昆仑山原及长江源区。高寒草甸在藏东、藏南、青海南部和祁连山东段高山带广泛发育[15]。目前针对青藏高原高寒草原和高寒草甸植被退化和土壤退化研究成果丰硕。高寒草原研究集中于三江源紫花针茅群落[16- 17]、青藏高原腹地青藏苔草草原[18]和青藏高原东部沙化高寒草原[19]等;高寒草甸研究集中于甘肃省玛曲县[20], 青海省果洛藏族自治州[21- 22],藏北那曲县[23]和青藏高原东北部海北站[24]等。Wang等[25]认为高寒草甸退化过程中群落物种组成改变,物种数减少,土壤肥力下降,土壤0—10cm根茎比例增加;周华坤等[16]发现高寒草原退化过程中,禾草地上生物量显著减少,杂类草地上生物量先增加后减少,莎草地上生物量不受退化影响;土壤湿度减小,土壤肥力下降。然而,很少有研究将青藏高原高寒草原和高寒草甸植被退化与土壤退化特征差异进行对比,这将不利于退化高寒草地的恢复治理和管理调控[16]。为此选取青藏高原典型高寒草原和典型高寒草甸为研究对象,对草地退化过程中有关植被群落结构、植物多样性、地上-地下生物量、根冠比及土壤理化性质进行了详细对比研究,以期揭示高寒草原和高寒草甸退化过程中的一些特征差异,为青藏高原退化草地恢复治理提供科学依据。
1 材料与方法
1.1 研究区概况
研究区为东北-西南样带,东北至祁连山脉,西南抵拉萨,西起青藏高原腹地长江源区,向东几乎达到青藏高原东缘,长1330km,宽820km(图1)。地理坐标29°42′N— 38°43′N, 90°37′E— 102°20′E,海拔约2700—4700m。属高寒大陆性季风气候,年平均气温-5.2—4.7 ℃,年平均降水66—560mm,一年内超过90%降水发生在暖季(4—10月)。草地下伏连续多年冻土,土壤发育微弱,平均厚度30—50cm。高寒草原属典型低温钙化土,土层薄,土壤贫瘠。高寒草甸属低温始成土,草毡层厚8—10cm,有机质层约20—30cm。由于海拔高,气温低,有机质大部分难以分解。高寒草原优势物种为禾本科针茅属(Stipa)和莎草科苔草属(Carex)等,伴生种有菊科蒿属(Artemisia)、豆科棘豆属(Oxytropis)等,高寒草甸优势物种为莎草科嵩草属(Kobresia),伴生种为菊科风毛菊属(Saussurea)、蓼科蓼属(Polygonum)等。
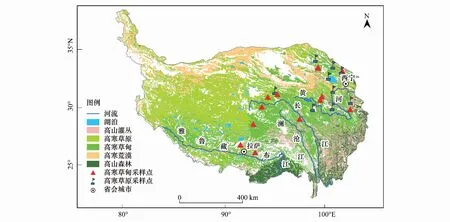
图1 青藏高原植被类型[26]及采样点分布概况Fig.1 Distribution of vegetation types and sampling locations on the Qinghai-Tibetan Plateau
1.2 研究方法
1.2.1退化等级划分
植被盖度对退化较敏感,因此,本研究将盖度作为划分退化等级的一种方法,研究结果仅代表以盖度划分的退化系列。参考Xue等[27]划分标准,依据植被盖度,将研究区高寒草甸划分为轻度退化(Light degradation,LD,植被盖度80%—90%)、中度退化(Moderate degradation,MD,植被盖度50%—80%)、重度退化(Severe degradation,SD,植被盖度15%—50%)和极度退化(Extreme degradation,ED,植被盖度<15%)4个梯度,与原生植被(None degradation,ND,优势种以莎草科嵩草属为主,植被盖度>90%)进行对照。高寒草原植被稀疏,退化梯度划分标准与高寒草甸略有不同,划分标准分别为ND(优势种以禾本科针茅属、莎草科苔草属为主,植被盖度>70%),LD(50%—70%),MD(30%—50%),SD(10%—30%),ED(<10%)。
1.2.2植被调查
2014年7月13日—9月20日,在青藏高原北麓河、青海湖、尕海等地典型高寒草地区开展植被调查。共设置22个样点,高寒草原设置9个样点,植被型以紫花针茅(Stipapurpurea)高寒草原、青藏苔草(Carexmoorcroftii)高寒草原为主。高寒草甸设置13个样点,植被型以高山嵩草(Kobresiapygmaea)高寒草甸、矮嵩草(Kobresiahumilis)高寒草甸为主。每个样点按退化梯度设置样地(至少包括四个退化梯度),每个样地随机设置3个1m×1m观测样方。调查植被盖度、多度、高度、频度、物种名称。共调查维管束植物29科153种。盖度采用数码相机(Canon,60D)拍照,再用CAN-EYE-V6313软件对照片图像进行分析,获取每个样方植被总盖度和每个物种分盖度。物种高度用钢卷尺测量,频度用物种出现的样方数/总样方数获得。
地上生物量采用刈割法收获,即将每个样方内植物地上部分全部刈割,分成莎草、禾草和杂类草,分别置入信封内,再将信封放入75℃烘箱内烘干至恒重并称重。地下生物量用直径为7cm土钻在每个样方随机钻取根系样品,分0—10cm、10—20cm、20—30cm和30—50cm 4个土层。根系样品带回实验室,过孔径0.28mm土壤筛,将根系冲洗干净。根据根的颜色、断面、柔韧性及是否附着毛根,分出活根和死根。将活根放入信封内,置入75℃烘箱内烘干至恒重并称重,作为地下生物量。本研究中地下生物量为0—50cm总地下生物量。地上和地下生物量均为当年净生产量,根冠比为当年9月份地下生物量与地上生物量之比。
1.2.3土壤理化性质测量
首先在每个样方用直径为7cm土钻随机钻取土壤样品,分0—10cm、10—20cm、20—30cm和30—50cm 4个土层。将新鲜土样置入铝盒内,立即称重后带回实验室,再将铝盒置入105℃烘箱烘干48h,至土壤样品恒重时称重,获取土壤体积含水量和土壤容重。
土壤有机碳采用重铬酸钾氧化法测定,pH值采用酸比重计测量(FE28K, Mettler Toledo, 上海),总氮通过德国哈瑙生产的VarioEL元素分析仪获取。由于高寒草甸优势种莎草根系50.61%分布在土壤表层0—10cm[28],因此本研究只分析了0—10cm土壤理化特征。
1.2.4数据分析
物种重要值(Species Importance Value,IV)=(相对高度+相对盖度+相对频度)/3。物种丰富度指数(Species Richness)用物种数(S)表示,Shannon-Wiener指数(H)计算公式如下:
式中,Ni为第i个物种的个体数,N=∑Ni,S为每个样方的物种数。
数据分析用IBM SPSS Statistics 23完成,相关性分析通过CANOCO 4.5和Pearson相关性分析完成,二维图用ArcGIS 10.1和Origin 8.5完成。
2 结果与分析
2.1 植被退化特征差异
2.1.1群落高度和植物多样性
表1显示,草地退化过程中,紫花针茅高寒草原群落高度显著降低(P<0.05),高山嵩草高寒草甸群落高度显著增加(P<0.05)。随着退化梯度加剧,高寒草原和高寒草甸物种丰富度指数和Shannon-Wiener指数均呈单峰式变化,峰值均出现在中度退化。高寒草甸退化过程中植物多样性更高。

表1 青藏高原退化高寒草地群落高度、盖度、植物多样性对比
ND:未退化None degradation;LD:轻度退化Light degradation;MD:中度退化Moderate degradation;SD:重度退化Severe degradation;ED:极度退化Extreme degradation; a、b、c、d、e表示不同退化梯度间差异水平(P<0.05)
2.1.2群落结构和物种组成
选取青海省祁连县龙洼沟冰草(Agropyroncristatum)高寒草原(38°12′11″N,100°09′31″E)和治多县北麓河高山嵩草高寒草甸(34°49′12″N,92°55′12″E[29])两个典型样地为例说明高寒草原和高寒草甸群落结构和物种组成变化(表2)。高寒草原在原生植被、轻度和中度退化建群种为冰草,重要值分别为0.40、0.30和0.23;重度退化梭罗草(Roegneriathoroldiana)(0.16)、紫花针茅(0.15)重要值明显高于其他物种;极度退化冰草消失,优势种为狼毒(Stellerachamaejasme)(0.21)和矮火绒草(Leontopodiumnanum)(0.11)和草地早熟禾(Poapratensis)(0.10)。高寒草甸原生植被建群种高山嵩草、矮嵩草、线叶嵩草(Kobresiacapillifolia)重要值分别为0.17、0.16和0.12;轻度退化高山嵩草(0.20)、矮嵩草(0.17)和青藏苔草(0.17)重要值最高;中度退化高山嵩草(0.15)、矮嵩草(0.15)和矮火绒草(0.13)为共同优势种;重度退化高原早熟禾(Poapratensissubsp.alpigena)(0.23)、丛生钉柱委陵菜(Potentillasaundersianavar.caespitosa)(0.14)成为建群种;极度退化矮火绒草(0.31)和块根紫菀(Asterasteroides)(0.21)重要值最高。因此,草地退化过程中,高寒草原优势种禾草的优势地位未改变,高寒草甸优势种嵩草逐渐被杂类草取代。

表2 青藏高原退化高寒草地物种组成变化差异
ND:未退化None degradation;LD:轻度退化Light degradation;MD:中度退化Moderate degradation;SD:重度退化Severe degradation;ED:极度退化Extreme degradation; IV: 重复值Important value
2.1.3地上-地下生物量及根冠比
图2表明,高寒草原退化过程中地上生物量显著下降了40.60%、68.62% 、75.68%和90.58%(P<0.05),地下生物量重度退化开始显著下降(P<0.05);根冠比退化过程中显著增加(P<0.05),由原生植被至极度退化增加了1.72倍;总生物量退化过程中分别下降了10.69%、30.32%、60.01%和82.26%。高寒草甸退化过程中地上生物量先保持稳定,极度退化显著降低了70.24%(P<0.05),地下生物量中度退化开始显著下降(P<0.05);根冠比退化过程中显著下降;总生物量退化过程中下降16.47%、54.22%、79.06%和95.03%。这说明高寒草原地上生物量较高寒草甸地上生物量对退化响应更敏感,高寒草甸地下生物量较高寒草原地下生物量对退化响应更敏感。高寒草甸总生物量较高寒草原总生物量对退化响应更敏感且下降幅度更大。
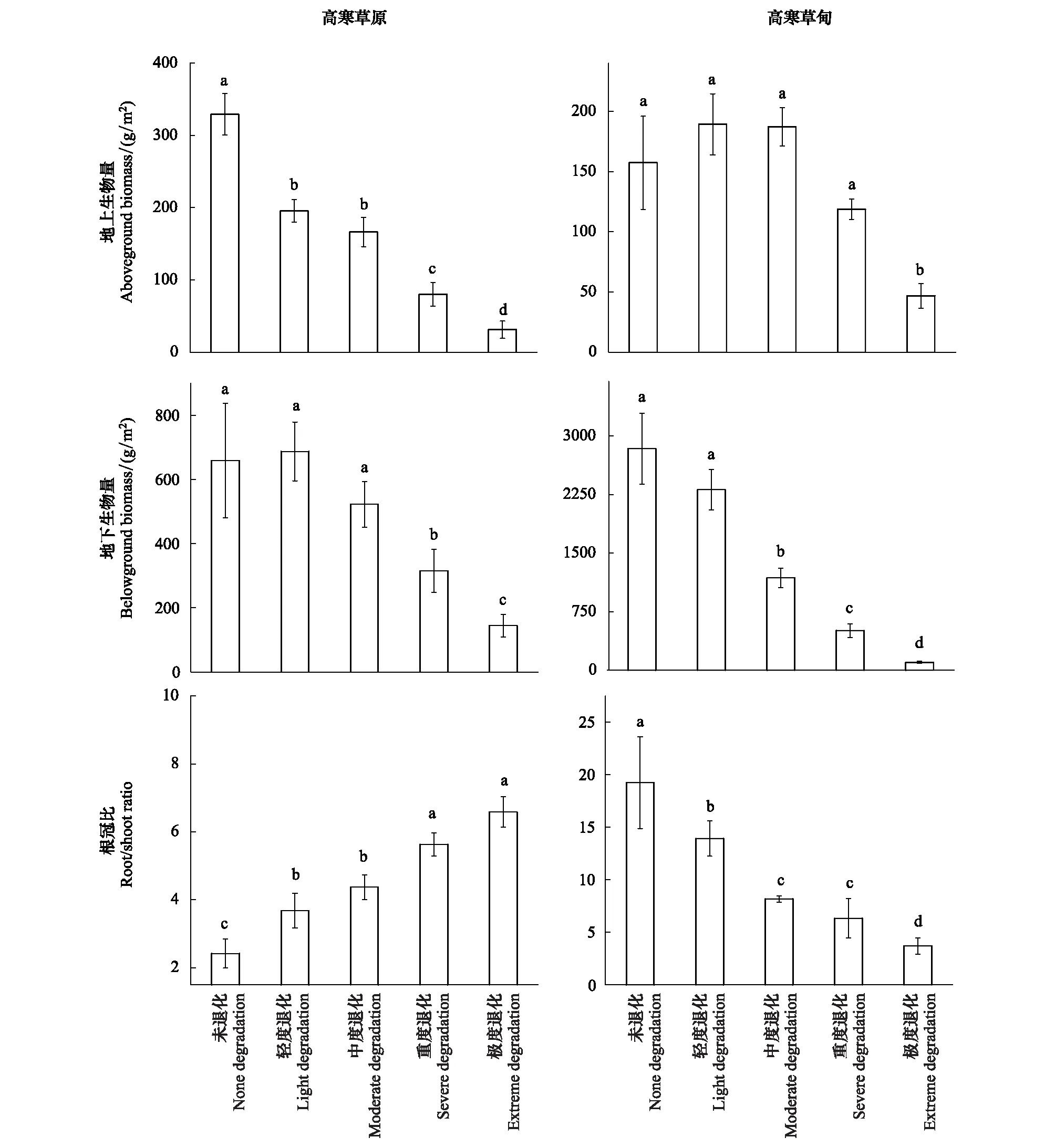
图2 高寒草原和高寒草甸不同退化梯度地上-地下生物量及根冠比对比Fig.2 Comparisons of above/belowground biomass and root/shoot ratio between alpine steppe and alpine meadow along degradation gradientsa、b、c、d表示不同退化梯度间差异水平(P<0.05)
2.1.4功能群地上生物量
由图3可知,随着退化梯度加剧,高寒草原莎草地上生物量变化不明显(P>0.05),禾草地上生物量显著降低(P<0.05),杂类草地上生物量先增加后减少。随着退化梯度加剧,高寒草甸莎草地上生物量显著降低,且彼此间差异显著(P<0.05),禾草和杂类草地上生物量先增加后减少。由原生植被至极度退化,高寒草原禾草地上生物量贡献率由88.12%减少至53.54%,杂类草地上生物量占比由0.08%增加至42.81%;由原生植被至极度退化,高寒草甸莎草地上生物量占比由69.15%减少至0.04%,杂类草地上生物量占比由12.56%增加至92.61%。
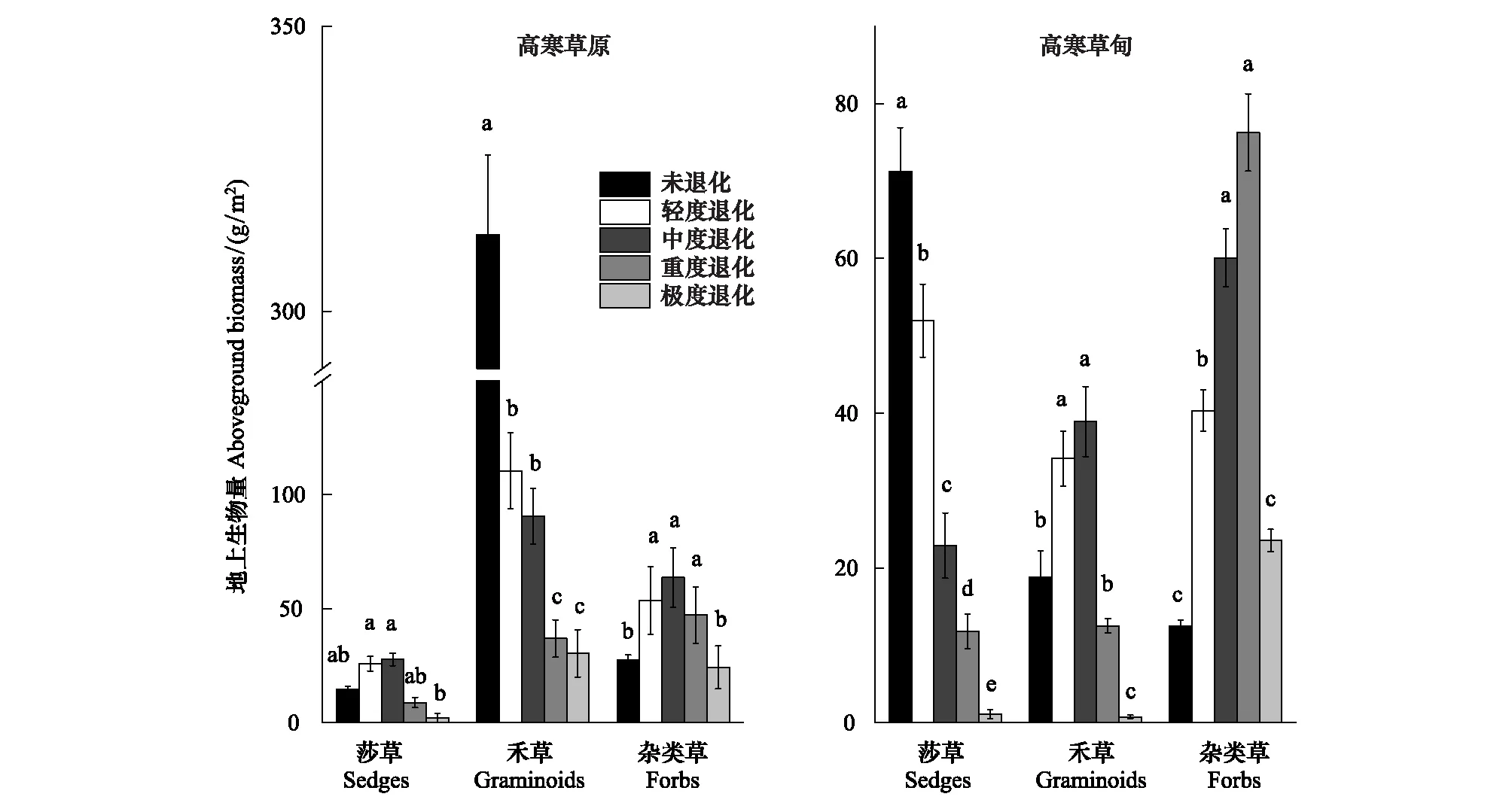
图3 高寒草原和高寒草甸不同退化梯度功能群地上生物量对比Fig.3 Comparisons of aboveground biomass of groups between alpine steppe and alpine meadow along degradation gradientsa、b、c、d、e表示不同退化梯度间差异水平(P<0.05)
2.1.5根系分配特征
由图4可知,随着高寒草原退化程度加剧,土壤0—10cm根系占比显著增加(P<0.05),由原生植被57.31%增加至极度退化75.63%;10—20cm、20—30cm和30—50cm根系占比均呈减小趋势。随着高寒草甸退化加剧,0—10cm根系占比显著减小(P<0.05),原生植被的根系占82.18%,至极度退化仅占44.76%。 10—20cm,20—30cm和30—50cm根系占比退化过程中均呈增加趋势。一般线性模型方差分析退化梯度和土壤深度交互效应结果显示:高寒草原达显著水平F(12,232)=1.732,P<0.05,高寒草甸达到极显著水平F(12,352)=4.259,P<0.001。以上结果表明:草地退化过程中,高寒草原根系向浅层迁移,高寒草甸根系向深层迁移。0—10cm高寒草甸根系随退化减少比例比高寒草原根系随退化增加比例高19.1%,说明高寒草甸根系比高寒草原根系退化过程中迁移速度更快。
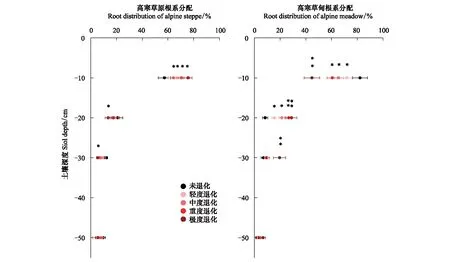
图4 高寒草原和高寒草甸不同土壤深度和退化梯度的根系分配对比Fig.4 Comparisons of roots distribution between alpine steppe and alpine meadow at both soil depthes and degradation gradients*和**分别表示0—10cm、10—20cm、20—30cm、30—50cm不同退化梯度间根系占比差异显著(P<0.05)和极显著(P<0.01)
2.2 土壤退化特征
由表3可知,随着退化梯度加剧,高寒草原土壤含水量变化不明显(P>0.05);土壤有机碳退化过程中分别减少27.08%、31.55%、70.18%、83.85%;总氮退化过程中分别流失27.16%、27.76%、35.82%、67.16%;土壤容重退化过程中分别增加了27.47%、31.87%、34.07%和39.56%。高寒草甸退化过程中土壤含水量显著降低(P<0.05);土壤有机碳退化过程中分别降低44.46%、37.88%、72.77%、91.43%;总氮退化过程中分别减少32.30%、22.43%、55.97%、72.63%;土壤容重退化过程中分别增加了25.68%、32.43%、58.11%和81.08%。由此可见,高寒草甸土壤含水量、土壤有机碳、总氮及土壤容重对退化响应均比高寒草原更强烈。
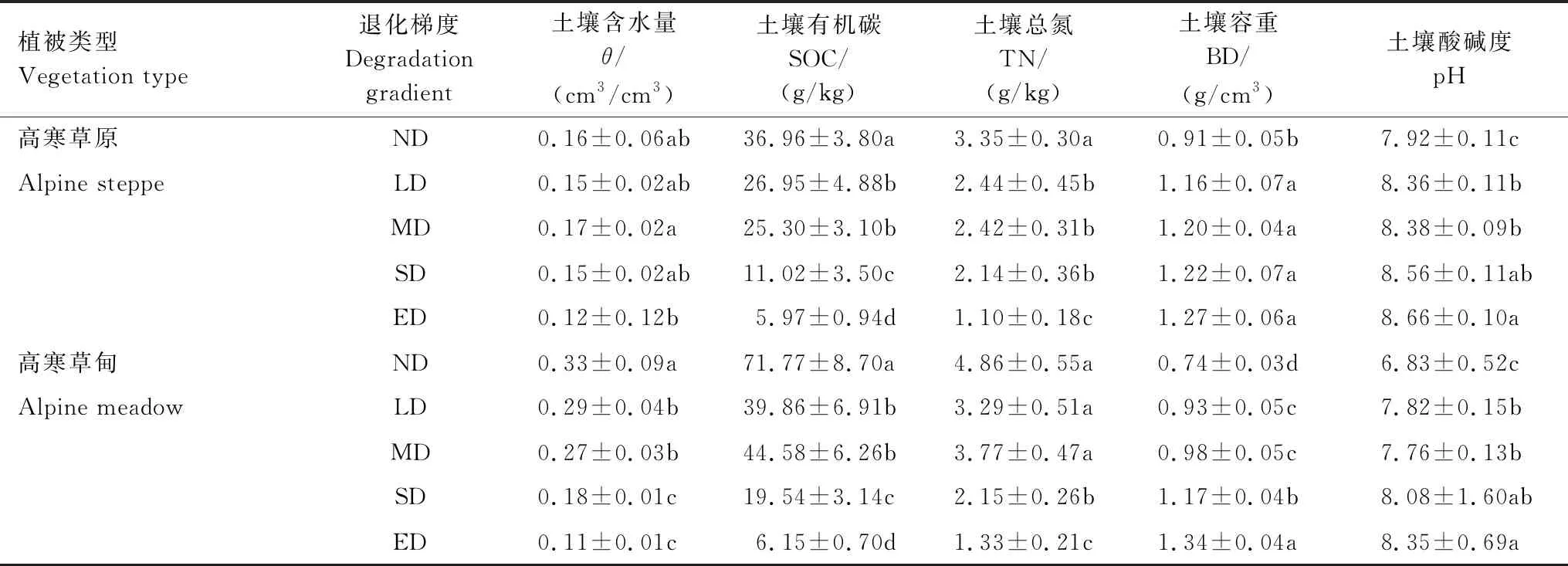
表3 高寒草原和高寒草甸不同退化梯度土壤理化性质对比
SOC:soil organic carbon;TN: total nitrogen (TN);BD: bulk density; a、b、c、d表示不同退化梯度间差异水平(P<0.05)
2.3 植被特征与土壤理化性质关系
图5是高寒草原和高寒草甸五个退化梯度总体地上-地下生物量、高度、盖度、物种丰富度、Shannon-Wiener指数及功能群莎草、禾草和杂类草地上生物量与土壤含水量、有机碳、总氮、土壤容重及pH值相关性对比。高寒草原地上生物量、禾草地上生物量及群落高度与土壤含水量、土壤有机碳、总氮显著正相关(P<0.05),与土壤容重极显著负相关(P<0.01),地下生物量与土壤容重显著负相关(P<0.05);高寒草甸地上生物量、禾草和杂类草地上生物量、群落高度、盖度、物种丰富度、Shannon-Wiener指数均与土壤有机碳和总氮正相关,地下生物量同土壤容重负相关,莎草地上生物量和土壤含水量正相关。Pearson相关性分析均达到极显著水平(P<0.01)。
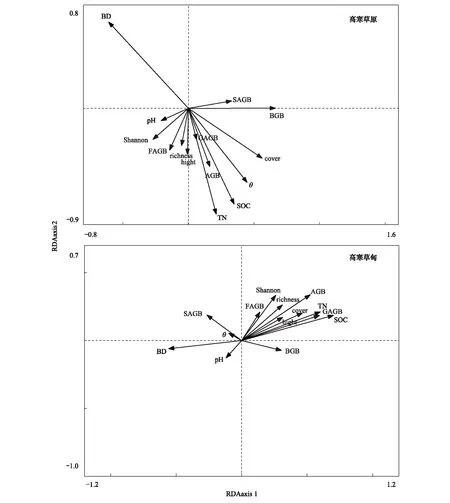
图5 高寒草原和高寒草甸植被因素和土壤特征RDA排序对比Fig.5 Comparisons of RDA ordination biplot of plant variables and soil conditions between alpine steppe and alpine meadow图中θ表示土壤体积含水量;SOC:Soil organic carbon,土壤有机碳;TN:Total nitrogen,总氮;BD:Bulk density,土壤容重;pH:土壤酸碱度;AGB:Aboveground biomass,地上生物量;SAGB:Sedge aboveground biomass,莎草地上生物量;FAGB:Forb aboveground biomass,杂类草地上生物量;GAGB:Graminoid aboveground biomass,禾草地上生物量;BGB:Belowground biomass,地下生物量;cover:盖度;hight:高度;richness:丰富度;Shannon:Shannon多样性
3 讨论
草地退化是一个复杂的过程。青藏高原草地退化受自然因素和人类不合理放牧共同影响[30]。就放牧而言,高寒草原和高寒草甸退化过程中植被群落结构、生产力的差异首先归因于放牧食性。牲畜对优势种禾草和莎草优先采食,造成禾草和莎草盖度减小,群落高度差减小,群落郁闭度降低,使竞争中处于弱势的物种,主要是低矮的豆科和毒杂草在光的竞争中受益。优势种禾草和莎草减少,杂类草增加[31],群落结构和多样性发生变化。功能群生物量占比随之改变。
家畜类型在很大程度也会导致高寒草原和高寒草甸草地退化和植被退化差异。高寒草原和高寒草甸放牧畜种以牦牛、藏绵羊为主。牦牛和藏绵羊对青藏高原下垫面植被的影响有所不同[32]。相关研究表明,高寒草原和高寒草甸同等放牧强度下,单纯放牧绵羊区域的植被生物量、草地植物高度和密度明显高于单纯放牧牦牛区域[33]。植物丰富度和Shannon多样性指数变化相反。单纯放牧绵羊的草地较为松软,透气透水性高;0—10cm、0—20cm和0—30cm土壤速效钾、速效氮、速效磷、全氮、全磷、有机质均高于单纯放牧牦牛的草地。单纯放牧牦牛草地退化更严重,其原因如下:一是食性差异,牦牛对低矮植株选择舔食,绵羊很少舔食。植物种子大多在地表富集,舔食使牦牛吃进消化道的种子量比绵羊大得多。二是践踏强度不同,牦牛和绵羊体重差别大,牦牛对地表植被践踏更严重,造成土壤容重增大,土壤透气透水性降低,影响植物根系发育。三是牦牛粪被作为燃料捡拾,很少留在草地上,绵羊粪大多留在草地上,因而放牧绵羊的草地土壤养分较高,而放牧牦牛的草地养分长期亏损。
功能群生理生态特征差异也会影响高寒草地退化过程中植被群落结构。禾草直立高大,比低矮的杂类草占据更高的生态位,对光资源竞争中处于优势。本研究中退化高寒草甸表层土壤容重增加,土壤含水量减少。表层土壤粗骨化,再加上啮齿动物洞穴增多,导致表层土壤水分和养分向深层入渗[19]。禾草和直根系杂类草向土壤深层伸展,吸收土壤深层水分和养分。研究发现禾草根系可延伸至地下85cm[34]。莎草根状茎盘结缠绕,不利于向地下伸展,只能利用土壤表层水分和养分。这一解释符合经典生态位理论[35],在环境胁迫条件下,处于不同生态位的物种利用不同的空间资源。
4 结论
(1)随着退化梯度加剧,高寒草原群落高度显著降低(P<0.05),高寒草甸群落高度显著增加(P<0.05)。草地退化过程中,高寒草原和高寒草甸物种丰富度和Shannon-Wiener指数均呈单峰式变化,高寒草甸植物多样性比高寒草原丰富。
(2)高寒草原退化过程中地上生物量显著下降(P<0.05),地下生物量重度退化开始显著下降(P<0.05),根冠比增加了1.72倍;高寒草甸退化过程中地上生物量先保持稳定,极度退化显著降低70.24%(P<0.05),地下生物量中度退化开始显著下降(P<0.05),根冠比显著下降(P<0.05)。
(3)就地上生物量而言,高寒草原对退化响应更敏感;就总生物量而言,高寒草甸对退化响应更敏感且下降幅度更大。
(4)高寒草原地上生物量和禾草地上生物量均与土壤有机碳、总氮显著正相关(P<0.05);高寒草甸莎草地上生物量与土壤含水量显著正相关(P<0.05),禾草和杂类草地上生物量与土壤有机碳和总氮显著正相关(P<0.05)。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
海外文摘·文学版(2021年3期)2021-05-19
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
畜牧兽医科学(2019年24期)2019-09-10
活力(2019年21期)2019-04-01
南方农业·下旬(2017年11期)2018-01-09
绿色科技(2017年15期)2017-09-01
中国饲料(2017年15期)2017-08-22
