
绵羊Izumo1基因多态性及其与产羔数关联分析
2020-03-17 09:19胡文萍董新龙田志龙汤继顺刘秋月王翔宇张效生张金龙王金玉储明星
中国农业大学学报 2020年1期
胡文萍 董新龙,2 田志龙 汤继顺,3 刘秋月 王翔宇 狄 冉 张效生 张金龙 王金玉 储明星*
(1.中国农业科学院 北京畜牧兽医研究所/农业农村部动物遗传育种与繁殖重点实验室,北京 100193; 2.扬州大学 动物科学与技术学院,江苏 扬州 225009; 3.安徽省农业科学院 畜牧兽医研究所,合肥 230031; 4.天津市畜牧兽医研究所,天津 300381)
动物受精的成败决定种群能否在自然界稳定存在。哺乳动物受精过程中主要经历了精子获能、穿越放射冠、识别卵子透明带、发生顶体反应、精卵粘附与融合、卵母细胞第二极体排出、两性原核的融合等步骤[1-2]。早期研究表明致育蛋白(Fertilin)存在于精子质膜上,是一种在精子-卵质膜粘附中起作用的精子表面蛋白,并可能参与受精卵融合[3-6]。致育蛋白由Fertilin α和Fertilin β 2 个亚基组成:敲除Fertilin α基因的雄性小鼠仍能生育[7];敲除Fertilinβ基因小鼠表现出雄性不育,主要原因不是质膜融合失败,而是精子无法与透明带粘附且从子宫向输卵管的迁移发生障碍[8]。
Inoue等[9]发现精子表面蛋白Izumo1 (Izumo sperm-egg fusion 1, Izumo1)与精卵结合相关。已有研究发现:卵子表面的Juno蛋白是Izumo1在卵子表面的受体[10];Izumo1和Juno的相互作用是精卵质膜融合所必须,Izumo1和Juno也是目前发现的精子和卵子质膜上的第一个配体-受体蛋白对。Izumo 是免疫球蛋白超家族(Immunoglobulin superfamily, IgSF)的成员[11-12]。而Izumo1是Izumo基因家族的成员之一,Izumo基因家族有4 个成员,分别为Izumo1、Izumo2、Izumo3和Izumo4,拥有同源的N端结构域,“Izumo结构域”。Izumo1~3是跨膜蛋白,均在睾丸上特异性表达。Izumo1也在人[14]、小鼠[9]、猪[1]、绵羊[15]等哺乳动物受精过程中表达。Izumo4是可溶性蛋白,在包括睾丸的多种组织中表达[9,13]。Izumo1的细胞外结构域中有1 个免疫球蛋白(Immunoglobulin, Ig)结构域,并在成熟精子的顶层上表达,顶体反应之后,Ig结构域在精子表面暴露。Izumo1的Ig结构域上存在1 个潜在的糖基化位点,在C-末端还有1 个跨膜结构域和1 个短的胞质内尾巴,属I型膜蛋白。
NCBI上公布的特克赛尔羊Izumo1基因位于第14 号染色体上,其DNA全长3 326 bp,拥有8 个外显子和7 个内含子,编码320 个氨基酸。Izumo1基因编码的蛋白质位于精子质膜内部,在精子上特异性表达[14]。在顶体反应发生时,Izumo1蛋白会从精子头部前端重新定位到发生融合的位点,包括整个精子头部和精子赤道部位,使得在顶体反应后暴露在精子头部的膜外。通过Izumo1的抗体抑制试验能够明显地抑制精卵间的结合和融合过程[1,9,11],将Izumo肽段直接注射到体内可以造成雌性小鼠免疫性不孕[16]。Izumo基因双敲除的雌性小鼠具有正常生育能力,但是雄性小鼠虽然能够正常生长、产生精子和进行交配,却无法产生后代。通过体外受精试验发现,Izumo1-/-雄性小鼠的精子虽然可以与去透明带卵母细胞正常结合,但不能继续融合而是积留在质膜外侧的卵周隙中无法完成受精过程。将Izumo缺陷型精子注射到野生型卵母细胞中,能正常的产生子代,且其子代拥有正常的生育能力。人类精子也含有Izumo,添加抗人类Izumo的多克隆抗体能使精子无法与去透明带的仓鼠卵融合[9]。这些结果均表明Izumo1参与介导并影响精卵融合过程。
目前,对于Izumo1基因的研究多在哺乳动物精卵融合方面。牛[17]、猪[1]、内蒙古白绒山羊[14,18]、绵羊(品种未知)[14,18]和斑马鱼[19]的Izumo1基因被成功克隆。内蒙古白绒山羊、绵羊(品种未知)2者的Izumo1基因同源性高达99.9%,并与牛、鼠和人Izumo1基因高度同源[18]。然而,Izumo1基因在各个物种中的多态性以及与产生子代数目的相关性研究,特别是有关多羔和常年发情的高繁殖力绵羊小尾寒羊[20-21]Izumo1基因的克隆,多态性检测及与产羔数的关联研究尚未见报道。本课题组前期对来自10 个绵羊品种的99 个个体进行了全基因组重测序,获得了大量的基因多态性数据。为研究Izumo1基因的多态性及与产羔数的关系,本研究拟利用PCR和直接测序法对小尾寒羊和苏尼特羊Izumo1基因DNA进行扩增和测序,并与本课题组前期绵羊重测序数据比对,在多羔和单羔绵羊群体中筛选出SNP位点,并采用Sequenom MassARRAY®技术进行Izumo1基因分型,研究其SNP位点的各种基因型在各群体中的分布,以期获得与繁殖相关的候选SNPs位点,为绵羊遗传育种研究提供新的分子标记。
1 材料与方法
1.1 材料及试剂
选择760 只年龄在3 岁左右的经产绵羊,包括:有产羔数记录的多羔品种小尾寒羊380 只;5 个单羔品种共计380 只(单羔品种分别是苏尼特羊100 只,滩羊80 只,萨福克羊39 只,杜泊羊30 只,草原型藏羊131 只)。其中:小尾寒羊来自山东省郓城县诚联小尾寒羊种羊场和山东省章丘市晟益牧业有限公司,苏尼特羊来自内蒙古巴彦淖尔市乌拉特中旗民羊牧业有限责任公司,滩羊来自宁夏盐池县宁夏朔牧盐池滩羊繁育有限公司,萨福克、杜泊羊来自北京市顺义区北京奥鑫牧业有限公司;草原型藏羊来自西藏当雄县。对所有绵羊进行颈静脉采血,每只绵羊分别采血10 mL,经EDTA抗凝处理后4 ℃保存备用。
2×TaqPCR Master Mix(MT201-01)购自北京博迈德;TaKaRa LATaq®with GC Buffer (RR02AG)、TaKaRa MiniBEST Agarose Gel DNA Extraction Kit Ver.4.0 (9762)、E.coliDH5α Competent Cells (9057)、pMDTMe18-T Vector Cloning Kit (6011)均购自宝生物工程(大连)有限公司。
1.2 DNA提取及检测
参照血液基因组DNA提取试剂盒(天根生化科技有限公司,北京)说明书提取DNA。用Nanodrop 2000检测DNA的纯度和浓度,1.5% 的琼脂糖凝胶电泳检测其完整性。
1.3 引物设计与合成
根据绵羊Izumo1基因全基因组序列(GenBank登录号为:NC_019471.2),采用Primer Premier 5.0软件设计Izumo1基因DNA序列扩增引物。Izumo1基因DNA序列较长,为了方便测序和提高测序准确度,故分段设计4 对扩增引物,分别是P1、P2、P3和P4。引物由北京天一辉远生物科技有限公司合成。引物序列、扩增片段大小及退火温度见表1。
1.4 DNA序列扩增
以提取的血液基因组DNA为模板,用上、下游分段引物扩增目的基因DNA序列。
使用TaKaRa公司高保真LATaq酶体系进行PCR扩增,具体反应体系如下:0.25 μL TaKaRa LATaq(5 U/μL),10 μL 2× GC Buffer I (5 mmol/L Mg2+Plus),3 μL dNTP Mixture (各2.5 mmol/L),4.25 μL RNase-free ddH20,0.5 μL上游引物,0.5 μL下游引物,1.5 μL血液DNA模板,总计20 μL反应体系。
1.4.2PCR反应程序
Izumo1基因:95 ℃预变性5 min;95 ℃变性30 s,Ta退火温度退火1 min,72 ℃延伸2 min,35 个循环;72 ℃延伸8 min。反应结束后,PCR产物进行1.5%琼脂糖凝胶电泳检测所得片段大小,将阳性扩增产物送上海生工生物工程有限公司克隆测序。
1.5 SNP位点筛选
DNAMAN软件进行多重比对和分析目的基因DNA序列,并结合Chromas软件中的峰图判断和本课题组前期重测序数据,以同一位点不同碱基出现比例>30%认定为SNP位点。初步筛选得到SNP位点,鉴定同义突变和非同义突变。

表1 绵羊Izumo1基因DNA扩增引物Table 1 Primers used for amplifying DNA of Izumo1 gene in sheep
1.6 基因分型
对筛选获得的目的基因SNP位点,通过Sequenom MassARRAY®(Sequenom iPLEXTMassay, San Diego, CA)基因分型系统对小尾寒羊、苏尼特羊、滩羊、萨福克羊、杜泊羊和草原型藏羊血液DNA样品进行SNP分型,试验步骤按照仪器操作指南进行。试验由北京君诺德生物技术有限公司完成。
1.7 统计分析
用Popgene 32(version 3.2)软件和PIC-CALC程序对获得的SNP位点分型结果进行处理,计算其杂合度He(Heterozygosity)、多态信息含量PIC(Polymorphism information content)等。采用SPSS 18.0软件卡方检验对2 个品种的各SNPs位点基因型频率和基因频率进行差异显著性检验,并与产羔数进行相关性分析。相关性分析模型为:
yijkl=μ+Genotypei+Pj+Sk+eijkl
式中:yijkl为性状观察值;μ为总体均数;Genotypei为基因型效应;Pj为胎次效应;Sk为场次效应;eijkl为随机误差。假定eijkl相互独立,服从N(0,σ2)分布。
2 结果与分析
2.1 血液DNA检测结果
利用NanoDrop 2000超微量分光光度计和琼脂糖凝胶电泳检测小尾寒羊和苏尼特羊组织DNA的纯度、浓度和完整性。检测结果发现,D260nm/D280nm比值都在1.8~2.0范围内,浓度均在30 ng/μL 以上,电泳条带合格,可用于后续试验。
2.2 DNA序列扩增结果
凝胶电泳结果显示PCR扩增的DNA片段条带明亮且单一,大小与设计一致,说明扩增结果良好(图1)。使用DNAMAN软件对测序得到的DNA片段进行序列拼接,成功得到2 种绵羊Izumo1基因完整DNA序列。特克赛尔羊Izumo1基因DNA全长序列为3 326 bp,本试验扩增得到的小尾寒羊Izumo1基因DNA全长序列3 385 bp,苏尼特羊Izumo1基因DNA全长序列3 382 bp。BLAST结果显示二者与特克赛尔羊Izumo1基因DNA序列的相似性都高于95%。与NCBI数据库中提供的特克塞尔羊Izumo1基因DNA序列相比:在Izumo1基因DNA序列605 bp位置,小尾寒羊和苏尼特羊分别多出了57和55 bp的序列(57 bp: ACCCCCCACCCCCCCCGCGCAGATGAGGCCA-CACTGGAAAAGGCATCCTGGAGTTTG;55 bp: ACCCCCCACCCCCCGCGCAGATGAGGCCACA-CTGGAAAAGGCATCCTGGAGTTTG)。
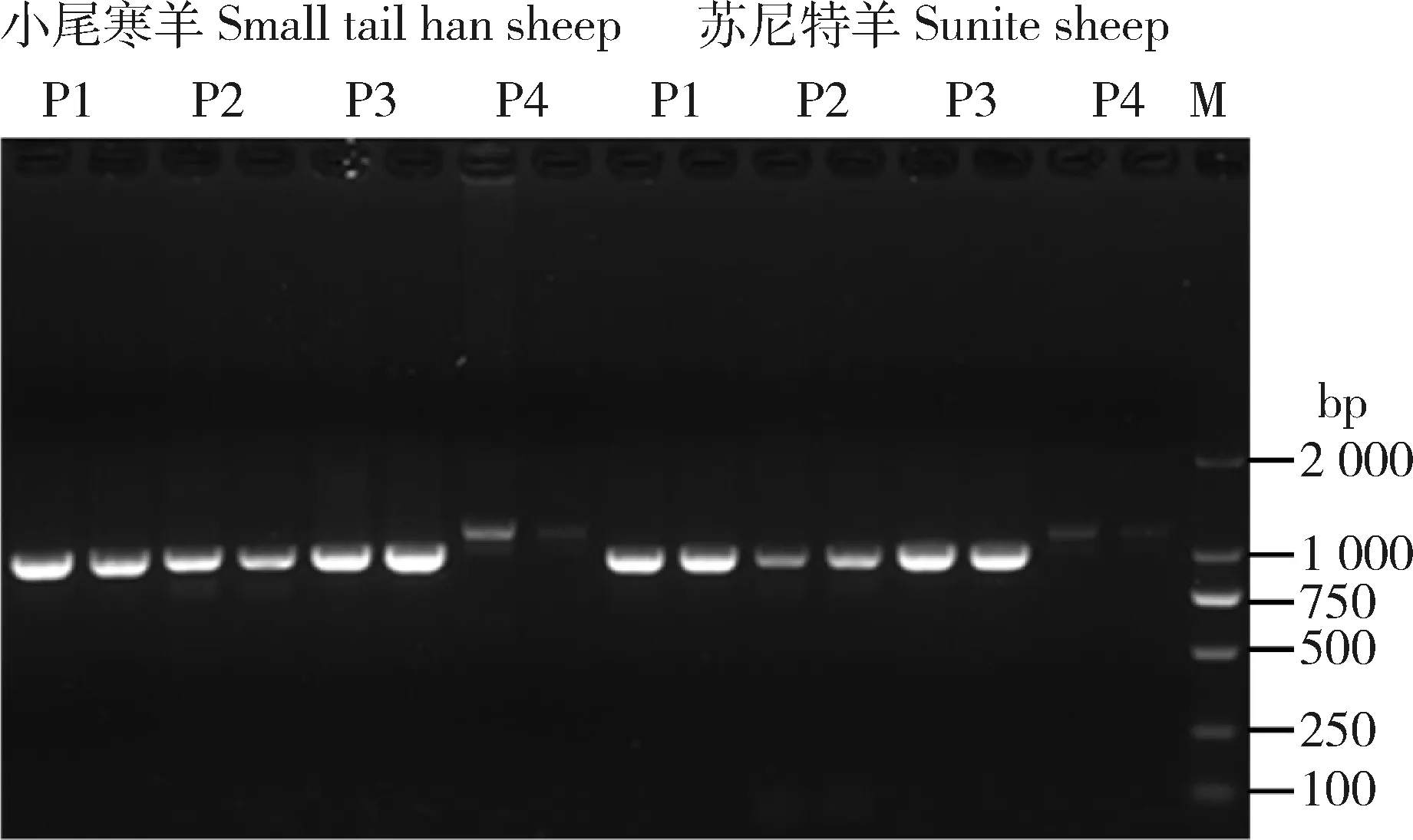
M:DL2 000 DNA分子量标记; P1~ P4:绵羊Izumo1基因DNA扩增引物。
M, DL2 000 DNA marker; P1-P4, primers used for amplifying DNA ofIzumo1gene in sheep.
图1Izumo1基因序列分段扩增产物电泳图
Fig.1 Electrophoresis of segmental amplification products ofIzumo1gene
2.3 SNP位点筛选结果分析
用4 对引物分别对小尾寒羊和苏尼特羊的60 个样品进行目的基因序列的SNP位点筛选,对PCR扩增特异性好的单一条带进行双向测序,测序峰图清晰可辨的位点用于SNP统计共获得了5 个SNP位点(g.54409483A>G,g.54410565A>G,g.54411606C>A,g.54411750T>C,g.54411792A>G)(图2)。另外结合本课题组前期对10 个绵羊品种的99 个个体进行的全基因组重测序数据,筛选得到Izumo1基因的另外3 个SNP突变位点(g.54409033T>A, g.54412107C>A,g.54412135A>G)。共筛选得到8 个潜在的SNPs位点, 6 个位于外显子区域,均为错义突变,2 个位于5’UTR。其中A/G转换的数量最多,共有4 个;T/C 转换的数量为1 个;C/A颠换2 个;T/A颠换1 个;不存在G/C颠换和G/T颠换。转换的数量占总检测到的SNPs位点的62.5%,颠换占37.5%。

图2 通过扩增获得的5个Izumo1基因SNP位点
Fig.2 Five SNPs ofIzumo1gene obtained by amplification
2.4 Izumo1基因SNP位点与绵羊产羔性状关联分析
根据分型所获得的数据,对上述8 个位点进行了初步筛选,将在单、多羔绵羊群体中位点分布没有差异的SNP位点排除,最终筛选获得的g.54412135A>G和g.54412107C>A 2个位点。通过基因分型发现:Izumo1基因g.54412135A>G位点在多羔和单羔绵羊品种中存在GG、GA和AA 3种基因型,g.54412107C>A在多羔品种中存在CC和CA 2 种基因型,而在单羔品种中存在CC、CA和AA 3种基因型(表2)。g.54412135A>G位点在小尾寒羊、滩羊和草原型藏羊品种中存在GG、GA和AA 3种基因型,而在苏尼特羊中存在GG和GA 2种基因型,在萨福克羊和杜泊羊中仅有GG 1种基因型;g.54412107C>A在小尾寒羊、苏尼特羊、萨福克羊和草原型藏羊品种中存在CC和CA 2种基因型,而在滩羊和杜泊羊品种中存在CC、CA和AA 3种基因型(表3)。Izumo1基因的g.54412107C>A位点的基因型频率和基因频率在单、多羔绵羊群体间的分布差异均极显著(P<0.01)。
利用Popgen 32软件统计分析绵羊Izumo1基因2 个品种各SNP位点的多态性。由表3可知,在6 个品种中的g.54412135A>G多态位点多态信息含量(PIC)都属于低度多态(PIC<0.25);杜泊羊的g.54412107C>A多态位点属于中度多态(0.25
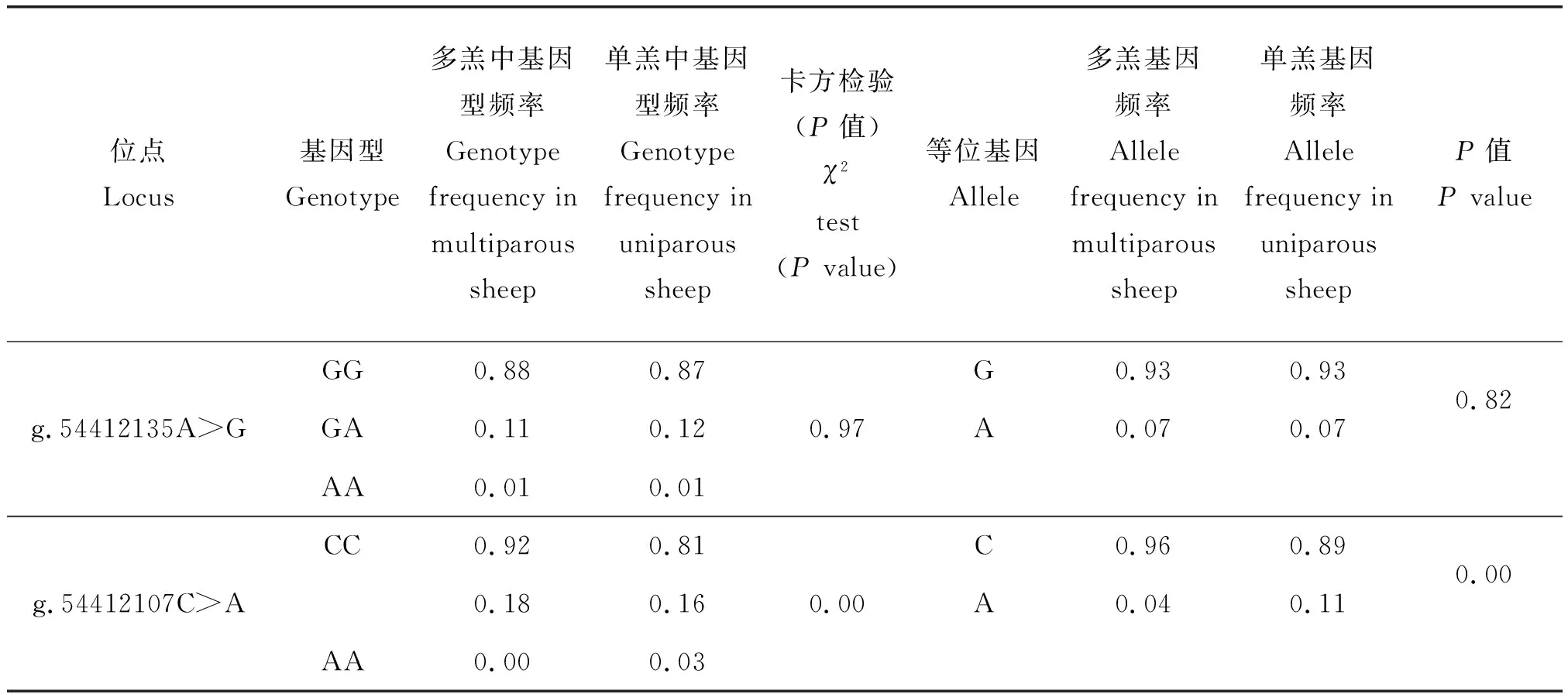
表2 在单、多羔绵羊品种Izumo1基因2 个位点的基因型频率和等位基因频率分析Table 2 Genotype and allele frequencies of two SNPs of Izumo1 gene in monotocous and polytocous sheep
注:P<0.01表示差异极显著。
Note:P<0.01 indicates extremely significant difference.
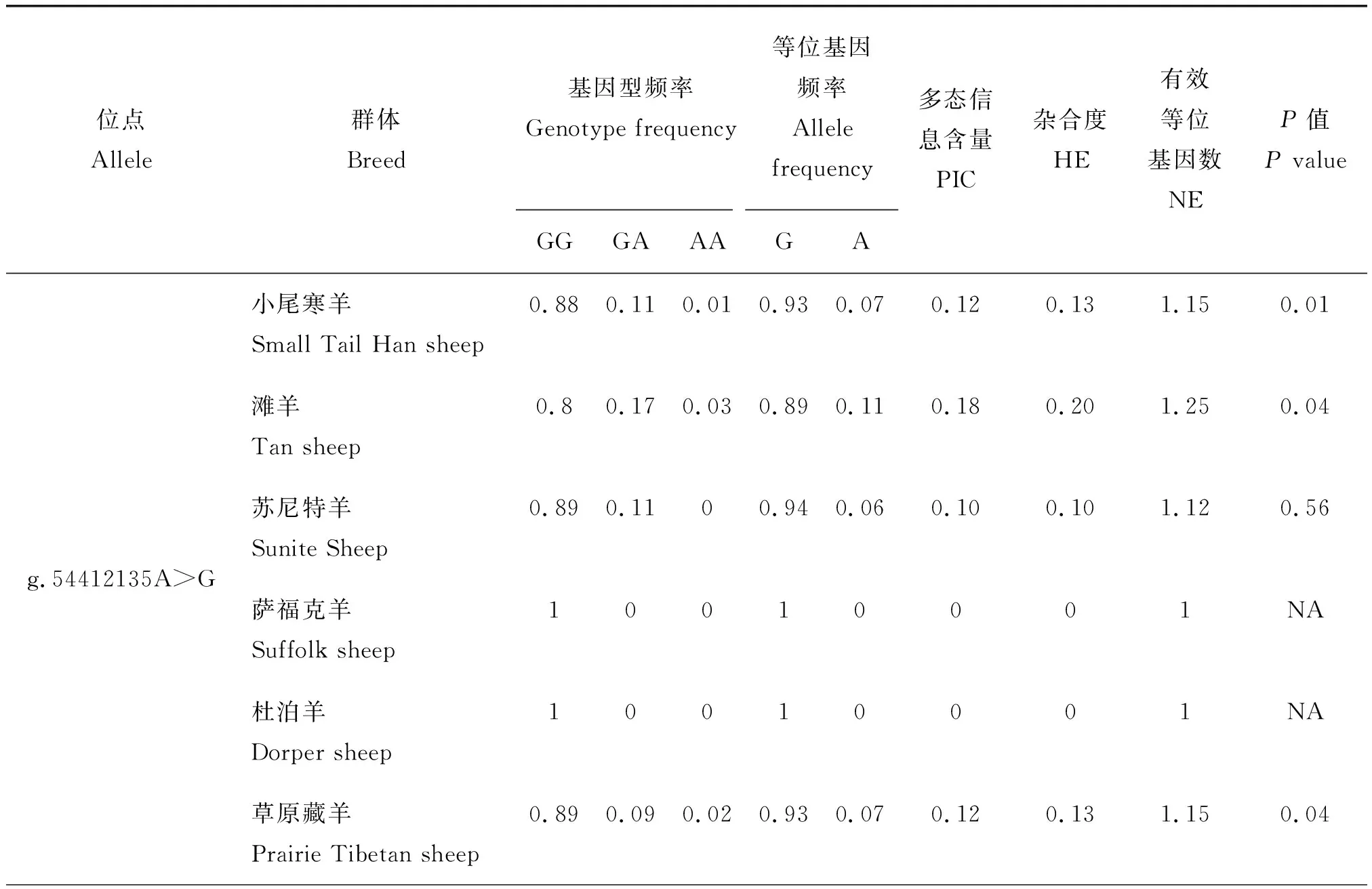
表3 6个绵羊品种中Izumo1基因的2个SNPs位点的群体遗传学分析Table 3 Genetic polymorphism information of Izumo1 gene two SNPs in six sheep breeds

表3(续)
注:P>0.05表示位点在该品种中处于哈代温伯格平衡状态;P<0.05表示位点在该品种中不处于哈代温伯格平衡状态。
Note:P>0.05 indicates the locus was under Hardy-Weinberg equilibrium.P<0.05 indicates the locus was not under Hardy-Weinberg equilibrium.
Izumo1基因g.54412135A>G和g.54412107C>A位点的不同基因型与小尾寒羊产羔数的关联分析统计结果。可看出2 个位点不同基因型与小尾寒羊不同胎次产羔数之间,不存在显著关联(P>0.05)(表4)。
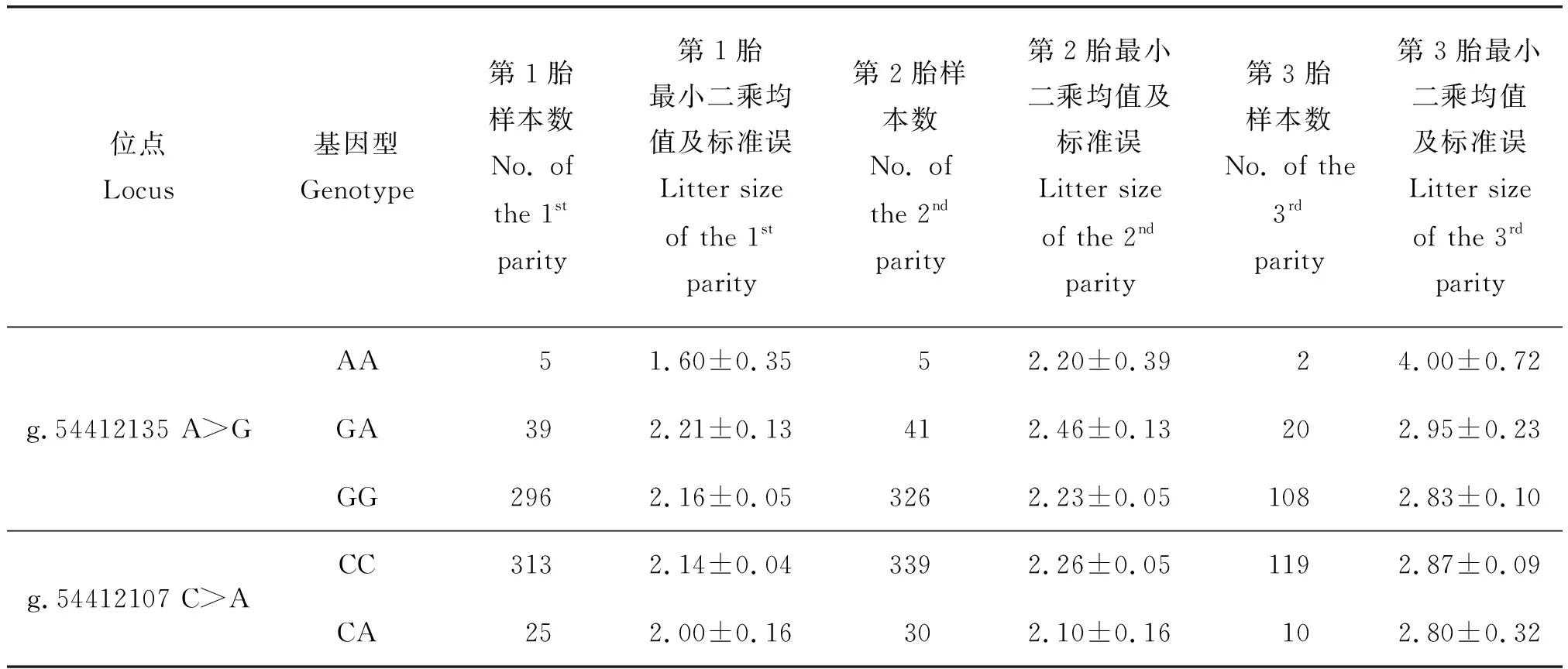
表4 Izumo1基因2 个多态位点的不同基因型与小尾寒羊产羔数的关联分析Table 4 Association of differrent SNP genotypes of the Izumo1 gene with litter size in Small Tail Han sheep
3 讨论与结论
基因序列的外显子区域是相对保守的,因此在外显子区SNP位点比较少,因此,外显子出现变异的频率仅为内含子等区域的1/5[22]。外显子区域中发生变异的影响远>内含子区,因为外显子的变异可能会造成氨基酸的改变,从而改变蛋白质的性质与功能,然而,真核生物的基因表达调控非常复杂,是基因与多种转录因子共同参与、相互作用的结果。基因的非翻译区(UTR)常常参与基因的表达调控。5′UTR常常可以决定转录的水平和方式,而 3′UTR 可以调控mRNA的稳定性[23-25]。因此,外显子区和5′UTR的基因变异常常会直接影响生物性状,在遗传育种的研究中意义重大。
自2005 年Inoue等[9]发现Izumo1与精卵结合相关后,其功能和作用机制的研究在哺乳动物中展开,虽有报道陆续克隆了牛[17]、猪[1]、山羊[14,18]、绵羊[14,18]和斑马鱼[19]的Izumo1基因,而对于其单核苷酸多态性关注度并不高,Izumo1基因在各个物种中的多态性与产生子代数目的相关性的研究还未见报道。目前在Ensemble数据库(http:∥asia.ensembl.org/Ovis_aries/Transcript/ProtVariations?db=core;g=ENSOARG00000012130;)中发布的绵羊(Ovisaries)Izumo1基因的SNPs位点只有7 条记录。本研究在Izumo1基因DNA序列中筛选到8 个SNPs位点,丰富了绵羊Izumo1基因的单核苷酸多态性。其中,在外显子区筛选到6 个SNPs位点并均为错义突变,遗憾的是无法在6 个绵羊品种中进行基因分型,只有2 个处于5′UTR中的位点分型成功。
产羔数是绵羊重要的繁殖性状,已有研究对其研究主要聚焦在母羊排卵方面,发现了诸如生长分化因子9(GDF9)、骨形态发生蛋白15(BMP15)和骨形态发生蛋白受体IB(BMPR1B)等高繁殖力主效基因[26-30],尤其是显著影响小尾寒羊排卵数的BMPR1B基因FecB突变[31]。精子Izumo1蛋白,作为一种跨膜蛋白,主要对哺乳动物受精过程中的精卵融合具有重要作用[9],能够通过与卵子的识别和融合影响受精过程,猜测其可能对于绵羊繁殖也至关重要[14]。Inoue等[9]利用重组Izumo蛋白制备的多克隆抗体进行免疫标识,发现小鼠Izumo蛋白大小为56.4 ku,仅在睾丸组织中表达。Kim等[1]研究证实Izumo1蛋白位于精子质膜内部,在精子上特异性表达,只在精卵融合时才会在精子头部检测到。本研究只对小尾寒羊母羊进行了产羔数的关联分析,发现Izumo1基因的2 个多态性位点与小尾寒羊母羊不同胎次产羔数之间不存在显著关联。推测Izumo1基因可能在母羊个体本身并没有重要的生物学作用,与母羊的产羔数性状并不相关。在其他物种中,Izumo1是否与产生子代数量相关有待进一步研究。Izumo1作为精卵质膜融合的关键性配体蛋白,直接决定了精子是否能够正常与卵子发生融合,推测Izumo1基因可能与公羊的繁殖力性状有关。本研究克隆了完整的小尾寒羊和苏尼特羊的Izumo1基因,并在Izumo1基因中获得了8 个SNPs位点。其中:Izumo1基因的g.54412107C>A位点基因型频率和等位基因频率在单、多羔绵羊群体间的分布差异均极显著。Izumo1基因g.54412135A>G和g.54412107C>A位点的不同基因型和小尾寒羊不同胎次的产羔数之间没有显著关联(P>0.05)。
猜你喜欢
畜牧与饲料科学(2022年6期)2022-12-19
畜牧与饲料科学(2022年6期)2022-12-19
云南畜牧兽医(2020年6期)2020-12-18
新农业(2017年1期)2017-03-16
兽医导刊(2016年6期)2016-05-17
兽医导刊(2016年6期)2016-05-17
中国质量与标准导报(2014年10期)2014-02-28
新疆农垦科技(2014年10期)2014-02-28
当代畜禽养殖业(2014年4期)2014-02-27
山东畜牧兽医(2010年3期)2010-08-15
