
土壤微生物碳素利用效率研究进展
2020-03-27 09:24于贵瑞
生态学报 2020年3期
陈 智, 于贵瑞
1 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室,北京 100101 2 中国科学院大学资源与环境学院,北京 101408
通过有效增加碳(C)固持和合理减少C排放来共同控制和减缓全球变暖是全人类应对气候变化达成的共识[1]。为制定有效的增汇措施,人们迫切需要准确模拟和预测全球变暖与地球生态系统间的相互影响,尤其是陆地生态系统对全球变暖的反馈作用与机制[2- 3]。已有大量研究指出,全球变暖会促进土壤C释放,对全球变暖产生正反馈[2,4]。但是,越来越多的研究开始发现,微生物在调控陆地生态系统对全球变化的反馈中起着重要作用,并且可能改变预期的反馈效应[5- 7]。例如,长期的增温反而抑制微生物生物量和酶活性,从而减少对土壤有机碳的分解。微生物的生理代谢过程及其对外界环境变化的响应与适应成为影响陆地生态系统的反馈作用的关键[5]。
土壤微生物连接着土壤圈、生物圈、大气圈、水圈、岩石圈的物质循环,参与了土壤中几乎所有的物质转化过程[8]。土壤微生物的生长直接取决于微生物的碳素利用效率(Microbial carbon use efficiency, CUE),即微生物将吸收C转化为生物量C的效率。在大量的土壤C循环模型中,微生物CUE被设定为一个常数[9- 11]。然而,野外观测和室内培养实验并不支持这一假设。土壤微生物CUE可能随着外界环境和养分条件等的变化而显著变化。有研究指出,土壤微生物CUE随着土壤养分有效性的增加而增加[12- 13],而随着温度的升高而降低[5,14]。但是,对于这些潜在因素的影响并没有一致性的认识。例如,在北美大草原的研究表明,水分胁迫抑制微生物的生长和CUE[15]。然而,Herron等[16]发现土壤含水量的降低对土壤微生物CUE并无显著影响。这些不同的研究结论体现了人们对于土壤微生物CUE变异及其影响机制认识的不足,这制约了人们对陆地生态系统的反馈的准确模拟与预测[17]。
本研究对现有文献进行梳理和分析,概述土壤微生物CUE的定义和测定方法,重点综述和分析CUE的变异特征及其影响因素,并指出未来的研究重点,以期为当前地球系统模型的改进及对未来的气候变化趋势预测提供理论依据。
1 土壤微生物碳素利用率的定义
植被通过光合作用将大气中CO2转化为有机物质,形成生态系统的净初级生产力。绝大部分的植被生产力需要在分解者-土壤微生物的分解矿化作用下还原为无机养分,再被植被吸收利用,实现生态系统中物质和能量的生物地球化学循环。微生物的生理代谢过程实则是同化和异化代谢相耦合的过程。微生物将植被光合产物一部分同化为微生物生物量,另一部分以呼吸代谢形式释放到大气中。此过程中,微生物将植被生产力转化为微生物生物量的效率称为微生物的碳素利用效率[13,18],也称为微生物生长效率或底物利用效率[19- 21]。土壤微生物CUE是土壤C循环过程中的关键生态学参数,它直接影响着生态系统的C素滞留时间、周转速率以及土壤的C存储能力[7,22- 23]。
在生态学研究中,微生物CUE通常表示为微生物的生长(μ)与吸收(U)的比值[13,18],即CUE=μ/U。微生物从外界吸收C主要用于微生物的生长(μ)、呼吸代谢(R)、胞外酶和代谢产物的分泌(EX)以及微生物死亡(BD)4个方面(图1)。根据质量守恒原理,微生物U表示为:
U=μ+R+EX+BD
其中,土壤微生物呼吸(R)包括微生物用于生长(RG)、维持(RM)、胞外酶生产(RE)以及溢流过程(RO)中产生的呼吸[13],即:
R=RG+RM+RE+RO
根据CUE定义和质量守恒方程,微生物CUE表示为:
在自然生态系统中,EX和BD通常难以测定,并且相对于生长和呼吸量,EX和BD量很微小而常被认为可以忽略[13]。因此,CUE通常被认为是μ与R两个过程的平衡关系,即,
这一定义被广泛用于当前的微生物代谢和土壤碳循环模型中[13,18]。
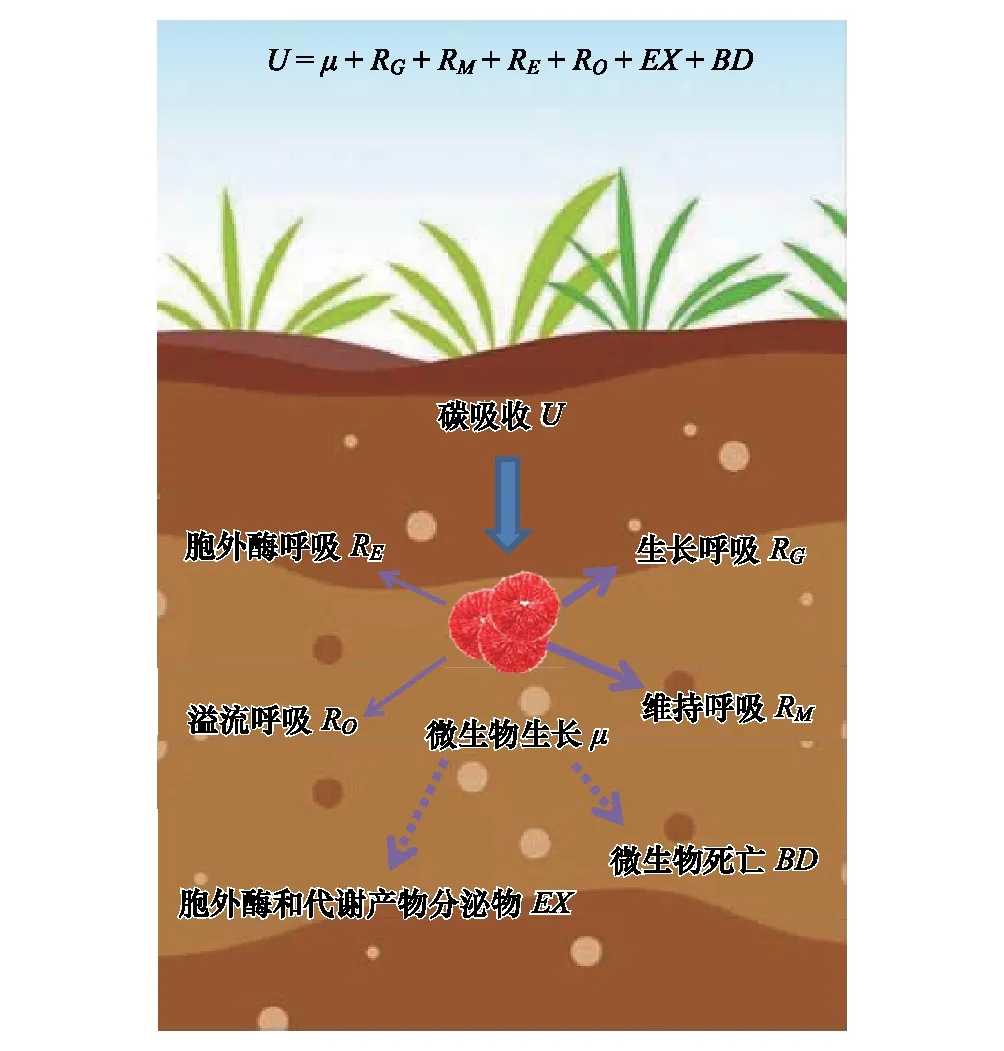
图1 微生物生长代谢组分和质量平衡方程 Fig.1 Microbial metabolic components and equilibrium equation根据土壤微生物CUE的定义和Manzoni等[13]提出的土壤微生物代谢质量平衡方程绘制; U, 微生物碳吸收microbial carbon uptake; μ, 微生物生长microbial growth; RG, 微生物生长呼吸respiration for microbial growth; RM, 微生物维持呼吸respiration for microbial maintenance; RE, 胞外酶呼吸respiration for extracellular enzyme production; RO, 溢流呼吸overflow respiration; EX, 胞外酶和代谢产物分泌物excretion of extracellular enzymes and metabolites; BD, 微生物死亡microbial mortality
2 土壤微生物碳素利用率的测定方法
土壤微生物CUE多采用室内培养结合质量守恒法和标记物示踪法进行测定。质量守恒法是直接测定物质质量或浓度的变化,根据物质守恒原理计算CUE。标记物示踪法则是通过标记底物,有效追踪底物的利用路径,计算底物被用于生长和呼吸的比例进行测定,是目前较为广泛采用的方法。根据研究途径、研究对象(微生物或底物)的不同,现有分析方法可大致分为基于微生物生长速率测定法、基于微生物生物量测定法、基于底物吸收速率测定法和基于底物浓度变化测定法。这些方法各具优缺点和适用范围(表1)。
2.1 生长速率法
生长速率法是最早的测定方法之一,它通过直接测定微生物的生物合成速率和呼吸速率计算微生物CUE。以3H-thymidine、3H-leucine等作为标记底物,通过短时间的培养,在三氯乙酸的消解作用下分离出合成的核酸或蛋白质,结合放射性同位素分析,测定微生物将3H-thymidine合成为DNA或将3H-leucine合成为蛋白质的速率[24-26]。该方法直接准确,主要用于水生生态系统。在陆地土壤中,采用以18O标记的重水H218O进行培养,测定微生物从水中吸收18O合成DNA的量,结合微生物DNA和生物量C的转化系数计算土壤微生物CUE。该方法可以直接测定微生物的生物合成速率,但主要用于短时期培养[27-28]。
2.2 生物量法
在陆地生态系统,例如土壤中,通常基于微生物生物量的变化计算CUE。添加标记的活性有机碳作为底物进行室内培养,结合氯仿熏蒸法测定微生物生物量的变化。该方法简单,可操作性强,是一种常用的标准方法。但是,由于在较短时期内难以捕捉微生物群落的生长及维持呼吸,该方法在短期内所测定的活性标记物转化为生物量的速率并不完全等同于微生物的生长速率,而更多的是群落的吸收速率,因此存在高估CUE的现象[29- 31]。
2.3 底物吸收速率法
底物吸收速率法是从观测底物变化的角度测定CUE。将3H-thymidine、3H-leucine等作为标记底物,进行短期室内培养测定其被吸收利用的速率。该方法相对于微生物生物量测定法,考虑了微生物产物的损失。但是,底物的吸收速率通常难以准确测定,只适用于短期的室内培养分析。
2.4 底物浓度变化法
与底物吸收速率法相似,底物浓度变化法也是从底物变化的角度测定CUE。该方法通过添加高浓度的底物(葡萄糖, 醋酸等),经过一段时间的室内培养,阶段性的测定底物浓度的变化,结合同期的呼吸量计算CUE。该方法通常不需要对底物进行同位素标记,但需要提供高浓度的底物介质。由于该方法需要一定长度的培养时间,因此考虑了微生物产物的代谢和分泌过程。

表1 微生物碳素利用率测定和计算方法比较
CUE, 碳素利用率carbon use efficiency;μ, 微生物生长速率microbial growth rate;R, 微生物总呼吸速率microbial total respiration rate;U, 底物吸收速率substrate uptake rate;ΔCB, 微生物生物量变化量microbial biomass change;ΔCS, 底物浓度变化量substrate concentration change;RCUM, 累积呼吸速率cumulative respiration rate;AE, 元素(E)的吸收效率assimilation efficiency of element E;BC∶E, 微生物生物量的C∶E比the C∶E ratio of microbial biomass; TERC:E, 微生物最优生长所需C∶E比阈值threshold of C:E ratio required for microbial optimum growth
2.5 化学计量比法
基于化学计量比法是一种不需要直接测定CUE而根据元素化学计量限制理论计算CUE的方法,被广泛用于土壤和凋落物分解模型中[18,32]。根据化学计量平衡理论,当外源养分中的必需元素(如N,P)与C素的比例低于微生物最优生长所需N/P:C元素比阈值时,微生物的生长将会受到该元素有效性的制约。CUE与养分的元素计量比存在一阶函数关系,表示为CUE=AE×BC:E/TERC:E。其中,AE表示必需元素(E)的吸收效率,BC∶E表示微生物生物量的C∶E 比,TERC:E表示微生物最优生长所需C∶E比阈值[33- 35]。该方法具有理论基础,不需要实验测定,可参数化和模拟,但是是一种经验关系,主要用于土壤碳循环过程模型的模拟中。
3 土壤微生物碳素利用率的变异性
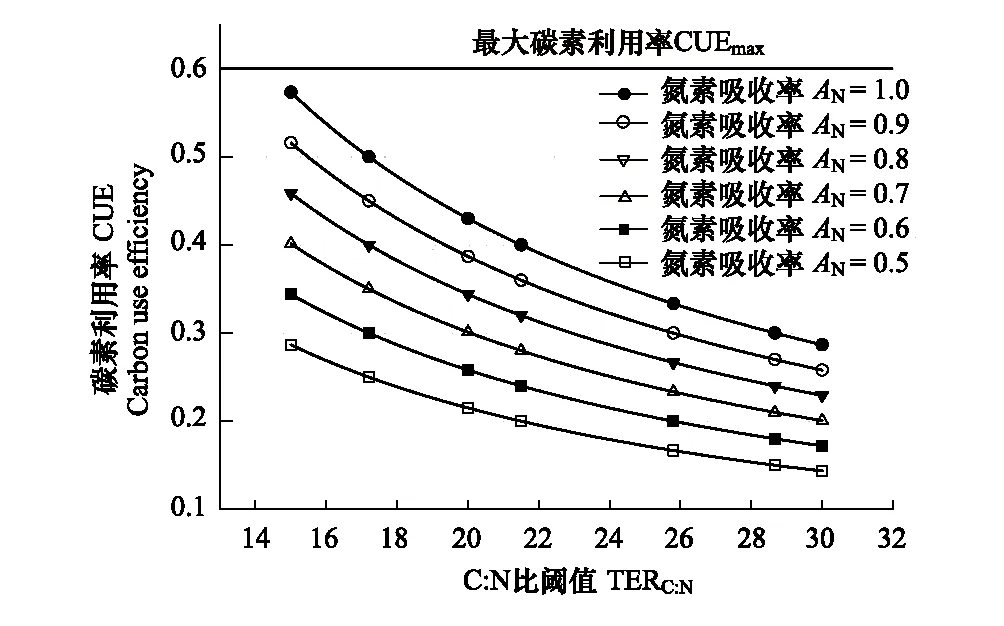
图2 微生物碳素利用率CUE与C:N比阈值TERC:N和氮素吸收率AN的关系Fig.2 Relationships between microbial carbon use efficiency and threshold of C:N and nitrogen uptake efficiency根据生物化学元素计量平衡理论绘制,参见Sinsabaugh等[29]
在自然生态系统中,土壤微生物CUE并非常数。在理想状态下,微生物只进行有机物同化并完全同化底物,那么碳素利用率为1。但是微生物的生长始终伴随着呼吸代谢消耗,在热力学限制下,微生物的最大CUE(CUEmax)小于1[38- 40]。微生物的实际生长受自身化学计量平衡的调控,表现为CUE随着必需元素(E)的吸收效率、微生物的C∶E比和最优生长C∶E比阈值的变化而变化(图2)。当氮素吸收效率AN=1,土壤微生物CUE在C:N比阈值TERC:N为15时可接近于最大CUE(CUEmax≈0.6);随着TERC:N增加到30,土壤微生物CUE则降到CUEmax的1/2(CUEmax/2≈0.3)(图2)。在实际土壤环境中,AN通常小于1,因此微生物实际CUE难以达到CUEmax。从ATP生成、电子传递和能量转化3个不同途径测定的结果表明,受到热力学限制,微生物的实际最大CUE约为0.6[38- 39]。Sinsabaugh等[29]的研究指出,在考虑了微生物维持代谢消耗之后,热力学最大CUE约为0.55。
基于实测数据的整合结果表明,土壤微生物CUE可在0.2—0.8的范围内变化。Sinsabaugh等[41]通过建立的生态酶活性、生物量组成和养分浓度的化学计量模型,结合全球生态酶活性数据得出土壤微生物CUE平均约为0.29,接近于CUEmax/2。基于C:N元素化学计量平衡方程估算的结果也得出,土壤微生物CUE平均约为0.27±0.11,与基于C:P元素计量方程的估算结果相近(0.25±0.12)[18]。但是,实验结果也显示,不同土壤层的微生物CUE存在差异。例如,矿物层微生物CUE(0.284±0.005)高于有机层微生物CUE(0.205±0.008)[42]。不同微生物种群也表现出不同的CUE。Sinsabaugh等[43]基于实验观测数据的整合分析得出,细菌的CUE约为0.336±0.213,高于真菌的CUE(0.326±0.196)。土壤微生物CUE还因植被类型的不同而不同。Takriti等[44]对西伯利亚植被样带的调查研究表明,土壤微生物CUE从草甸草原向泰加林、苔原逐渐降低。
在当前大量的生物地球化学循环模型中,土壤微生物CUE则通常被设定成一个参数,集中在0.25—0.6的范围(表2)。在不同的生物地球化学模型中,因考虑不同底物组成等影响,CUE参数值存在差异。例如CENTURY模型,地下和地上有机质的分解采用不同土壤微生物CUE,分别为0.45和0.55[9]。而Daisy、NCSOIL、ICBM等模型则考虑碳库活性的影响,对于活性碳库,CUE为0.6,而对惰性碳库的CUE小于0.6[10- 11,45](表2)。但是,Manzoni等[32]指出,实测的土壤微生物CUE结果往往低于模型的预设值,当前的模型在一定上低估了真实的异氧呼吸通量。
4 土壤微生物碳素利用率的影响因素
土壤微生物CUE受到来自热力学、生理学和生态学的调控,包括温度、水分等环境因子、土壤质地、底物和养分有效性、微生物群落组成等多种因素的影响。
4.1 热力学限制
微生物的碳吸收过程是同化和异化的耦合过程,微生物在转化植被光合产物为自身生物量的同时,时刻伴随着自身呼吸的代谢消耗。在热力学限制下,微生物的理论最大CUE约为0.8[40]。这表明,不超过80%的吸收有机C可被微生物用于自身生长,而至少20%的C需要用于维持呼吸的能量消耗。然而,实际的土壤微生物CUE难以达到理论的最大值,因为在生理和生态环境因子的制约下,微生物还需投入更多的能量到呼吸和养分吸收等过程。
4.2 生态环境因子限制
环境因子,例如温度、湿度、降水、土壤水分,会影响微生物代谢,改变μ和R间的平衡关系,从而影响CUE[5,13]。大量研究表明,土壤微生物CUE对增温呈现负反馈,CUE随着温度的升高而降低[5,13- 14,53- 54]。这是因为在温度的调控下,微生物的生长和代谢速率均随着温度的升高以Arrhenius方程的形式增加[55- 57]。然而,微生物呼吸代谢的温度敏感性高于生长响应[55],微生物呼吸的增加速度快于微生物生长,使得CUE降低[5,54,58]。Steinweg等[59]的研究指出,温度每升高1℃,土壤微生物CUE约减少0.009。高温胁迫下微生物CUE的负反馈效应更为显著[53]。模拟结果表明,30年的连续增温降低了吸收C被用于微生物生长的比例,CUE从0.31减少到0.23[5]。但是,也有研究指出,温度的升高,土壤微生物CUE并未发生显著变化[37,60]。土壤微生物CUE对温度的响应方式受底物的组成和代谢阶段的影响。Öquist等[61]的研究指出,单分子结构底物供给下的土壤微生物CUE随着温度的升高而降低,而多聚体结构底物下的土壤微生物CUE则不随温度的升高而改变。一些研究还指出,长期的增温将会使微生物产生适应性。长期增温下微生物将降低其基础呼吸速率[62]。在哈佛森林开展的连续增温实验表明,18年的5℃增温使得土壤微生物CUE随温度升高而降低的程度要低于连续2年的增温效应[6]。由于微生物的热适应性与微生物群落组成改变、养分有效性降低、微生物代谢途径的转变等有关,同时也受底物和观测方法的影响,微生物对温度的响应及其机理仍存在很多不确定性。
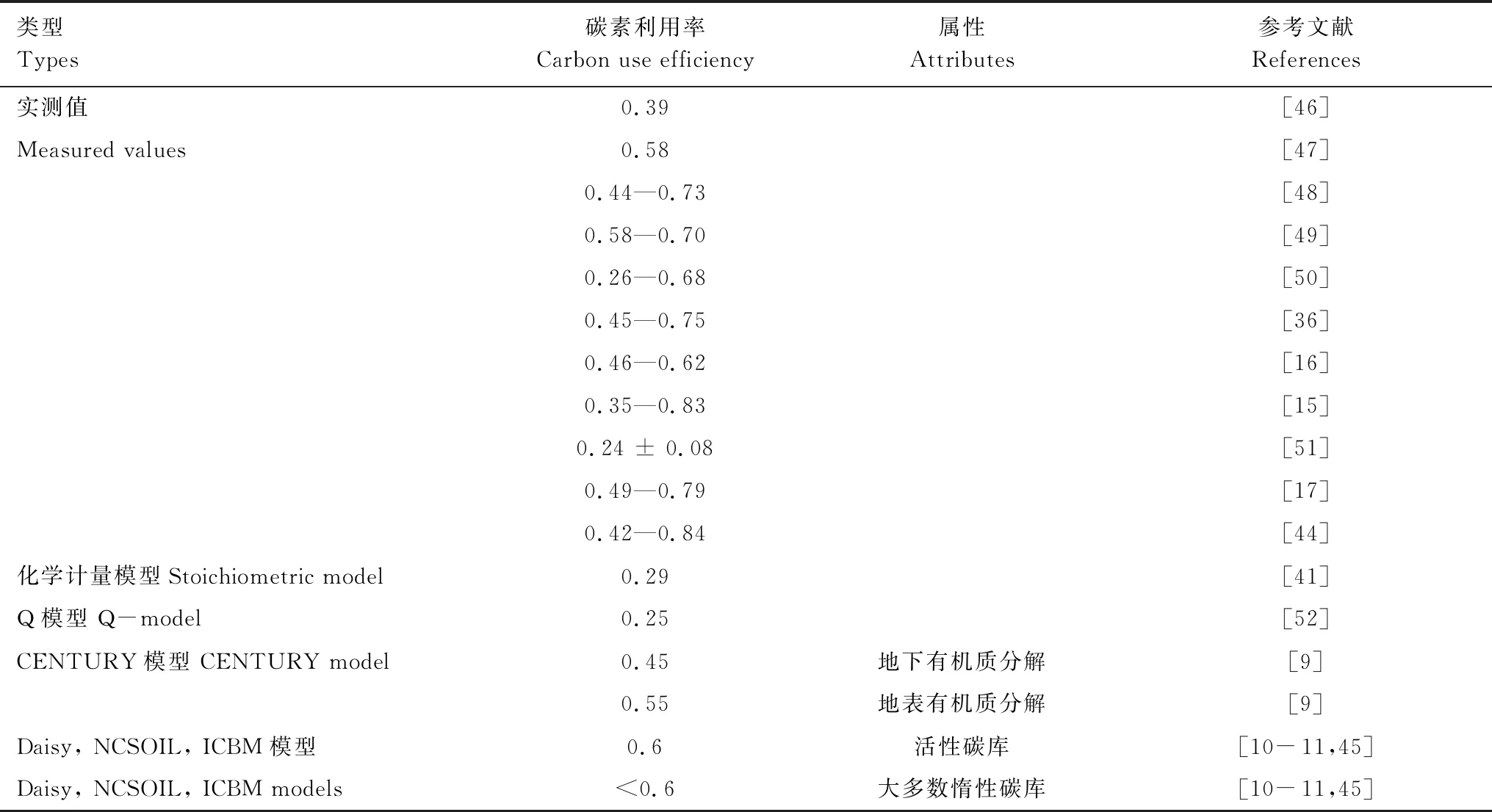
表2 微生物碳素利用率的变异性
土壤湿度和水分有效性是影响微生物生长和呼吸过程,进而影响CUE的另一重要环境因子[15]。与温度的影响相似,土壤水分有效性对CUE的影响复杂而多变,受到土壤水分胁迫强度、时长以及土壤类型的影响。研究表明,在短期的水分胁迫下,土壤水势的降低会激发微生物对水分胁迫的响应,促进微生物通过增加渗透压调控或采用短期休眠来减少干旱的影响,使得土壤微生物CUE增加[16,63]。然而,长时间的水分胁迫会降低土壤底物的溶解性和吸收性,从而抑制微生物的生长[13,64]。同时,长期的水分胁迫还会增加微生物的代谢消耗,进而降低CUE[15]。研究表明,厌氧环境下的土壤微生物CUE低于有氧环境[36]。在厌氧环境下,土壤微生物的代谢产物将由CO2转变为不能被完全氧化的CH4形式释放,CUE降低[65]。
4.3 底物质量和有效性
微生物吸收利用的底物来自于土壤有机碳和植被凋落物,不同的底物质量将会显著影响土壤微生物CUE[17]。这种影响源自底物的不同物质组成、分解过程、还原性程度以及有效性等。分解结构复杂的底物(如木质素和酚类等),需要经历更多的酶促分解反应过程,增加了微生物的呼吸代谢,CUE会降低[61,66]。对难分解性底物的降解还将增加胞外酶代谢,也使得CUE降低[67]。Takriti等[44]的研究指出,土壤微生物CUE在地理分布上的分异主要受到各样点土壤底物质量差异的影响。
底物的C还原性程度(γS)是影响土壤微生物CUE的另一个重要因素。C还原性程度是指每摩尔C的化学能,通常用每摩尔C的电子当量表示。被微生物利用的主要底物的γS通常介于3—5的范围内(如有机酸、葡萄糖、碳水化合物和脂类),与土壤微生物的C还原性程度相当(γB≈4.2)[39]。当底物的γs低于微生物γB时,由于单位底物所具有的能量不能满足一个单位生物量生产的能量需求,土壤微生物CUE较低[39- 40]。对16种不同还原性程度的底物CUE的分析结果表明,糖类CUE(0.667)> 氨基糖CUE(0.601)> 氨基酸CUE(0.551)> 有机酸CUE(0.498)[17]。
此外,土壤微生物CUE还会受到底物养分有效性的影响。当养分有效性和组成发生改变时,微生物会通过改变其化学酶调控的碳同化途径来响应养分的变化[68- 70]。大量研究表明,土壤微生物CUE会随着养分有效性的增加而增加[12- 13]。微生物碳吸收速率与养分浓度间存在饱和函数关系。在资源限制下,微生物将维持能满足吸收系统资源消耗下的最优碳吸收速率[41,70]。当养分有效性增加,养分浓度超过微生物的平衡浓度时,将会促进碳吸收,从而提高CUE。相反,养分的限制则会降低CUE。养分的限制会改变微生物自身的分解和合成耦合代谢过程,增加胞外酶和多糖等代谢产物,使CUE降低[71- 72]。大量的N添加实验也证明这一点,N素等养分的添加,会刺激微生物的活性或降低微生物呼吸代谢,微生物表现出高的CUE,尤其在N素限制的环境下[20,51,73- 74]。N添加主要促进了微生物对活性C的利用效率[74]。有研究也指出,长期的N添加后活性的C库被逐渐分解,微生物群落会最终转变为受C素等能量限制,从而土壤微生物CUE降低。由于大部分养分添加实验仍处在有限的时间范围,揭示微生物如何响应与适应养分有效性的变化还需要更深入的长期观测研究。
为满足对多种养分元素的需求,土壤微生物还会进行细胞内的资源调节和再分配,进一步影响微生物的生长和CUE[75- 78]。当受某种养分限制时,微生物会投入更多的能量来获取缺乏的营养元素,从而抑制了微生物生长和CUE[12- 13]。例如,受P素限制时,微生物会增加用于P元素资源获取的投入,使得CUE降低[29]。实验表明,养分的不足将抑制微生物的生长,使得CUE降低[61]。这种养分有效性的限制作用通常是多种养分有效性共同限制的结果[29]。
4.4 生态化学计量学限制
底物的元素化学计量比的变化显著影响微生物的生长和CUE[13,79]。分解者-微生物具有稳定的元素化学计量比,且不同于外界环境和分解底物的化学计量组成。研究表明,土壤微生物的C/N约为7—8.6[79- 81],高于浮游生物(6.6)和水生生物(8.3),而低于土壤有机碳(12.3)和植被凋落物(18.2—75)[79- 83]。土壤微生物的C/P约为23—60[79- 80],显著低于浮游生物(106)、水生生态系统(166)、土壤有机碳(72)和植被凋落物(100—837)[79- 83]。
根据化学计量比限制理论,土壤微生物会优先吸收利用元素组成最适宜其生长所需的物质,以有利于维持自身的化学计量比平衡。当底物的C含量与必需元素E含量的比值(C∶E)低于微生物最优生长所需元素比例的阈值(TERC∶E)时,有利于元素的矿化;而当C∶E高于TERC∶E时,有利于元素的固持。因此,分解N、P元素含量高的底物,土壤微生物表现为C限制,CUE较高。当分解底物N、P元素含量低时,土壤微生物可能有两种反应:一是抑制对底物C和养分的吸收;二是将多余的C以溢流呼吸或代谢分泌物的形式释放[35]。第一种反应将不影响微生物的CUE,然而第二种反应由于增加了呼吸代谢和分泌物释放,将会降低土壤微生物CUE。
大量研究指出,土壤微生物CUE与土壤有机质C∶N、C∶P比等呈负相关关系[33,42,44]。Lee 等[73]的研究发现,农田和落叶林中的土壤微生物CUE均随着土壤可溶性C∶N的升高而降低。Takriti等[44]对不同海拔上土壤微生物CUE的变异研究也指出,在有机层和矿物层表层土壤中,随着纬度的升高,土壤C∶N升高,土壤微生物CUE呈下降趋势。
4.5 微生物群落组成
土壤微生物CUE还受到微生物群落结构和组成的影响,因为不同的微生物种群具有不同的有机质分解和吸收速率[84- 85]。有研究表明,采用‘机会’生长策略的快速生长型微生物通常更适应高养分含量的环境,相对于缓慢生长的微生物具有较低的CUE[86- 88]。Adu等[47]的研究也指出,以真菌为主的微生物群落的CUE往往高于细菌为主的微生物群落。真菌的C∶N∶P变异范围通常宽于细菌,并且真菌的C∶N高于细菌具有更高的C素需求,表现出高的CUE[87]。但是,也有研究指出,土壤群落的CUE并无显著差异[19- 20]。Thiet等[20]在农田生态系统的研究中发现,高真菌/细菌比的群落与低真菌/细菌比的群落的CUE并无显著差异,分别为0.59±0.02和0.61±0.01。有研究也指出,底物C∶E的增加可以增加真菌群落的CUE,但却降低细菌群落的CUE[87]。微生物群落的种间竞争会降低微生物的CUE[89]。由于微生物的群落组成极易受到外界环境变化以及人为活动的影响,也随着底物质量和组成的变化而变化,量化不同群落组成的CUE的差异仍需进一步深入研究[19,87]。
5 问题与展望
5.1 研究尺度辨析
微生物CUE通指微生物吸收利用C并将其转化为自身产物的效率,但它却具有特定的时间和空间属性[90]。Geyer等[91]将微生物CUE分为3个层次:种群CUE(CUEP)、群落CUE(CUEC)和生态系统CUE(CUEE)。这3个层次的CUE表征了从种群到生态系统不同尺度上微生物的生物合成效率,包含了不同的微生物生理和化学过程[91]。前期大量的研究更多关注的是种群和群落尺度的微生物CUE,近期有越来越多的研究着眼于探讨生态系统尺度CUE的变化及其对环境的响应[92- 93]。不同研究尺度、观测阶段和测定条件下获取的CUE结果比较将给解析结果带来更大难度和不确定性[90]。在未来的CUE研究中,亟需更清晰地辨明微生物CUE的特定研究尺度和过程以准确地比较和应用。
5.2 代谢组分分析
当前土壤CUE的计算方法不统一并且存在高估现象。微生物的呼吸是多个组分的呼吸的总和,包括了用于生长的生长呼吸、有机体运转的维持呼吸、分泌胞外酶的呼吸以及溢流的呼吸。当前微生物CUE研究大多只考虑了微生物的生长呼吸或生长和维持呼吸,极少考虑胞外酶和溢流呼吸组分[13]。这使得CUE被高估,同时也使得CUE的计算结果存在很大差异。在今后的研究中,还需充分考虑胞外酶和溢流呼吸的损失。例如,在室内培养的测定过程中,选取适宜的培养时长观测包含微生物产物代谢和分泌的完整过程。此外,采用同位素标记与代谢组学相结合方法测算酶的产量和转化。
5.3 多要素交互影响研究
由于复杂的土壤-植物-微生物交互作用,环境变化对土壤微生物CUE的影响以及土壤微生物CUE对全球变化的响应与反馈仍存在很大的不确定性。例如,土壤微生物CUE是否随着温度升高而持续降低?是否存在热适应性?长期的外源养分输入下土壤微生物CUE将如何变化?等。由于土壤微生物CUE对环境变化的响应与适应因不同的底物质量、有效性、元素化学计量组成、微生物活性、群落组成而复杂多变[58],在未来的过程机理研究中,还需要综合探讨环境和生物多要素交互影响下,微生物的生理响应过程与对CUE的影响和调节机制。
5.4 土壤碳循环模型优化
土壤微生物CUE是土壤C循环模型中的重要参数。现有许多模型假定CUE是一个常数或很少考虑,使得无法准确模拟土壤微生物CUE对外界环境条件变化的动态响应过程及其对土壤C循环的影响[7,94]。有的模型则将微生物CUE表达为温度、养分有效性或养分元素比的一阶函数关系,而忽略了微生物的生理过程,如微生物代谢途径、群落组成、种间关系等的变化。此外,大量生物地球化学模型中,微生物呼吸过程通常被认为主要与微生物C平衡相关,而与N平衡无关,更是缺乏考虑P元素的动态变化的影响[29,35]。在未来的土壤C周转模型优化中,还需综合考虑元素计量平衡制约、微生物对外界环境条件变化的动态响应过程,以及微生物代谢途径、群落组成、种间关系、微生物周转等生理过程变化。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
今日农业(2020年19期)2020-12-14
科学(2020年2期)2020-08-24
河南科学(2020年3期)2020-06-02
山东化工(2019年18期)2019-10-16
科技资讯(2018年16期)2018-10-26
科技资讯(2017年12期)2017-06-09
草业科学(2015年9期)2015-12-11
中国科技信息(2015年2期)2015-11-16
