
渤海当年生小黄鱼矢耳石微结构特征*
2020-03-25 04:06张廷廷王玉堃金显仕
渔业科学进展 2020年2期
张廷廷 王玉堃 袁 威 金显仕,2 陈 超 孙 耀,2
渤海当年生小黄鱼矢耳石微结构特征*
张廷廷1王玉堃1袁 威1金显仕1,2陈 超1孙 耀1,2①
(1. 中国水产科学研究院黄海水产研究所 农业农村部海洋渔业可持续发展重点实验室 青岛 266071;2. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071)
以渤海海域采集的小黄鱼()幼鱼为研究对象,分析其矢耳石的微结构特征。结果显示,小黄鱼矢耳石中心核呈圆形,平均直径为(24.35±0.72)μm,核内耳石原基为深黑色的圆形结构,平均直径为(12.07±0.58) μm,第1日轮距中心核的距离为(14.11±1.08) μm。初次摄食标记轮出现在第4条日轮处,标记轮暗带颜色加深、清晰可辨,该标记轮距中心核的距离为(20.67±2.28) μm。小黄鱼矢耳石前20条日轮,轮纹由圆环状逐渐变为椭圆,沿最长轴方向的轮纹宽度变化范围为6.51~14.37 μm,且随着日龄的增加而逐渐变宽;20~40条日轮间轮纹宽度呈显著升高,40~85日龄轮纹宽度变化相对平稳,第56~60条轮纹处出现轮纹,平均宽度最大值为41.59 μm。在小黄鱼矢耳石日轮沉积过程中,伴随着亚日轮结构的形成,亚日轮清晰度差,轮纹宽度较窄且波动较大,在显微镜下对视野轻微调焦时会暂时性“消失”。本研究对小黄鱼的矢耳石特征结构进行了分析,以期为野生小黄鱼及其他鱼种的日轮判断提供参考。
小黄鱼;耳石;日轮;微结构
小黄鱼()隶属于鲈形目(Perciformes)、石首鱼科(Sciaenidae),广泛分布于渤海、黄海、东海以及朝鲜半岛西岸近海海域(主要集中在28°00¢N以北、125°30¢E以西),属暖温性、近底层洄游性鱼类,是我国重要的海洋经济鱼类之一(熊瑛等, 2014)。近几十年来,受捕捞业日趋发达、环境恶化等影响,小黄鱼资源量明显降低,其生活史、选择型也随之发生一系列变化,如个体小型化、性成熟提早及渔获物以幼鱼为主等(金显仕, 1996; 张国政等, 2010)。
耳石(Otolith)是位于硬骨鱼类内耳膜迷路内的一种钙化组织,是鱼体控制身体平衡的重要器官,同时记录了鱼类生命周期内大量的生物与生态学信息,可以作为鱼龄鉴定、产卵地调查及早期生长特征研究的材料(Arkhipkin, 2000)。陆化杰等(2012)根据阿根廷滑柔鱼()耳石微结构推测其日龄,并分析了其个体早期生长发育特征;Hislop等(2001)研究了鮟鱇鱼)耳石日轮特征以及产卵场的分布;Toshihiko等(2007)通过对大马哈鱼()耳石日轮及特殊标记轮的研究反演其早期生活史特征;黎雨轩等(2013)对三门湾和吕四近岸海域小黄鱼仔稚鱼阶段矢耳石(Sagitta)微细结构的研究发现,不同海域小黄鱼仔稚鱼耳石微结构具有显著性差异,可以用于区分不同海域的小黄鱼种群,这些研究较好地揭示了鱼类的早期生态习性和生理反应等基础科学问题。然而并不是每尾鱼耳石上均有清晰可辨的日轮,在耳石样本日轮判读过程中,这种模糊往往会导致对日轮计数的误判。日轮沉积过程中往往伴随着亚日轮结构的形成,相邻2条日轮之间亚日轮的数量与鱼的种类、生存环境及摄食情况等有关。本文以渤海小黄鱼幼鱼为研究对象,对其矢耳石轮纹结构特征进行分析,以期为小黄鱼及其他鱼种的日轮判断提供参考。
1 材料与方法
1.1 样本采集与处理
于2009年8月9日在119.25°E、37.5°N位置采集小黄鱼样品,冷藏带回实验室测定其标准体长(cm)和体重(g)。用镊子将左、右矢耳石从球状囊中轻轻取出,参照Correia等(2011)操作方法,将耳石置于2 ml离心管中,加入30%的H2O2溶液,浸泡12 h后用蒸馏水充分清洗,若耳石表面仍存有黏连组织,重复上述操作。然后,耳石于40℃烘箱中烘干,称重待用。
1.2 耳石磨片制备
实验使用Struers包埋剂EpoFix Hardener与EpoFix Resin,并将其按3∶25重量比例混匀,取适量于柱状模具中,静置12 h待其凝固,再将耳石平放于模具中,重复上述操作进行包埋。然后,用DTQTM5低速精密切割机将其切割成包有耳石的方块,用热熔胶将方块固定于载玻片,沿耳石冠状面进行研磨。研磨过程中,依次用P240、P1200、P2500和P4000规格的碳化硅砂纸研磨耳石至其核心,再用0.3 μm氧化铝粉在水绒布上对研磨好的耳石切面进行抛光,完成一面后,将耳石翻面,重复上述操作研磨抛光,直至核心区域出现清晰的轮纹。最后,将制备好的耳石切片进行标记、保存。
1.3 耳石轮纹结构观察与轮纹计数
将耳石制片置于Nikon Eclipse Ci显微镜下进行观察,分别在100×(物镜10×,目镜10×)、200×(物镜20×,目镜10×)下对耳石微细结构进行观察拍照,而后使用PhotoShop CS6软件将图片合并,获得完整的轮纹结构。随着鱼体的生长,其耳石形态亦发生变化,且结构趋于复杂,耳石长轴方向轮纹稀疏且多有轮纹紊乱不清区段,短轴方向轮纹相对密集且清晰。只有少数耳石制片的轮纹结构在长轴方向清晰可辨,多数耳石沿长轴方向进行日轮判定中需用短轴进行校正。
日轮计数参照Campana等(1985)的方法,每个样品至少计数3次,误差在10%内时取其平均值;误差大于10%的样品则摒弃。
2 结果与讨论
2.1 渤海小黄鱼矢耳石形态及横截面特征
小黄鱼矢耳石不透明,近椭圆形,前端较宽,后端较窄,外侧面有多个疣状突起(图1a),内侧面较为平滑,近背部一侧具有明显的听沟,头部近圆形,尾部为管状,呈现“蝌蚪状”(图1b)。
沿小黄鱼矢耳石冠状面研磨,研磨截面如图1c所示。由中心核到边缘的不同路径中,日轮清晰度及轮纹宽度有明显差异。由核心至第20条日轮之间,轮纹由圆环状逐渐变为椭圆。随着日龄增加,日轮也相应地呈现为不规则曲线状,可能与次生核的形成及数量有关。刘志远等(2012)对大黄鱼()矢耳石形态研究发现,在稚鱼期早期阶段,耳石长宽比迅速增加,耳石上开始形成第1个次生核,随后次生核数量逐渐增加,多次级生长区以各自次生核为中心呈扇形生长,此时耳石边缘呈现数个疣状突起而变得不规则。
2.2 渤海小黄鱼矢耳石中心核
小黄鱼矢耳石中心核呈圆形,平均直径为(24.35±0.72) μm,核内耳石原基为深黑色的圆形结构,平均直径为(12.07±0.58) μm (图1d)。大黄鱼中心核直径为(20.04±0.942) μm (刘志远等, 2012),与本研究小黄鱼中心核相近。胭脂鱼()中心核直径为(11.14±1.74) μm (雷毅等, 2017),黄唇鱼()中心核直径为(68.15±17.13) μm (区又君等, 2011),与小黄鱼中心核直径差异显著,一定程度上反映了不同鱼种间的差异性。
小黄鱼矢耳石多具有1个中心核和1个原基,个别耳石出现2个中心核和2个原基,且2个中心核的直径存在显著差别(图1e)。在草鱼()、唐鱼()、黄颡鱼()(史方等, 2006; 宋昭彬等, 2003; Tzeng, 1992)的耳石微结构中,多数为1中心核1原基,少数耳石具有1中心核2原基、2中心核2原基及3中心核3原基。目前,这些多核多原基的产生机理尚不清楚,待今后进一步研究。
2.3 渤海小黄鱼矢耳石标记轮
2.3.1 小黄鱼矢耳石第1日轮 中心核外围是明暗相间的同心环结构,暗带和明带构成的排列规整的环纹即为日轮,距中心核最近的日轮称为第1日轮(Initial daily increment),第1日轮距中心核的距离为(14.11±1.08) μm (图1d)。
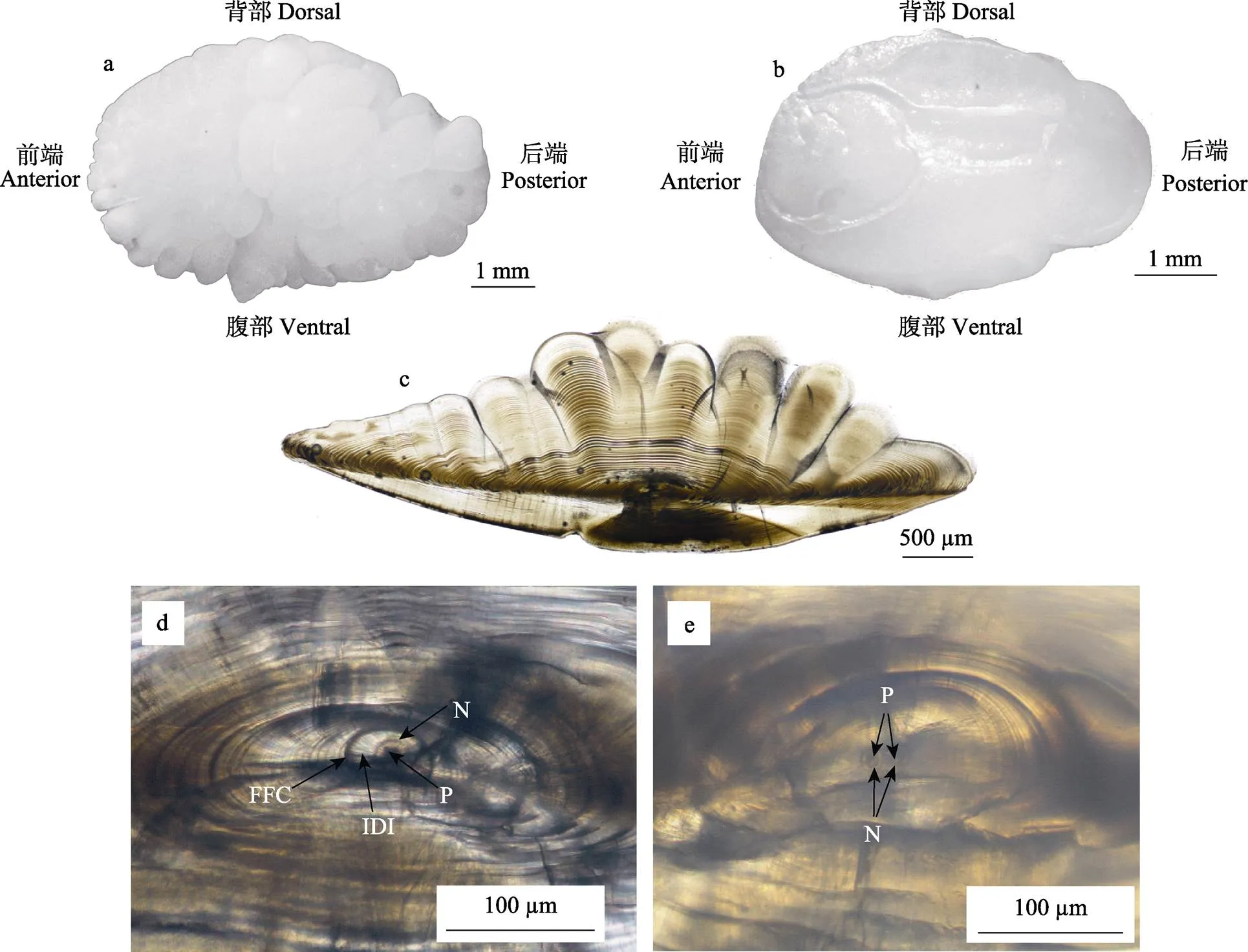
图1 小黄鱼矢耳石形态
a: 矢耳石外侧形态; b: 矢耳石内侧形态; c: 90 d小黄鱼矢耳石研磨面; d: 中心核结构; e: 2中心核2原基结构 FFC: 初次摄食标记轮; IDI: 第1日轮; N: 核心; P: 原基
a: Lateral morphology of sagittal; b: Medial morphology of sagittal; c: Grinding plane of sagittal ofof 90 d; d: Nucleus structure; e:Sagittal with two nuclei and two primordia FFC: First-feeding check; IDI: Initial daily increment; N: Nucleus; P: Primordium
黎雨轩等(2013)发现,小黄鱼矢耳石最早在胚胎发育末期出现,孵化后第2天形成第1日轮。但不同鱼种耳石第1日轮的形成时间存在差异,河鲈()第1日轮在孵化前出现,通常认为与血管、眼睛等的发育有关(Kristensen, 2008);色林错裸鲤()第1日轮于受精卵孵化当天形成(Ding, 2015);骨唇黄河鱼()仔鱼出膜后第2天形成第1日轮(严太明等, 2014),与小黄鱼第1日轮形成时间一致;但亦有多种鱼耳石上第1轮纹的形成发生在仔鱼初次摄食时,如竹荚鱼() (Jordan, 1994)。第1日轮的确定是推断鱼类个体日龄的基础,是鱼类早期生活史研究过程中重要的组成部分。
2.3.2 小黄鱼矢耳石初次摄食标记轮 本研究中小黄鱼矢耳石前3条日轮的暗带颜色较浅,第4条轮纹的暗带颜色较深、清晰可辨,推测该轮纹为初次摄食标记轮(王菲等, 2010)。该标记轮距中心核的距离为(20.67±2.28) μm (图1d)。詹炜等(2016)对小黄鱼早期发育发现,4日龄时仔鱼口裂形成,开始第1次摄食,该论述与本研究结论相吻合。小黄鱼仔鱼开口后,鱼体处于由内源性营养向外源性营养过渡时期,而初次摄食标记轮的出现,可能是由于营养物质成分的变化而使其遭遇某种压力,导致耳石上形成特殊标记轮。由于环境压力及特殊生活史的影响,鱼体内会产生相应的生理变化,从而形成记载鱼体生长和发育过程的标记轮,如孵化标记轮、初次摄食标记轮、变态标记轮以及放流标记轮等(Cieri, 2001)。因此,辨别耳石上的特殊标记轮,对分析鱼体生长发育过程中经历的事件及持续时间,发挥有力的佐证作用。
2.4 小黄鱼日龄组成与轮纹宽度
对67尾样品鱼的左、右矢耳石进行磨片观察,只有20枚矢耳石在长轴方向具有较为清晰的轮纹结构,利于轮纹宽度分析。生长轮宽度的变化也反映鱼类的早期生活史,小黄鱼矢耳石前85条轮纹在最长轴方向的宽度变化如图2所示。中心核外前20条日轮,轮纹由圆环状逐渐变为椭圆,轮纹平均宽度的变化范围为6.51~14.37 μm,且随着日龄的增加而逐渐变宽;20~40条日轮间轮纹宽度呈显著升高,该阶段小黄鱼应处于变态期(詹炜等, 2016),鱼体各结构发育完善,活动能力逐渐增强,并伴随摄食能力的提高,鱼体处于快速生长期,因而耳石沉积率变大,最终导致轮纹宽度的显著增加。40~85条日轮的宽度变化相对平稳,56~60条轮纹处出现轮纹,平均宽度最大值为41.59 μm。大西洋鳕()矢耳石轮纹宽度范围为0.8~10 μm (Hüssy, 2010),日本鯷鱼()矢耳石轮纹宽度范围为2~20 μm (Ryuji, 2008),这应为种间差异所致。
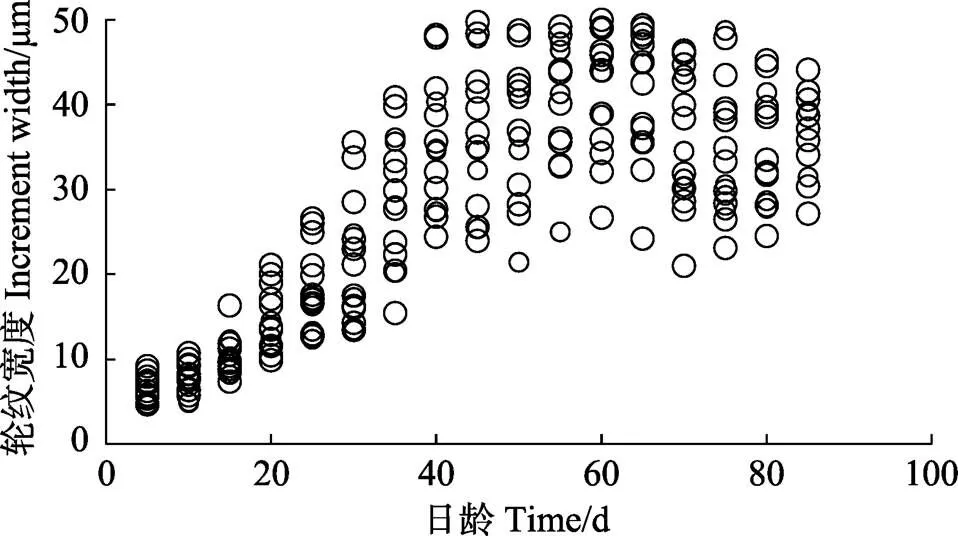
图2 渤海小黄鱼矢耳石前85条轮纹宽度变化
2.5 小黄鱼矢耳石亚日轮结构
耳石日轮沉积过程中,伴随有亚日轮的形成,日轮暗带排列规整、颜色深,亚日轮暗带宽度不规律、颜色较浅。亚日轮宽度与日轮宽度密切相关,日轮宽度大,亚日轮宽度亦随之增大。如图3a所示,由核心到边缘的测量路径中,A区域轮纹宽度范围为12.17~13.86 μm,亚日轮宽度为2.14~3.28 μm(图3b);B区域日轮宽度为21.89~26.72 μm,亚日轮宽度为6.00~11.67 μm(图3c)。温度、摄食频率等因素的波动在日轮基础上形成亚日轮结构,由于外界影响因子发生的强度、频率及持续时间不稳定,因此,相邻2条日轮之间形成的亚日轮数量、宽度及明暗度也不同(Campana, 1985)。相对于日轮结构,亚日轮清晰度稍差,轮纹宽度较窄且波动较大,在显微镜下对视野轻微调焦时会暂时性“消失”,该现象同Powell等(2000)对亚日轮特性的描述一致。Dougherty(2008)在对黄线狭鳕()耳石的研究中将宽度小于0.5 μm的轮纹判定为亚日轮,但不同鱼种间耳石沉积速率差异很大,本研究中,亚日轮最大轮纹宽度为11.67 μm,该宽度显著大于小黄鱼前10条日轮的轮纹宽度,所以对于小黄鱼而言,不适合将某一轮纹宽度作为区分日轮与亚日轮的依据。
a:由核心到边缘轮纹宽度的测量路径;b:A区域的日轮结构与亚日轮结构;c:B区域的日轮结构与亚日轮结构 A:A区域;B:B区域;DI:日轮;IDI:第一日轮;N:核心;P:原基;SDI:亚日轮
a: Incremental measurement route from center to edge; b: Daily increment and sub-daily increment in area A; c: Daily increment and sub-daily increment in area B A: Zone A; B: Zone B; DI: Daily increment; IDI: Initial daily increment; N:Nucleus; P:Primordium; SDI: Sub-daily increment
本研究发现,在2种情况下亚日轮结构更为普遍。在耳石快速沉积的区域,随着日轮暗带宽度增大,亚日轮形成的可能性更大,在显微镜下也更容易看到,因此,对视野中轮纹密集且宽度变窄的区域,应轻微调焦来辨别亚日轮,减少计数过程中的误差。另外是耳石制片过程中的人为因素,耳石研磨过度有时会使亚日轮结构更为明显,大大增加了分辨二者的难度。与其他鱼种相比,小黄鱼具有较发达的耳石,日轮有机物填充带宽度大,亚日轮也更为普遍。因此,在耳石研磨过程中应注意制片的厚度,避免耳石研磨过度造成的轮纹计数误差(Stevenson, 1992)。
Arkhipkin AI, Bizikov VA. Role of the statolith infunctioning of the acceleration receptor system in squids and sepioids. Journal of Zoology, 2000, 250(1): 31–55
Campana SE, Neilson JD. Microstructure of fish otoliths. Canadian Journal of Fisheries and Aquatic Sciences, 1985, 42(5): 1014–1103
Cieri MD, McCleave JD. Validation of daily otolith increments in glass-phase American eels(Lesueur) during estuarine residency. Journal of Experimental Marine Biology and Ecology, 2001, 257(2): 219–227
Correia AT, Pipa T, Gonalves JMS,. Insights into population structure ofalong the SW Portuguese coast from otolith elemental signatures. Fisheries Research, 2011, 111(1/2): 82–91
Ding CZ, Chen YF, He DK,. Validation of daily increment formation in otoliths forin the Tibetan Plateau. China Ecology and Evolution, 2015, 16(5): 3243–3249
Dougherty AB. Daily and sub-daily otolith increments of larval and juvenile walleye pollock,(Pallas), as validated by alizarin complexone experiments. Fisheries Research, 2008, 90: 271–278
Hislop JRG, Gallego A, Heath MR,. A synthesis of the early life history of the anglerfish,(Linnaeus, 1758) in northern British waters. ICES Journal of Marine Science, 2001, 58: 70–86
Hüssy K, Hinrichsen HH, Fey DP,. The use of otolith microstructure to estimate age in adult Atlantic cod. Journal of Fish Biology, 2010, 76(7): 1640–1654
Jin XS. Ecology and population dynamics of small yellow croaker () in the Yellow Sea. Journal of Fishery Sciences of China, 1996, 3(1): 32–46 [金显仕. 黄海小黄鱼()生态和种群动态的研究. 中国水产科学, 1996, 3(1): 32–46]
Jordan AR. Age, growth and back-calculated birth date distributions of larval jack mackerel,(Pisces: Carangidae), from eastern Tasmanian coastal waters. Australian Journal of Marine and Freshwater Research, 1994, 45(1): 19–33.
Kristensen PB, Closs G, Lokman M,. Otolith formation, microstructure and daily increment validation in juvenile perch. Journal of Fish Biology, 2008, 73(6): 1478–1483
Lei Y, Zhou Y, Yang K,. Otolith microstructure character analysis between wild-caught and hatchery-breeding. Sichuan Journal of Zoology, 2017, 36(3): 285–292 [雷毅, 周宇, 杨坤, 等. 野外采集和人工饲养胭脂鱼耳石微结构特征分析. 四川动物, 2017, 36(3): 285–292]
Li YX, Tang JH, Xu XM,Comparision of otolith microstructures in small yellow croaker larvae and juveniles from Sanmen Bay and Lusi. Marine Fisheries, 2013, 35(4): 423–431 [黎雨轩, 汤建华, 徐献明, 等. 三门湾和吕四近岸小黄鱼仔稚鱼耳石微结构特征比较. 海洋渔业, 2013, 35(4): 423–431]
Liu ZY, Li SF, Xu XM,. Morphological development and microstructure of sagittal otolith of large yellow croaker,during larval and early juvenile stages. Journal of Fishery Sciences of China, 2012, 19(5): 863–871 [刘志远, 李圣法, 徐献明, 等. 大黄鱼仔稚鱼不同发育阶段矢耳石形态发育和微结构特征. 中国水产科学, 2012, 19(5): 863–871]
Lu HJ, Chen XJ, Fang Z,. Statolith microstructure and growth characteristics ofin the southwest Atlantic Ocean. Progress in Fishery Sciences, 2012, 33(3): 15–25 [陆化杰, 陈新军, 方舟, 等. 西南大西洋阿根廷滑柔鱼耳石微结构及生长特性. 渔业科学进展, 2012, 33(3): 15–25]
Ou YJ, Liao R, Li JE,. The morphology of otolith and its characteristics of microstructure in. Guangdong Agricultural Sciences, 2011, 38(12): 123–124, 148 [区又君, 廖锐, 李加儿, 等. 黄唇鱼的耳石形态和微结构特征. 广东农业科学, 2011, 38(12): 123–124, 148]
Powell AB, Laban EH, Holt SA. Validation of age estimates from otoliths of larval and juvenile spotted seatrout,. Fishery Bulletin, 2000, 98(7): 650–654
Ryuji Y, Ichiro A, Isamu M. Daily age of adult Japanese anchovyoff eastern Honshu, Japan by otolith daily increment. Fisheries Science 2008, 74(6): 1348–1350
Shi F, Sun J, Lin XT,. Otolith ontogeny and increment formation in larval. Chinese Journal of Zoology, 2006, 41(4): 10–16 [史方, 孙军, 林小涛, 等. 唐鱼仔鱼耳石的形态发育及日轮. 动物学杂志, 2006, 41(4): 10–16]
Song ZB, Cao WX. Dimensions of otolith primordia and nuclei in larval grass carp and silver carp. Journal of Sichuan University (Natural Science), 2003, 40(4): 774–777 [宋昭彬, 曹文宣. 草鱼和鲢仔鱼耳石原基和中心核特征的研究. 四川大学学报: 自然科学版, 2003, 40(4): 774–777]
Stevenson DK, Campana SE. Otolith microstructure examination and analysis. Journal of Experimental Marine Biology and Ecology, 1993, 171(2): 297–298
Toshihiko S, Toshiki K, Jiro S,. Otolith microstructure of chum salmon: Formation of sea entry check and daily deposition of otolith increments in seawater conditions. Fisheries Science, 2007, 73: 27–37
Tzeng WN, Yu SY. Effects of starvation on the formation of daily growth increments in the otoliths of milkfish(F.) larvae. Journal of Fish Biology, 1992, 40(1): 39–48
Wang F, Cheng JY, Luo HZ. Relationship between daily-growth increment of otolith and growth of Japanese parrotfish () Journal of Fishery Sciences of China, 2010, 17(2): 320–328 [王菲, 程家骅, 罗海忠. 东海条石鲷仔鱼耳石日轮与生长的关系. 中国水产科学, 2010, 17(2): 320–328]
Xiong Y, Liu HB, Liu PT,. Reconstructing habitat history ofin Lusi coastal waters of Jiangsu Province, China based on otolith microchemistry. Chinese Journal of Applied Ecology, 2014, 25(3): 836–842 [熊瑛, 刘洪波, 刘培廷, 等. 基于耳石元素微化学的江苏吕泗近岸小黄鱼生境履历重建. 应用生态学报, 2014, 25(3): 836–842]
Yan TM, Hu JX, Yang T,. Study on the otolith development and the formation of increments in larvae and juvenile of. Acta Hydrobiologica Sinica, 2014, 38(4): 764–771 [严太明, 胡佳祥, 杨婷, 等. 骨唇黄河鱼耳石早期形态发育和轮纹特征研究. 水生生物学报, 2014, 38(4): 764–771]
Zhang GZ, Li XS, Zhu JC,. The growth characteristics of small yellow croaker(Bleeker, 1987) underyearling in the central and southern Yellow Sea. Progress in Fishery Sciences, 2010, 31(3): 15–22 [张国政, 李显森, 朱建成, 等. 黄海中南部小黄鱼当年幼鱼生长特征的研究. 渔业科学进展, 2010, 31(3): 15–22]
Zhan W, Lou B, Chen RY,. Observation of embryonic, larva and juvenile development of small yellow croaker,. Oceanologia et Limnologia Sinica, 2016, 47(5): 1033–1039 [詹炜, 楼宝, 陈睿毅, 等. 小黄鱼()胚胎发育及仔、稚鱼形态特征观察. 海洋与湖沼, 2016, 47(5): 1033–1039]
Sagitta Microstructure Characteristics of Young of the Year (YOY)in the Bohai Sea
ZHANG Tingting1, WANG Yukun1, YUAN Wei1, JIN Xianshi1,2, CHEN Chao1, SUN Yao1,2①
(1. Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Science, Qingdao 266071; 2. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266071)
Sagitta microstructure of young of the yearcollected in the Bohai Sea was studied, which is routinely used for age analyses. Results showed that there was usually one circular nucleus with one deeply black primordium in it, and only a few samples presented two nuclei or primordia. Average diameter of sagitta nucleus and primordia were (24.35±0.72) μm and (12.07± 0.58) μm, respectively.The first increment was observed on the second day, with a distance of (14.11± 1.08) μm to the central nucleus. The first feeding check appeared on the 4th increment, with a distance of (20.67±2.28) μm to the central nucleus, displaying larger width,deeper color, and higher clarity.From the otolith nucleus to approximately the 20th increment, increments gradually changed from a ring shape to an ellipse. Increment widths in the long axis ranged from 6.51 μm to 14.37 μm and tended to increase with age. Increment widths dramatically rose until approximately the 40th increment, reaching a value of (36.05±8.32) μm. Thereafter, increment mean width did not show any clear trend.The process of daily increment deposition ofwas accompanied by the formation of sub-daily increments. Sub-daily increment deposits displayed a lack of periodicity, irregularity, low clarity, and incompleteness compared with daily increments. Changing the depth of the field while observing these less defined increments resulted in a temporary elimination of the sub-daily increments from view. The analysis of otolith increment deposition has been widely applied and developed in age validation of larvae and juvenile fishes. The counting results directly determine analytical accuracy of the hatching date, spawning field distribution, and migration of wild individuals. The numerous presence of sub-daily increments will increase the difficulty of discriminating daily increments.has well-developed sagitta, which possesses greater width of daily increment and a greater number sub-daily increments. Therefore, distinguishing daily increments from sub-daily increments is the primary task of the research on the sagitta microstructure ofIn the present study, the sagitta microstructure characteristics ofwere analyzed to provide a reference for daily increment judgment of wildand other fish species. Meanwhile,this study will supplement the biological information onand provide data for resource surveys of wild populations.
; Sagitta; Daily increment; Microstructure
S917.4
A
2095-9869(2020)02-0035-06
孙 耀,研究员,E-mail: sunyao@ysfri.ac.cn
2019-02-21,
2019-06-04
2019-03-14
* 国家重点基础研究发展计划(2015CB453303)和国家自然科学基金青年科学基金项目(41606195)共同资助 [This work was supported by the National Basic Research Program of China (2015CB453303), and the Youth Program of National Natural Science Foundation of China (41606195)]. 张廷廷,E-mail: 13256879033@163.com
10.19663/j.issn2095-9869.20190221001
http://www.yykxjz.cn/
张廷廷, 王玉堃, 袁威, 金显仕, 陈超, 孙耀. 渤海当年生小黄鱼矢耳石微结构特征. 渔业科学进展, 2020, 41(2): 35–40
Zhang TT, Wang YK, Yuan W, Jin XS, Chen C, Sun Y. Sagitta microstructure characteristics of young of the year (YOY)in the Bohai Sea. Progress in Fishery Sciences, 2020, 41(2): 35–40
SUN Yao, E-mail: sunyao@ysfri.ac.cn
(编辑 冯小花)
猜你喜欢
小猕猴学习画刊(2021年9期)2021-10-11
中国食用菌(2021年3期)2021-05-08
植物研究(2021年1期)2021-02-26
中老年保健(2020年3期)2020-12-09
小读者(2020年4期)2020-06-16
老友(2018年7期)2018-08-08
养生保健指南(2017年12期)2017-12-07
科学生活(2017年9期)2017-10-16
作文与考试·小学高年级版(2017年9期)2017-05-16
食品与生活(2016年10期)2016-10-13
