
干旱胁迫及复水下朱槿生长及根际土壤微生物多样性的变化特征
2020-03-23 06:07黄旭光秦玲黄丽丹陆炎松罗恩波黄玲璞刘岳飞
热带作物学报 2020年2期
黄旭光 秦玲 黄丽丹 陆炎松 罗恩波 黄玲璞 刘岳飞


摘 要:以朱槿品种‘紫牡丹为试验材料,采用盆栽试验,研究了苗期重度干旱胁迫及复水对其根系生理特性的影响,并运用高通量测序技术分析干旱胁迫及复水下朱槿根际土壤微生物多样性的变化特征。结果显示:与正常处理(CK)相比,重度干旱胁迫及复水处理(T2)的总根长、根表面积及根尖数等根系生长指标呈现显著减少的结果,但与重度干旱胁迫(T1)处理相比,均显著增加。通过对朱槿根际土壤样品中细菌群落结构的研究发现,属分类水平上,厌氧绳菌属(Anaerolineaceae),酸杆菌属(Acidobacteria)、酸微菌属(Acidimicrobiales)、幽门螺杆菌属(Sphaerobacter)、链霉菌属(Streptomyces)等是不同处理土壤样品中的优势菌群。T1处理下的嗜青霉属(Algiphilus)、奥利螺旋菌属(Aureispira)、绿线菌属(Chloronema)、扁棒壳属(Acrospermum)为不同处理下的特殊细菌。真菌方面,属的分类水平上发现,粪壳菌属(Sordariale)、散囊菌属(Eurotiales)、拟青霉属(Pseudallescheria)、格孢腔菌属(Pleosporales)、枝孢霉属(Cladosporium)等是不同处理土壤样品中的优势菌群;T1和T2处理下的样品中,侧角藻属(Goniomonas)、齿盘菌属(Lamprospora)和索罗迪夫斯菌属(Sorodiplophrys)为不同处理下的特殊真菌属。其中,有促生作用的细菌如酸微菌属( Acidimicrobium)、厌氧绳菌属(Anaerolineaceae)嗜青霉属(Algiphilus)以及具有一定生防作用真菌如拟青霉属(Pseudallescheria)、枝孢霉属(Cladosporium)等均可以合理的应用在农业生产中,为植物的抗干旱胁迫以及提高朱槿在干旱胁迫下的水分利用做出贡献。
关键词:干旱胁迫;高通量测序;朱槿;根系;微生物多样性
中图分类号:S154.3 文献标识码:A
Abstract: A pot experiment using a cultivar ‘Purple Peony as the experimental material was conducted under drought and rehydration condition. The effects of drought at seedling stage and rehydration on the physiological characteristics of roots and the soil microbial diversity in the rhizoshere were studied. The total root length, root surface area and root tip numbers were significantly decreased in the drought and rehydration treatment (T2), compared to the normal condition (CK), but they were significantly increased compared with the drought stress without rehydration (T1). Under drought stress (T1), Algiphilus, Aureispira, Chloronema, Acrospermum were the special bacteria. Anaerolineaceae, Acidobacteria, Acidimicrobiales, Sphaerobacter and Streptomyces were the dominant bacteria in the rhizoshpres of all drought and normal treatments. Sordariale, Eurotiales, Pseudallescheria, Pleosporales and Cladosporium were the dominant fungi. Goniomonas, Lamprospora and Sorodiplophrys were the special fungi in drought stress (T1) and rehydration after drought stress (T2). Bacteria such as Acidimicrobium, Anaerolineaceae, Algiphilus, Pseudallescheria, Cladosporium, having certain biocontrol effects, could be reasonably used in agricultural production, which could contribute to the resistance of plants to drought stress and the improvement of water use of Hibiscus under drought stress.
Keywords: drought stress; high-throughput sequencing; Hibiscus; root system; microorganisms diversity
干旱一直以來是人类面临的主要自然灾害之一,随着人类的经济发展和人口膨胀,人类对水资源的污染越来越严重,水资源短缺的现象也日趋明显。此外,全球暖干化进程的加快,水资源短缺的形势愈发严峻,对农业和生态环境的可持续发展造成了严重的影响[1]。这也直接导致了全世界范围内干旱地区的扩大以及干旱化程度的加重,但干旱化问题也越来越引起全球范围内的广泛关注。
2 结果与分析
2.1 干旱胁迫及复水下的朱槿根系生长指标
由表1可知,与CK相比,T1的朱槿總根长、总根表面积及总根体积及根尖数均呈显著性减少的结果,而对于根均直径的影响不显著。T2的根均总根表面积及总根体积及根尖数与CK相比均有显著的减少,但与T1处理相比,均显著增加。与T1相比,T2和CK的总根长分别增加了29.41%和63.73%,总根表面积分别增加49.07%和90.66%,总根体积分别增加了71.43%和128.57%,根尖数增加量分别为33.04%和65.28%。以上结果说明,干旱胁迫对朱槿植株根系的总根长、总根表面积、总根体积及根尖数有显著的抑制作用,而干旱后复水对根系生长也有显著的补偿效应。
2.2 干旱胁迫及复水处理下朱槿幼苗生长指标
与CK相比,T1朱槿幼苗的株高及鲜重(地上部、根系)等指标均显著减少,株高、地上部和根系分别减少了73.25%、55.01%和63.63%;而干旱胁迫后复水处理对于地径、鲜重(地上部、根系)、及根冠比等指标并没有显著差异。3种不同处理对于朱槿幼苗的地径和根冠比等影响均没有显著的差异。由表2可知,干旱胁迫后复水对株高的补偿效应较明显,达到与CK相近的效果,而对于地径和根冠比等指标则没有显著的影响。
2.3 干旱胁迫及复水处理对朱槿根际土壤微生物类群的相关分析
以不同水分处理土壤样品分布的属分类水平
数为依据,用Mothur软件对各不同处理的土壤样品微生物多样性进行相关分析,构建Venn图(图1)。Venn图可展现多个样品中所共有的属分类水平数目,属分类水平数目反映环境样品的相似性及重叠情况。由图1可知,不同水分处理下的土壤中细菌的共有优势细菌属为594,占总细菌属的90.69%。T1的特有优势细菌属最多,占总细菌属的1.98%,而CK处理与T2的特有优势细菌属仅分别占总细菌属的0.46%和0.76%。真菌方面,不同水分处理下的朱槿根际土壤中真菌的共有优势真菌属为94,占总优势真菌属的68.12%。其中,T2的特有优势真菌属最多,占总优势真菌属的7.97%,而CK与T1的特有优势真菌属分别为4.35%和2.17%。
2.4 干旱胁迫及复水处理下朱槿土壤微生物类群特征
通过高通量测序技术对土壤样品微生物中的细菌部分进行V3~V4区测序,不同处理土壤样品共获得有效序列364 442条,片段长度在401~ 460 bp,测序覆盖度在98.27%~98.88%之间。基于其测序深度基本能够反映该区域所涵盖的细菌群落的结构以及种类等特性,可进行定量比较群落组成以及物种多样性的相对差异。
分析朱槿根际土壤样品细菌多样性发现,属分类水平上(图2),厌氧绳菌属(Anaerolineaceae)为朱槿根际土壤细菌的优势菌属,相对含量平均值约占总群落的8.30%~9.65%。其中,CK土壤中的相对丰度最低,而T2土壤中的相对丰度最高。此外,酸杆菌属(Acidobacteria)、酸微菌属(Acidimicrobiales)、幽门螺杆菌属(Sphae?robacter)、链霉菌属(Streptomyces)等也是不同处理朱槿根际土壤中的优势菌群。其中,CK的土壤样品中酸微菌属(Acidimicrobiales)的相对丰度占比最高,T1的土壤样品中的幽门螺杆菌属(Sphaerobacter)的相对丰度占比最低,说明水分含量的降低在一定程度上有降低幽门螺杆菌的作用。而T2的土壤样品中,热丁香类诺卡氏菌(Nocardioides)属的相对丰度占比最高,热丁香类诺卡氏菌(Nocardioides)最早分离自越南南部土壤,诺卡氏菌属成员的特征是革兰氏阳性,非酸性,好氧菌,过氧化氢酶阳性,横隔断裂成球状或棒状,是一种嗜热菌属[15]。此外,与CK相比,酸杆菌属(Acidobacteria)、链霉菌属(Strep?tomyces)等其他细菌群落在不同处理下相对丰度占比均发生一定程度的变化,但变化效果不显著。另一方面,T1的嗜青霉属(Algiphilus)、奥利螺旋菌属(Aureispira)、绿线菌属(Chlor?onema)、扁棒壳属(Acrospermum)为不同处理下的特殊真菌属。
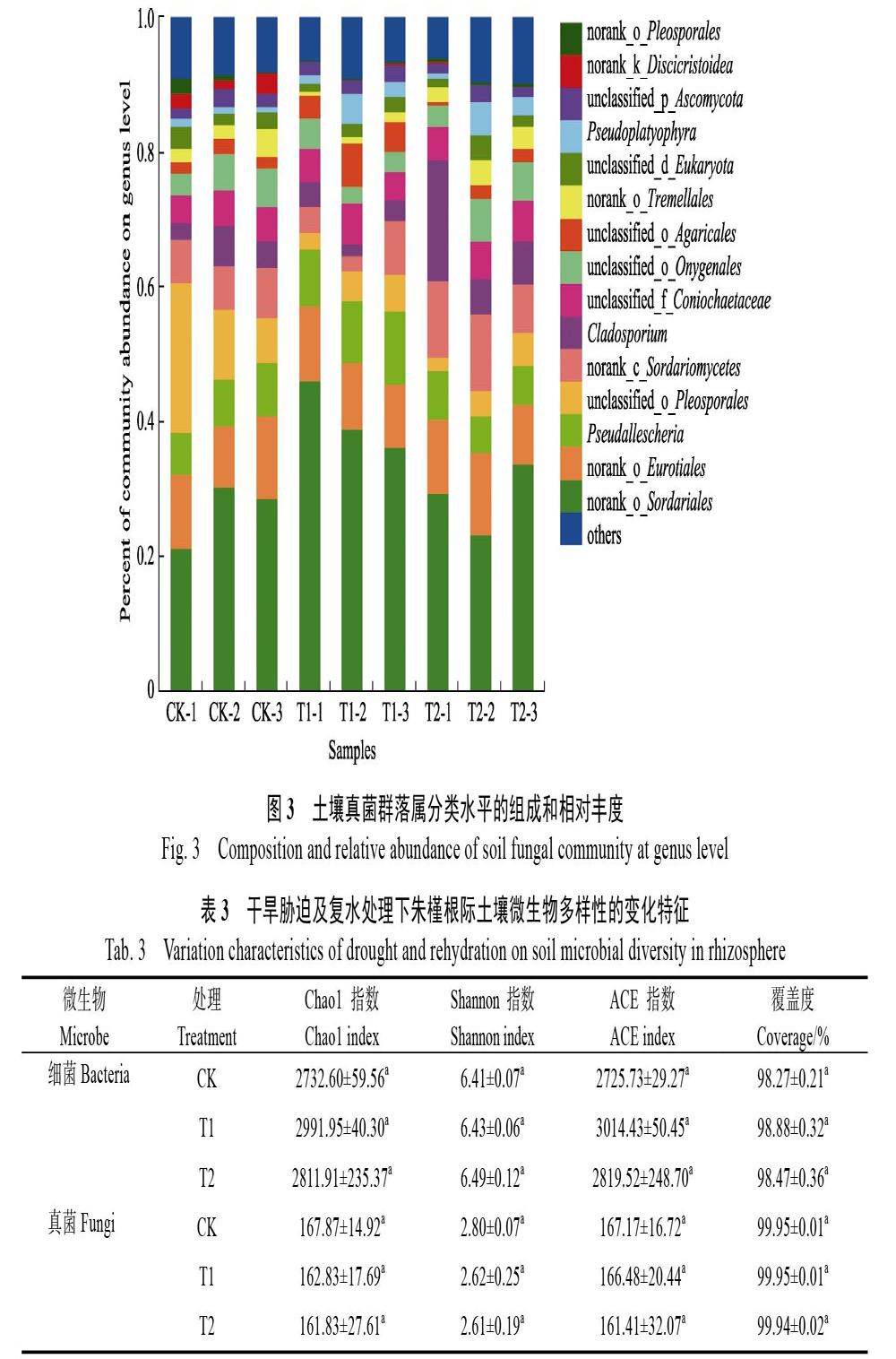
通过高通量测序技术对土壤样品微生物中的真菌部分进行V5~V7区测序,不同处理土壤样品共获得有效序列350 138条,片段长度在361~ 420 bp,测序覆盖度在99.54%~99.95%之间。分析朱槿根际土壤中真菌多样性发现:属分类水平上(图3),粪壳菌属(Sordariale)为朱槿根际土壤的优势真菌属。其中,以T1的根际土壤中相对丰度最高,占样品总丰度的40.38%,而CK与T2的相对丰度占比分别为27.24%和29.16%。此外,T1的拟青霉属(Pseudallescheria)、粪壳菌属(Sordariomycetes)、枝孢霉属(Clados?porium)等相对丰度占比均高于CK及T2处理。格孢腔菌属(Pleosporales)的相对丰度占比在CK的土壤样品中最高、占样品总相对丰度的12.73%,是其他2种处理下相对丰度占比的3.10~3.59倍。此外,与CK相比,干旱胁迫下,散囊菌属(Eur?o?tiales)、分生孢子虫属(Coniochaetaceae)等其他部分真菌群落的相对丰度占比并没有显著的变化。另一方面,T1和T2的样品中侧角藻属(Gonio?monas)、齿盘菌属(Lamprospora)和索罗迪夫斯菌属(Sorodiplophrys)为不同处理下的特殊真菌属。
2.5 干旱胁迫及复水处理下朱槿根际土壤微生物多样性的变化特征
由表3可知,与CK相比,干旱胁迫及复水处理下的朱槿土壤微生物的Chao1指数、Shannon指数、ACE指数及覆盖率均没有显著的变化。干旱胁迫及复水处理下朱槿根际土壤微生物的丰富度和多样性没有显著的变化。
3 讨论
逆境条件下,作物会通过体内一系列的生理生化变化去适应周围的环境,而其变化最终体现在植株外部形态上。干早脅迫限制了作物根系和地上部的生长,降低了作物的干物质积累[16]。与CK相比,T1的朱槿总根长、总根表面积、总根体积、根尖数以及朱槿幼苗的株高及鲜重(地上部、根系)等指标均显著减少,T2的总根表面积、总根体积及根尖数以及朱槿幼苗的株高及鲜重(地上部、根系)等指标与CK相比也是有一定程度的减少,但与T1相比,均显著增加,产生了较明显的补偿效应[17]。有研究表明,植物根系是植物直接吸收水分的重要器官,对植物的抗旱功能具有至关重要的作用。发达的根系系统可使植物充分吸收利用贮存在土壤中的水分,使植物度过干旱期。与地上部茎、叶生长反应相比较,干旱胁迫对根系生长的影响相对较小,结果使根冠比提高[18-19],而本研究中根冠比在不同处理下并没有显著变化,可能与试验处理时间及植物自身对逆境的补偿机制有关,具体原因还需要更深入的研究。
本研究基于高通量测序技术,分析朱槿根际土壤细菌多样性发现,属分类水平上,厌氧绳菌属(Anaerolineaceae)相对丰度在CK处理中最低,但在T2处理中相对丰度最高。厌氧绳菌属(Anaerolineaceae)细菌主要参与烷烃的产甲烷降解过程,烷烃最初可能通过添加富马酸被激活,被分解为脂肪酸,然后转化为乙酸[20]。此外,在不同处理条件下发现了通常在岩石中才被发现的且能氧化金属硫化物酸微菌属(Acidimicrobium)的相应序列,揭示出酸微菌亚纲(Acidimicrobidae)中还存在大量未知的放线菌新类群[21]。幽门螺杆菌属(Sphaerobacter)是一种微厌氧、化学异养、过氧化氢酶-阳性的嗜热链球菌[22]。本研究发现,不同水分处理下对土壤中的幽门螺杆菌属(Sph?aerobacter)的丰度比例没有显著地变化。
对朱槿根际土壤真菌多样性分析发现,粪壳菌属(Sordariale),拟青霉属(Pseudallescheria)、枝孢霉属(Cladosporium)真菌在T1的相对丰度占最高;格孢腔菌属(Pleosporales)真菌在CK的相对丰度最高。部分研究已表明,拟青霉属(Pseudallescheria)是一些植物寄生线虫的重要天敌,能够寄生于卵,也能侵染幼虫和雌虫,可明显减轻多种作物根结线虫、胞囊线虫、茎线虫等植物线虫病的危害,是1种有潜力的生防真菌[23]。枝孢霉属(Cladosporium)在植物受到重金属污染时,可减轻Hg2+和Cd2+等金属离子对植物的的毒害作用[24]。
T1的嗜青霉属(Algiphilus)为不同处理的特殊细菌属,该属菌株最早是从海洋甲藻(CCAP 1121/2)的实验室培养中分离到一株严格耐氧、耐盐性强的革兰氏阴性杆状细菌,其能够降解二和三环多环芳烃。该属菌株显示出较窄的营养谱,更优先食用脂族和芳烃化合物,以及少量有机酸[25],对缓解土壤中多环芳香烃的危害有一定的作用。此外,奥利螺旋菌属(Aureispira)、绿线菌属(Chlo?ronema)、扁棒壳属(Acrospermum)也是T1处理的特殊细菌属。另外,T1和T2处理的样品中侧角藻属(Goniomonas)、齿盘菌属(Lamprospora)和索罗迪夫斯菌属(Sorodiplophrys)为不同处理下的特殊真菌属。
由于全球气候变化,长期干旱、局部地区缺水的极端事件可能会进一步增加。为了应对其造成的恶化影响,需要采取广泛的适应和缓解战略。但传统的策略的制定时间长、成本高,需要开发简单、低成本的生物方法来管理非生物胁迫。植物相关微生物对非生物胁迫具有重要的抗逆性,其生物可能包括轮藻面、内生细菌和共生真菌,并通过触发渗透反应和诱导植物中新基因等多种机制运作。通过基因工程和植物育种开发抗逆性品种是必要的,但这是一个漫长的过程,而微生物接种以减轻植物的压力是一种成本效益更高的环境友好型选择,可在较短的时间内获得[26-29]。如果能利用微生物对极端生物的耐受性、其普遍性、遗传多样性、微生物与作物的相互作用以及开发其在农业生产中成功应用的方法,就可发挥微生物重要的作用[30-31]。微生物除了通过产生胞外多糖和形成生物膜来影响根际土壤的理化性质外,还可通过诱导保护剂和热休克蛋白等不同的机制,影响高等植物对非生物胁迫的响应[32]。本研究中,干旱胁迫下的朱槿土壤微生物的Chao1指数、Shannon指数、ACE指数及覆盖率均没有显著的变化,但与CK相比,T1的朱槿总根长、总根表面积、总根体积、根尖数以及朱槿幼苗的株高及鲜重(地上部、根系)等指标均显著减少。其结果可能与微生物和植物对水分胁迫条件下的适应性以及水分胁迫的临界值不同有关。具有促生作用的细菌如酸微菌属(Acidimicrobium)、厌氧绳菌属(Anaerolineaceae)等,以及具有一定生防作用真菌如拟青霉属(Pseudallescheria)、枝孢霉属(Cladosporium)和嗜青霉属(Algiphilus)等均可合理的应用在农业生产中,为植物的抗干旱、提高植物在干旱胁迫下对水分的利用率等作出贡献。此外,本研究还可对植物在冷害、盐害、金属毒性和高温等非生物胁迫下的生存难题提供一个以微观世界的角度来解决宏观问题的方向。
参考文献
Schutter M E, Sandeno J M, Dick R P. Seasonal, soil type, alternative management influences on microbial communities of vegetable cropping systems[J]. Biology and Fertility of Soils, 2001, 34: 397-410.
陈甲林, 史佑海, 梁伟红. 海南扶桑品种资源调查及其园林应用研究[J]. 热带农业科学, 2009, 29(3): 24-28.
嵇 含. 南方草木状[M]. 广州: 广东科技出版社, 2009.
王光华, 金 剑, 徐美娜, 等. 植物、土壤及土壤管理对土壤微生物群落结构的影响[J]. 生态学杂志, 2006, 25(5): 550-556.
刘玮琦, 茆振川, 杨宇红, 等. 应用16S rRNA基因文库技术分析土壤细菌群落的多样性[J]. 微生物学报, 2008, 48(10): 1344-1350.
Elsas J D V, Garbeva P, Salles J. Effects of agronomical measures on the microbial diversity of soils as related to the suppression of soil of soil borne plant pathogens[J]. Biodegradation, 2002, 13(1): 29-40.
Blum A. Effective use of water (UEW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress[J]. Field Crops Research, 2009, 112(2): 119-123.
Shane M W, McCully M E, Canny M J, et al. Seasonal water relations of Lyginia barbata (Southern rush) in relation to root xylem development and summer dormancy of root apices[J]. New Phytologist, 2010, 185(4): 1025-1037.
Kato Y, Okami M. Root growth dynamics and stomatal beha?viour of rice (Oryza sativa L.) grown under aerobic and floo?ded conditions[J]. Field Crops Research, 2010, 117(1): 9-17.
Hsiao T C. Plant responses to water stress[J]. Annual Review of Plant Physiology, 1973, 24: 519-570.
黎 裕. 作物抗旱鉴定方法与指标[J]. 干旱地区农业研究, 1993(1): 91-99
Riley D, Barber S A. Salt accumulation at the soybean (Glycine max (L.) Merr.) root-soil interface[J]. Proceedings Soil Science Society of America, 1970, 34 (1): 154-155.
张志良, 瞿伟菁. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2003.
戴雅婷, 闫志坚, 解继红, 等. 基于高通量测序的两种植被恢复类型根际土壤细菌多样性研究[J]. 土壤学报, 2017, 54(3): 735-748.
Lee D W, Lee A H, Lee H, et al. Nocardioides litoris sp. nov. isolated from the Taean seashore[J]. International Journal of Systematic and Evolutionary Microbiology, 2017, 67(7): 2332-2336.
Wassmann R, Jagadish S V K, Heuer S, et al. Chapter 2 Climate change affecting rice production: The physiological and agronomic basis for possible adaptation strategies[J]. Advances in Agronomy, 2009, 101: 59-122.
趙丽英, 邓西平, 山 仑. 水分亏缺下作物补偿效应类型及机制研究概述[J]. 应用生态学报, 2004(3): 523-526.
Lotter J de V Beukes D L,Weber H W. Growth and quality of apples as affected by different irrigation treatments[J]. Journal of Horticultural Science, 1985, 60(2): 181-192.
李绍华. 果树生长发育、产量和果实品质对水分胁迫反应的敏感期及节水灌溉[J]. 植物生理学通讯, 1993, 29(1): 10-16.
Liang B, Wang L Y, Mbadinga S M, et al. Anaerolineaceae and Methanosaeta turned to be the dominant microorganisms in alkanes-dependent methanogenic culture after long-term of incubation[J]. AMB Express, 2015, 5(1): 37.
張 娇, 夏占峰, 贺江舟, 等. DNA提取方法对盐环境放线菌多样性分析的影响[J]. 微生物学报, 2013, 53(7): 746-757.
Hugenholtz P, Stackebrandt E. Reclassification of Sphaerobacter thermophilus from the subclass Sphaerobacteridae in the phylum Actinobacteria to the class Thermomicrobia (emended description) in the phylum Chloroflexi (emended description)[J]. International Journal of Systematic and Evolutionary Microbiology, 2004, 54(6): 2049-2051.
王昌家, 宋超英, 张新德, 等. 淡紫拟青霉菌料防治大豆胞囊线虫的后效研究[J]. 中国生物防治, 1997 (1): 27-29.
安红梅. 三峡库区耐淹植物内生真菌重金属抗性研究[D]. 重庆: 西南大学, 2015.
Gutierrez T, Green D H, Whitman W B, et al. Algiphilus aromaticivorans gen. nov., sp. nov., an aromatic hydrocarbon-degrading bacterium isolated from a culture of the marine dinoflagellate Lingulodinium polyedrum, and proposal of Algiphilaceae fam. nov[J]. International Journal of Systematic and Evolutionary Microbiology, 2012, 62(11): 2743-2749.
Grover M, Ali S Z, Sandhya V, et al. Role of microorganisms in adaptation of agriculture crops to abiotic stresses[J]. World Journal of Microbiology and Biotechnology, 2011, 27(5): 1231-1240.
Blanco, Bernard T. Osmoadaptation in rhizobia: ectoine- induced salt tolerance[J]. Journal of Bacteriology, 1994, 176(17): 5210-5217.
McLellan C A, Turbyville T J, Wijeratne E M, et al. A rhizosphere fungus enhances Arabidopsis thermotolerance through production of an HSP90 inhibitor[J]. Plant Physiol, 2007, 145(1): 174-182.
Neto D, Carvalho LM, Cruz C, et al. How do mycorrhizas affect C and N relationships in flooded Aster trifolium plants ?[J]. Plant and Soil, 2006, 279: 51-63.
Porras-Alfaro A, Herrera J, Sinsabaugh R L, et al. Novel root fungal consortium associated with a dominant desert grass[J]. Applied and Environmental Microbiology, 2008, 74(9): 2805-2813.
Redman R S, Sheehan K B, Stout R G, et al. Thermo- tolarance generated by plant/fungal symbiosis[J]. Science, 2002, 298(5598): 1581.
Chang W S, van de Mortel M, Nielsen L, et al. Alginate production by Pseudomonas putida creates a hydrated microenvironment and contributes to biofilm architecture and stress tolerance under water-limiting conditions[J]. Journal of Bacteriology, 2007, 189(22): 8290-8299.
猜你喜欢
伴侣(2022年5期)2022-06-16
湖北农业科学(2019年22期)2019-12-23
农家致富顾问·下半月(2019年7期)2019-09-28
中国中药杂志(2016年24期)2017-04-18
中国中药杂志(2017年4期)2017-03-28
中国中药杂志(2017年3期)2017-03-20
科技创新导报(2016年28期)2017-03-14
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
山东农业科学(2016年11期)2016-12-17
