
注意线索对自闭特质个体疼痛共情的影响:来自事件相关电位的证据*
2020-03-16 05:23:56李祚山向滨洋
心理学报 2020年3期
李 雄 李祚山 向滨洋 孟 景
(1重庆市人文社科重点研究基地课程与教学研究基地; 2重庆师范大学应用心理学重点实验室, 重庆 401331)
1 前言
共情(empathy)是人类作为“社会性动物”的重要基础, 使我们能够理解他人的心理和情感状态,能帮助我们有效地与他人沟通, 预测他人的行为、意图和感受(Singer & Lamm, 2009)。疼痛共情(empathy for pain)是共情典型的表现形式, 指个体对他人疼痛的感知、判断和情绪反应(Meng et al.,2012)。以往研究发现个体对疼痛线索自上而下的注意会调节其疼痛共情的加工机制, 与不注意图片中的疼痛线索相比, 当被试注意疼痛线索时, 与自身疼痛相关的脑区激活水平上升, 如扣带前回和右侧额中回等区域(Gu & Han, 2007), 事件相关电位(event related potentials, ERPs)的晚期认知评估阶段P3和LPC的波幅会增大(Fan & Han, 2008)。
自闭症谱系障碍(autistic spectrum disorder,ASD)是一种发育性障碍, ASD个体通常会在许多环境中表现出社会沟通与社会交往的持续性缺陷,并存在某些重复刻板的行为模式和狭窄的兴趣及活动(American Psychiatric Association, 2013)。Baron-Cohen认为ASD个体社会沟通与社会交往缺陷等社会性特征是他们共情能力的缺损导致的(Baron-Cohen, 2010)。因此, 共情障碍可能是ASD个体的核心症状之一(Adler, Dvash, & Shamay-Tsoory, 2015; Lawson, Baron-Cohen, & Wheelwright,2004)。虽然有研究结果支持了这一理论(Baron-Cohen & Wheelwright, 2004), 但是也有研究发现ASD个体无论是在识别他人的情绪(Evers, Steyaert,Noens, & Wagemans, 2015; Jones et al., 2011), 还是在更为复杂的情景中都没有表现出严重的共情缺陷(Scambler, Hepburn, Rutherford, Wehner, &Rogers, 2007), 这可能是由于在日常生活中ASD个体难以主动注意并加工复杂情境中的情绪信息(Begeer, Koot, Rieffe, Terwogt, & Stegge, 2008)。有实验室研究也发现, 当要求 ASD个体注意他人的面孔和声音中的情绪线索时, 他们与普通个体一样都出现内侧前额叶皮质激活水平上升, 而不要求注意情绪线索时则没有显著变化(Wang, Lee, Sigman,& Dapretto, 2007)。说明了对情绪线索自上而下的注意可能对 ASD个体的共情有着重要影响, 这一结论也得到了许多研究结果的支持(Fan, Chen,Chen, Decety, & Cheng, 2014; Hagenmuller, Rössler,Wittwer, & Haker, 2014)。
ASD的症状并不是非有即无的, 它在普通人群中是从正常到自闭的连续性分布(Hoekstra et al.,2010), 可以用自闭商数问卷(Autism-Spectrum Quotient, AQ) (Baron-Cohen, Wheelwright, Skinner,Martin, & Clubley, 2001)进行测量。通常, ASD个体的 AQ 分数分布在最高分一端(Gökçen, Petrides,Hudry, Frederickson, & Smillie, 2014), 而AQ分数较高但行为表现没有达到 ASD临床诊断标准的普通人被称为自闭特质(autistic traits)者。有研究证实了自闭特质个体与 ASD个体有相似的行为模式(Reed, Lowe, & Everett, 2011), 共情方面的研究发现普通个体的自闭特质分数越高, 其面部表情加工能力越低(Luo, Burns, & Xu, 2017), 在辨别视觉与听觉上呈现的疼痛刺激时, 自闭特质组相比于控制组正确率更低(Meng, Li, & Shen, 2017)。
最近的一项 ERP研究发现对疼痛线索自上而下的注意也会调节自闭特质个体的听觉疼痛共情(Meng, Shen, Li, & Peng, 2019b)。该研究通过视觉和听觉两个实验来探讨对疼痛线索自上而下的注意对疼痛共情的调节作用, 每个实验包括两个任务:1)疼痛判断任务, 要求被试判断手脚图片(视觉实验)或声音(听觉实验)中的人物是否疼痛, 该任务让被试注意疼痛线索; 2)数量判断任务(视觉实验)或性别判断任务(听觉实验), 要求被试判断图片中手的数量或声音的性别, 该任务让被试不注意疼痛线索。ERP结果发现在听觉模态下, 不注意疼痛线索时, 自闭特质个体比控制组在N1和P2上的平均波幅更小, 说明了当自闭特质个体不去主动地注意他人的疼痛时, 其疼痛共情能力减弱。然而, 该研究没有发现自闭特质个体在视觉模态下与控制组存在差异。这可能与视觉实验中采用手脚受伤的图片而没有采用面孔图片作为刺激材料有关。
ASD个体和自闭特质个体可能存在异常的面孔加工。有研究让 ASD个体观察正立和倒立的面孔, 发现 N170成分的潜伏期比普通个体更长; 普通个体加工正立的面孔时潜伏期比倒立的面孔更短, 但ASD个体没有显著差异(Khorrami, Tehrani-Doost, & Esteky, 2013), 说明ASD个体对于面孔信息的加工存在一定困难。这可能是由于 ASD个体对社交信息缺乏兴趣, 回避眼部、嘴部这类社会信息丰富的区域(Chita-Tegmark, 2016; Key & Stone,2012; Ma & Zhu, 2014; Wagner, Hirsch, Vogel-Farley,Redcay, & Nelson, 2013), 因此回避眼睛也是ASD的临床诊断标准之一(Lord et al., 2000)。以往研究还发现普通个体的自闭特质分数与高兴面孔的眼神凝视引起的注意定向呈负相关(Lassalle & Itier,2014), 与右侧后颞上沟区域的激活程度呈正相关,该区域与眼神接触和社会信号加工有关(Hasegawa et al., 2013)。因此本研究以疼痛面孔图片作为刺激材料, 进一步探究自闭特质个体在视觉模态下疼痛共情的认知神经机制。
先前有关疼痛共情的研究认为, 额顶叶 N1、P2、N2成分反映了疼痛共情加工过程中早期的情绪分享和疼痛感知, 顶枕叶P3和LPC则反映了晚期的认知评估(Cui, Ma, & Luo, 2016; Luo, Han, Du,& Han, 2018; Meng et al., 2019b); 而关于面孔的研究认为, 颞枕区 N170是反映面孔加工的特异性成分(Itier & Taylor, 2004)。综上, 本研究以上述ERP成分作为主要指标, 在我们前期研究的基础上, 通过高时间分辨率的 ERP技术, 从疼痛共情加工的不同阶段来探究视觉注意线索对自闭特质个体面孔疼痛共情的影响。
基于以往的研究证据, 本研究对实验结果提出以下预期:当自闭特质个体注意疼痛线索时, 对他人疼痛面孔的共情加工进程与控制组相似; 而当自闭特质个体不注意疼痛线索时, 对他人疼痛面孔的共情加工进程与控制组可能存在差异, 这种差异主要体现在疼痛共情加工的晚期阶段。具体来说, 对于疼痛图片, 进行疼痛判断任务时自闭特质组与控制组在早期成分(N1、P2或 N2)和晚期成分(P3或LPC)均没有显著差异; 而进行吸引力判断任务时,两组在晚期成分(P3或LPC)存在显著差异, 而在早期成分(N1、N2或P2)不存在差异。这表达了对视觉疼痛线索自上而下的注意会调节自闭特质个体对他人疼痛面孔的共情神经反应。
2 方法
2.1 被试
采用自闭商数问卷(Autism-Spectrum Quotient,AQ) (Baron-Cohen et al., 2001)中文成人版(Liu,2008)测查了2502名大学生的自闭特质。根据前人研究, 按照AQ分数前、后10%分为高AQ组和低AQ组(Meng et al., 2017; Meng et al., 2019b), 再从两组中分别随机选取 30名大学生作为被试, 作为自闭特质组和控制组, 具体被试信息见表 1。所有被试均为右利手, 无精神病史, 视力或矫正后视力正常。被试在实验前均被告知了实验程序, 并签署了知情同意书, 在实验后会获得一定报酬。

表1 被试年龄与AQ分数信息表
2.2 刺激材料
实验材料选自前人研究(Hu, Abbasi, Zhang, &Chen, 2018), 为黑底灰度的中国化面孔图片, 无名人面孔, 面孔表情呈中性。用Adobe Photoshop CS6对图片进行统一标准化的处理(如图 1所示), 疼痛图片是在原始面孔图片的左侧脸颊上放置针头, 非疼痛图片是在左侧脸颊上放置棉签。所有图片由40名(女性20名)大学生使用9点Likert量表对图片的疼痛程度(1 = 完全不痛, 4 = 开始疼痛, 9 = 疼痛极限)、吸引力程度(1 = 非常不好看, 5 = 中等, 9 =非常好看)、情绪效价(1 = 非常不高兴, 5 = 中等, 9= 非常高兴)和情绪唤醒度(1 = 非常不兴奋, 5 =中等, 9 = 非常兴奋)进行评估, 最终筛选出分辨率、亮度和对比度一致, 图片尺寸均为 260 × 300像素的疼痛图片和非疼痛图片各30张, 一共60张(男女各半)。刺激材料评估数据见表2。
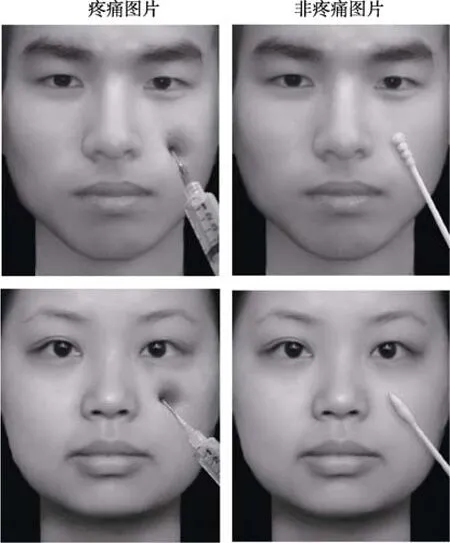
图 1 实验所用材料图片示例(左列为疼痛图片, 右列为非疼痛图片)
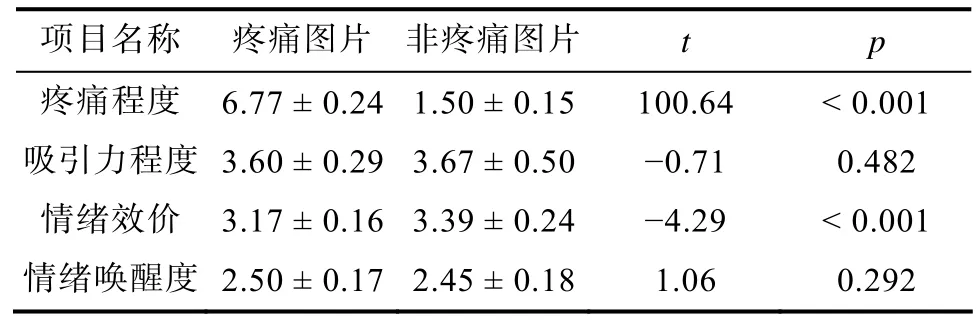
表2 刺激材料评估数据(M ± SD)
2.3 实验设计
本实验为2(组别:自闭特质组, 控制组) × 2(图片类型:疼痛图片, 非疼痛图片) × 2(任务类型:疼痛判断任务, 吸引力判断任务)三因素混合设计。其中组别是被试间变量, 图片类型和任务类型是被试内变量。实验共分为两个任务:任务一为疼痛判断任务, 要求被试判断图片中的人物是否疼痛; 任务二为吸引力判断任务, 要求被试判断图片中的人物是否具有吸引力。最后是主观等级评分, 要求被试按照9点Likert量表对全部60张图片的疼痛程度、吸引力程度和自身对图片的情绪反应进行评分。
2.4 实验程序
实验在一间温度适宜, 安静隔音的实验室里进行, 被试坐在一台24英寸的彩色液晶显示屏前, 被试距离屏幕约80 cm, 图片呈现在黑色背景的屏幕中央。图片呈现时的尺寸为13.5 cm × 11.5 cm (宽×高), 视角约为 9.6×8.2。实验程序由 E-prime 3.0编制。
在疼痛判断任务中, 被试需要判断图片中的人物是否疼痛, 每个trial中首先呈现一个200 ms的注视点“+”, 再随机呈现一个时间为 800~1500 ms的黑屏, 紧接着随机呈现一张图片, 当图片出现时要求被试又快又准确地按1、2键(按键进行被试间平衡)判断图片中的人物是否疼痛, 图片按键后消失, 若被试未作按键反应, 3000 ms后自动消失(如图2上行所示), 两个trial间有500 ms的时间间隔。4种类型图片每张重复呈现5次, 采用伪随机排列,共有300个trial, 分为3个block呈现, 每个block包含100个trial, 每两个block之间休息3~5分钟。在吸引力判断任务中, 被试需要在图片出现时又快又准确地按1、2键(按键进行被试间平衡)判断图片中的人物是否具有吸引力, 实验流程同疼痛判断任务(如图2下行所示)。对两个任务的呈现顺序进行被试间平衡, 被试进行疼痛判断任务与吸引力判断任务时, 同时记录EEG数据。
EEG记录结束之后进行主观等级评分, 让被试根据自己的主观感受对图片进行评分, 每个 trial中首先呈现一个 200 ms的注视点“+”, 然后随机呈现一张图片, 3000 ms后消失, 最后依次呈现疼痛标尺、吸引力标尺和情绪标尺, 要求被试根据 9点 Likert量表对每张图片中人物的疼痛程度(1 =完全不痛, 4 = 开始疼痛, 9 = 疼痛极限)、吸引力程度(1 = 非常好看, 5 = 中性, 9 = 非常不好看)以及自身对图片的情绪反应(1 = 非常高兴, 5 = 中性, 9= 非常不高兴)进行评分。
2.5 数据采集与分析
采用德国Brain Products公司的ERP记录系统,使用安装在 actiCHamp系统(Brain Vision LLC,Morrisville, NC, US)上的按国际10-20系统扩展的64导电极帽同时记录被试的脑电活动(electroencephalography, EEG), 记录时参考电极位于FCz处, 接地电极位于前额接地点上。EEG活动采用DC采集, 带通范围DC~280 Hz, 连续采样频率为1000 Hz。电极与头皮间的阻抗小于5 kΩ。
EEG数据的预处理和分析采用 MATLAB R2016a (Math Works, USA)和 EEGLAB 13.6.5b 组件(Delorme & Makeig, 2004)。分析时以两侧乳突平均值作为参考, 滤波带通为0.1~40 Hz。同时排除所有电极±80 μV的伪迹信号。分析时程截取图片刺激呈现前200 ms至呈现后1000 ms, 并以图片刺激呈现前200 ms作为基线。对有严重肌电干扰的试次进行排除(排除试次数8.78 ± 15.69), 眼动伪迹通过独立主成分分析(ICA)算法进行校正(Jung et al.,2001)。
本研究分析的 ERP成分包括疼痛刺激诱发的早期成分(N1、P2、N2)和晚期成分(P3和LPC), 以及面孔刺激诱发的特异性成分N170。根据地形图,波形图以及相关文献选取电极点和成分分析的时间窗(Decety, Yang, & Cheng, 2010; Meng et al.,2013; Rossion & Jacques, 2008)。N1和N2成分平均波幅分析的电极点为 Fz、AFz、FCz、F1、F2、AF3、AF4、FC1、FC2, 其中选取的时间窗分别为 N1:115~135 ms, N2: 225~245 ms。P2、P3、LPC 成分平均波幅分析的电极点为Pz、CPz、POz、P1、P2、CP1、CP2、PO3、PO4, 其中选取的时间窗分别为P2: 165~185 ms, P3: 295~315 ms, LPC: 400~800 ms。N170平均波幅分析的电极点为 PO7、PO8、P7、P8, 选取的时间窗为160~180 ms。
数据采用IBM SPSS Statistics 15.0进行统计分析。行为数据以被试的按键反应时(删除三个标准差以外的反应时)和主观等级评分为指标, ERP数据选取各个成分的平均波幅为指标, 对反应时和平均波幅分别进行 2(组别) × 2(图片类型) × 2(任务类型)三因素重复测量方差分析, 对主观等级评分进行2(组别) × 2(图片类型)两因素重复测量方差分析。F值比率的自由度通过 Greenhouse-Geisser检验矫正。
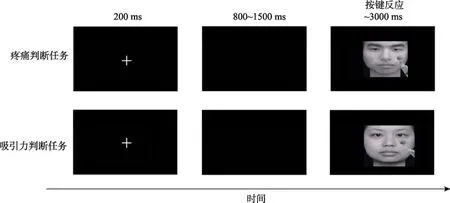
图2 实验流程图(上行为疼痛判断任务实验流程, 下行为吸引力判断任务实验流程)
3 结果
3.1 行为数据
行为数据描述性统计结果见表3。反应时方面,图片类型和任务类型的交互作用显著,F(1, 58) =7.65,p= 0.008,= 0.12。在疼痛判断任务中, 被试对疼痛图片的反应时(787.15 ± 302.65 ms)长于对非疼痛图片的反应时(767.41 ± 287.72 ms),F(1, 58) =4.20,p= 0.045,= 0.07; 在吸引力判断任务中,对疼痛图片的反应时(787.09 ± 139.97 ms)短于对非疼痛图片的反应时(817.29 ± 182.78 ms),F(1, 58) =4.87,p= 0.031,= 0.08。其他主效应和交互作用不显著(所有p> 0.05)。
主观等级评分方面, 在疼痛程度、吸引力程度、自身对图片情绪反应的主观等级评分上, 图片类型主效应显著[疼痛程度:F(1, 58) = 217.20,p< 0.001,= 0.79; 吸引力程度:F(1, 58) = 9.88,p= 0.003,= 0.15; 情绪反应:F(1, 58) = 14.14,p< 0.001,= 0.20]。疼痛图片的疼痛程度评分显著大于非疼痛图片, 吸引力程度评分显著小于非疼痛图片, 情绪反应评分显著大于非疼痛图片。其他主效应和交互作用不显著(所有p> 0.05)。
3.2 电生理证据
ERP数据方差分析结果见表 4, 不同条件下的波形图、地形图以及条形图见图3和图4。
N170 任务类型的主效应显著,F(1, 58) =4.59,p= 0.036,= 0.07, 吸引力判断任务(2.34 ±3.25 μV)的平均波幅显著大于疼痛判断任务(1.97 ±3.37 μV)。组别、图片类型和任务类型的交互作用显著,F(1, 58) = 10.90,p= 0.002,= 0.16。简单效应分析显示, 自闭特质组在疼痛判断任务中疼痛图片(1.55 ± 3.53 μV)的平均波幅显著小于非疼痛图片(1.94 ± 3.40 μV),F(1, 58) = 4.69,p= 0.035,=0.08; 在吸引力判断任务中疼痛图片(2.21 ± 3.24 μV)和非疼痛图片(2.12 ± 3.30 μV)没有显著差异,F(1,58) = 0.43,p= 0.517。控制组在疼痛判断任务[疼痛图片:2.37 ± 3.19 μV, 非疼痛图片:2.02 ± 3.46 μV;F(1, 58) = 3.83,p= 0.055]和吸引力判断任务[疼痛图片:2.39 ± 3.22 μV, 非疼痛图片:2.63 ± 3.38 μV;F(1, 58) = 2.78,p= 0.101]中疼痛图片和非疼痛图片没有显著差异。
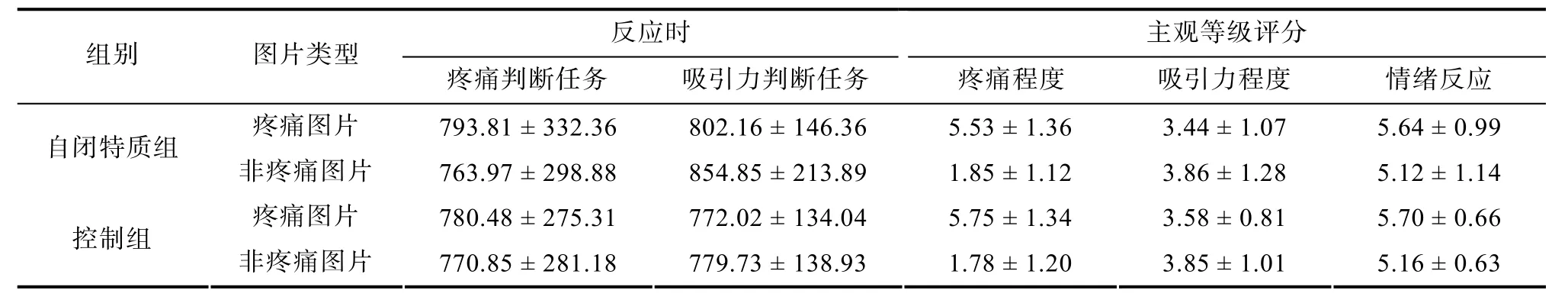
表3 行为数据描述统计(M ± SD)
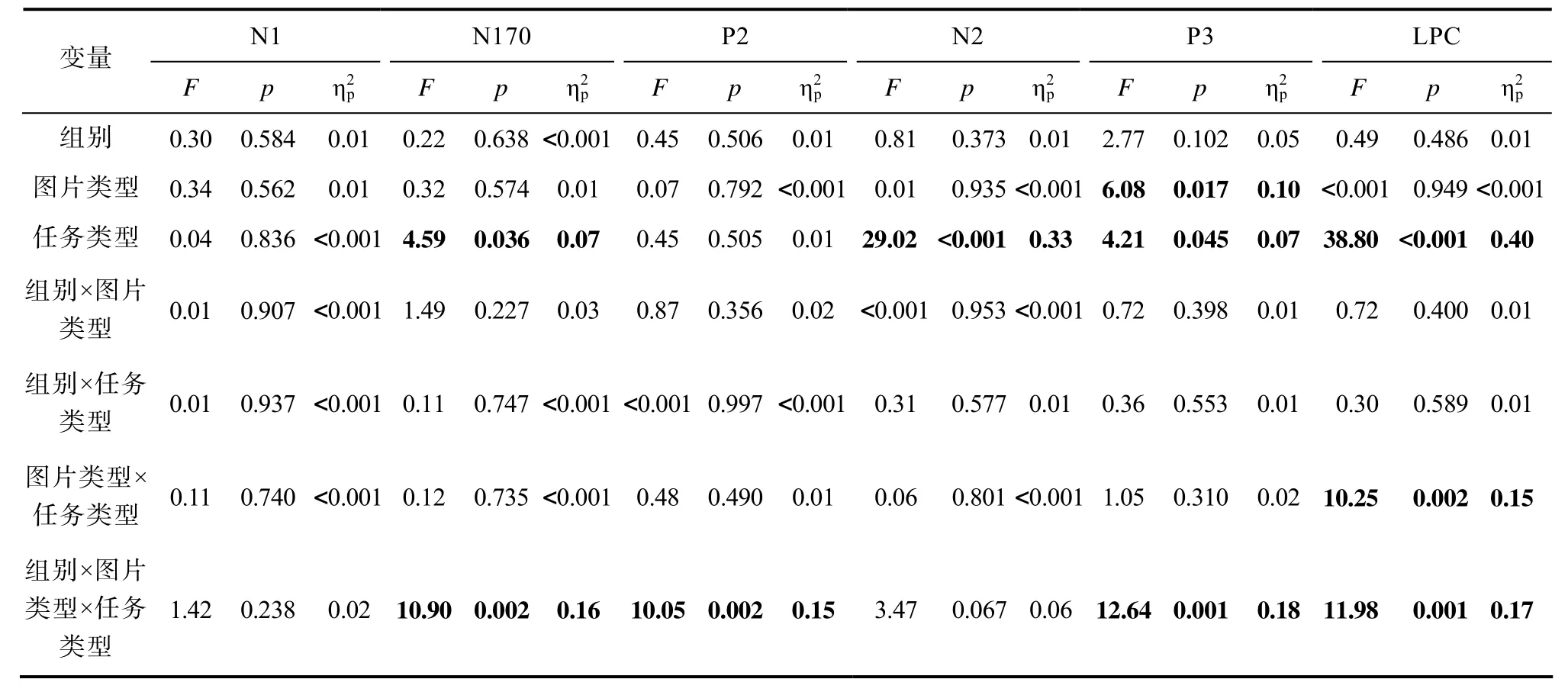
表4 ERP数据方差分析结果
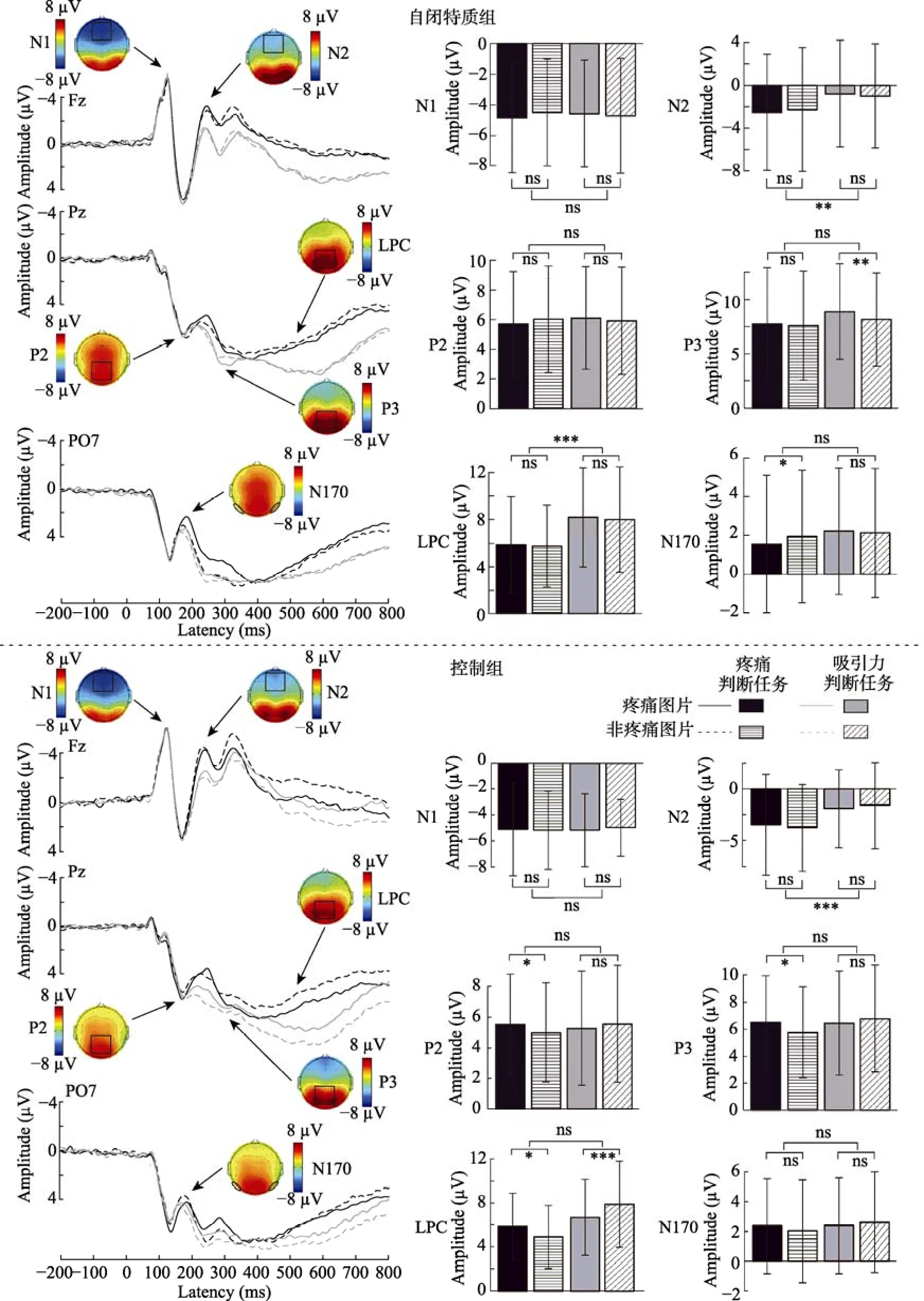
图3 自闭特质组和控制组在不同条件下的波形图、地形图和条形图
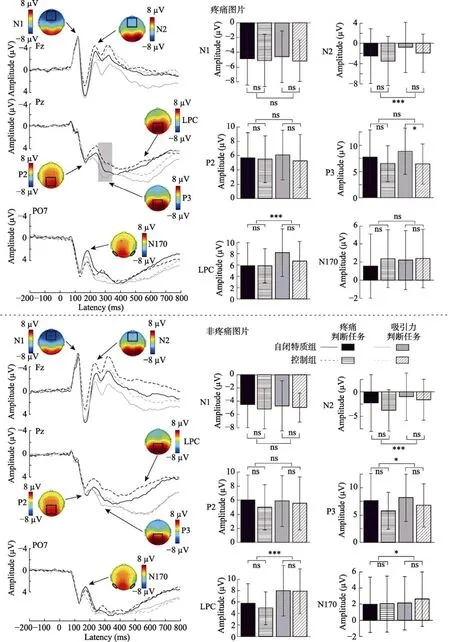
图4 两组被试对疼痛和非疼痛图片的波形图、地形图和条形图
N2 任务类型的主效应显著,F(1, 58) = 29.02,p< 0.001,= 0.33, 疼痛判断任务(−3.01 ± 5.10 μV)的平均波幅显著大于吸引力判断任务(−1.33 ±4.44 μV)。
P2 组别、图片类型和任务类型的交互作用显著,F(1, 58) = 10.05,p= 0.002,= 0.15。简单效应分析显示, 对控制组, 疼痛判断任务中疼痛图片(5.52 ± 3.26 μV)的平均波幅显著大于非疼痛图片(4.99 ± 3.23 μV),F(1, 58) = 5.68,p= 0.020,=0.09; 吸引力判断任务中疼痛图片(5.26 ± 3.71 μV)的平均波幅和非疼痛图片(5.55 ± 3.81 μV)没有显著差异,F(1, 58) = 2.16,p= 0.147。对自闭特质组, 疼痛判断任务[疼痛图片:5.68 ± 3.56 μV, 非疼痛图片:6.01 ± 3.59 μV;F(1, 58) = 2.19,p= 0.144]和吸引力判断任务[疼痛图片:6.09 ± 3.45 μV, 非疼痛图片:5.89 ± 3.61 μV;F(1, 58) = 0.99,p= 0.325]中疼痛图片和非疼痛图片没有显著差异。
P3 图片类型的主效应显著,F(1, 58) = 6.08,p= 0.017,= 0.10, 疼痛图片(7.39 ± 4.31 μV)的平均波幅显著大于非疼痛图片(7.07 ± 4.24 μV)。任务类型的主效应显著,F(1, 58) = 4.21,p= 0.045,=0.07, 疼痛判断任务(6.90 ± 4.35 μV)的平均波幅显著小于吸引力判断任务(7.56 ± 4.19 μV)。组别、图片类型和任务类型的交互作用显著,F(1, 58) =12.64,p= 0.001,= 0.18。简单效应分析发现, 对疼痛图片, 自闭特质组(8.87 ± 4.38 μV)在吸引力判断任务中平均波幅显著大于控制组(6.44 ± 3.84 μV),F(1, 58) = 5.19,p= 0.026,= 0.08; 自闭特质组在疼痛判断任务中与控制组没有显著差异[自闭特质组:7.72 ± 5.17 μV, 控制组:6.51 ± 3.43 μV],F(1,58) = 1.15,p= 0.287。对非疼痛图片, 自闭特质组在疼痛判断任务[自闭特质组:7.60 ± 5.01 μV, 控制组:5.77 ± 3.36 μV;F(1, 58) = 2.74,p= 0.103]和吸引力判断任务[自闭特质组:8.15 ± 4.31 μV, 控制组:6.77 ± 3.94 μV;F(1, 58) = 1.69,p= 0.199]中与控制组均没有显著差异。
LPC 任务类型的主效应显著,F(1, 58) =38.80,p< 0.001,= 0.40, 疼痛判断任务(5.60 ±3.37 μV)的平均波幅显著小于吸引力判断任务(7.69 ± 4.03 μV)。图片类型和任务类型的交互作用显著,F(1, 58) = 10.25,p= 0.002,= 0.15。疼痛判断任务中, 疼痛图片(5.87 ± 3.55 μV)的平均波幅显著大于非疼痛图片(5.34 ± 3.18 μV),F(1, 58) = 4.43,p= 0.040,= 0.07; 吸引力判断任务中, 疼痛图片(7.44 ± 3.90 μV)的平均波幅显著小于非疼痛图片(7.94 ± 4.18 μV),F(1, 58) = 6.07,p= 0.017,=0.10。组别、图片类型和任务类型的交互作用显著,F(1, 58) = 11.98,p= 0.001,= 0.17。简单效应分析显示, 对控制组, 疼痛判断任务中疼痛图片(5.86 ±3.02 μV)的平均波幅显著大于非疼痛图片(4.91 ±2.87 μV),F(1, 58) = 7.16,p= 0.010,= 0.11; 吸引力判断任务中疼痛图片(6.69 ± 3.47 μV)的平均波幅显著小于非疼痛图片(7.90 ± 3.94 μV),F(1, 58) =17.11,p< 0.001,= 0.23。对自闭特质组, 疼痛判断任务[疼痛图片:5.87 ± 4.07 μV, 非疼痛图片:5.76 ± 3.46 μV;F(1, 58) = 0.09,p= 0.765]和吸引力判断任务[疼痛图片:8.18 ± 4.21 μV, 非疼痛图片:7.99 ± 4.47 μV;F(1, 58) = 0.43,p= 0.516]中疼痛图片和非疼痛图片没有显著差异。
3.3 相关分析
将每个被试在AQ问卷上的得分与其分别进行疼痛判断任务和吸引力判断任务时诱发的各个ERP成分的平均波幅做皮尔逊积差相关分析。结果发现, 在 P3成分上存在一个显著相关:对疼痛图片进行吸引力判断任务时的平均波幅与被试的 AQ分数呈现显著正相关(r= 0.28,p= 0.029)。说明个体的AQ分数越高, 对疼痛图片进行吸引力判断时诱发的波幅越大。
4 讨论
本文以疼痛面孔图片作为刺激材料, 选取自闭特质组和控制组进行 ERP实验以探究注意线索对自闭特质个体面孔疼痛共情的影响。结果发现, 当自闭特质组完成吸引力判断任务时, 疼痛图片诱发的顶枕叶 P3平均波幅显著大于控制组; 而当完成疼痛判断任务时, 两组没有显著差异。但是, 在早期N1、P2和N2成分上都没有发现类似的差异, 即是注意指向或不指向疼痛线索时, 自闭特质组对他人疼痛面孔的早期共情神经反应均与控制组相似,说明自闭特质个体疼痛共情的早期加工阶段不会受到实验任务的影响。这可能是因为对疼痛面孔的早期情绪分享和疼痛感知是一个自动化的加工过程(Decety et al., 2010), 受到对疼痛线索自上而下注意的影响较小, 这也说明自闭特质个体的共情能力缺损可能主要体现在晚期的认知控制加工阶段。
上述结果支持了我们的假设, 说明了自闭特质个体对他人疼痛面孔的共情加工进程会受其对疼痛线索自上而下注意的调节, 并且这样的调节作用发生在疼痛共情的晚期加工阶段。相关分析的结果也支持了这一结论:个体的AQ分数与其对疼痛图片进行吸引力判断任务时诱发的 P3平均波幅呈显著正相关, 而与其对疼痛图片进行疼痛判断任务时诱发的 P3平均波幅不相关。说明当不注意疼痛线索时, 个体的 AQ分数越高, 对疼痛刺激的加工卷入的心理能量就越高。
对于反映面孔加工的 N170成分, 本研究结果发现注意面孔线索时, 颞枕区 N170平均波幅比不注意面孔线索时更大, 这与以往研究结果一致(Sreenivasan, Goldstein, Lustig, Rivas, & Jha, 2009),即注意指向面孔刺激时比不指向面孔刺激时诱发的 N170波幅更大。另外, 有研究发现面孔刺激相较于非面孔刺激也会诱发更大的N170波幅(Itier &Taylor, 2004; Rossion & Jacques, 2008), 说明将注意集中到面孔线索和面孔刺激本身都会引起对面孔更多的加工。N170反映的是对面孔结构的加工和面部空间关系的识别(Itier & Taylor, 2002), N170波幅增大说明主动注意面孔时增强了对他人面孔信息的感知水平。因此, 注意线索可能会调节个体对他人面孔的加工。
行为数据结果发现, 在疼痛判断任务中, 被试对疼痛图片的反应时长于对非疼痛图片的反应时;而在吸引力判断任务中, 对疼痛图片的反应时短于对非疼痛图片的反应时, 这与以往研究的结果是一致的(Meng, Shen, Li, & Peng, 2019a)。另外, 当注意疼痛线索时, 疼痛刺激诱发的晚期正成分 LPC平均波幅大于非疼痛刺激; 而当不注意疼痛线索时,疼痛刺激诱发的 LPC平均波幅小于非疼痛刺激。以往研究也得出了相似的结论(Mella, Studer, Gilet,& Labouvie-Vief, 2012), 说明要求注意疼痛线索时,个体会投入更多的心理资源来加工疼痛刺激, 从而需要更多的时间做出判断。这从行为和神经反应层面说明了注意和不注意疼痛线索时, 个体对疼痛刺激的加工模式可能存在差异。
本研究中发现注意和不注意疼痛线索时, 与疼痛共情相关的ERP成分(N2、P3和LPC)平均波幅存在差异, 这与先前研究的结果是一致的(Fan &Han, 2008; Mella et al., 2012; Meng et al., 2019b),说明了注意线索可能会影响个体的疼痛共情。前额叶 N2反应了个体早期阶段对他人疼痛的感知, 因此对疼痛线索自上而下的注意可能会影响疼痛共情早期加工阶段的认知资源分配。而在顶枕叶 P3和 LPC上, 发现注意疼痛线索时平均波幅比不注意疼痛线索时更小, 说明了疼痛共情的晚期加工阶段也会受到对疼痛线索自上而下注意的调节, 这一结果支持了以往研究的结论(Fan & Han, 2008)。但以往有研究发现, 注意疼痛线索时P3和LPC波幅比不注意疼痛线索时更大, 且认为P3和LPC反应了对他人疼痛的认知评估(Meng et al., 2019b), 可能是由于本研究中的吸引力判断任务更为复杂, 使得不注意疼痛线索时可能对疼痛刺激产生了更复杂的认知评估, 占据了更多的心理资源, 从而波幅增大。此外, 在控制组上发现完成疼痛判断任务时疼痛图片诱发的 P2平均波幅大于非疼痛图片, 这与以往研究结果一致(Meng et al., 2019a), 但在自闭特质组上两种图片没有差异, 这提示注意线索对自闭特质个体和控制组疼痛共情加工进程的调节作用可能不一致。
此外, 本研究结果发现相比于控制组, 自闭特质组对疼痛图片进行吸引力判断任务时诱发的 P3平均波幅更大, 但是对疼痛图片进行疼痛判断任务时两组没有显著差异。说明当自闭特质个体不注意疼痛线索时, 可能比控制组对疼痛面孔刺激加工的程度更高; 而注意疼痛线索时, 两组对他人的疼痛面孔有相似的神经反应。这说明视觉模态的注意线索可能会影响自闭特质个体对他人疼痛共情的加工进程。但先前研究以手脚图片作为视觉注意线索,发现注意线索对自闭特质个体疼痛共情的加工进程不存在调节作用, 因为无论注意指向或者不指向疼痛线索, 自闭特质个体和控制组都有相似的神经反应(Meng et al., 2019b)。这可能是由于自闭特质个体和控制组对手脚图片的注意特征相似, 而对疼痛面孔的注意模式存在差异, 因为以往研究认为ASD个体可能存在异常的面孔加工模式(Khorrami et al., 2013)。另外, 在疼痛判断任务中, 发现自闭特质组与控制组在加工疼痛图片时 P3波幅上没有差异, 说明当自闭特质个体主动注意疼痛线索时,其疼痛共情的加工进程与控制组相似, 并没有表现出明显的共情缺损。这为先前研究的观点提供了证据, 即ASD个体的共情缺陷可能是其不能自发、主动注意并加工社会信息导致的(Begeer et al., 2008;Senju, 2013)。
P3反应了疼痛共情加工过程中晚期的认知评估和认知控制, 以及对刺激的有意识评价(Song et al., 2016)。以往研究发现疼痛刺激会比非疼痛刺激诱发更大的P3波幅(Meng et al., 2012), 本研究结果与此一致, 说明疼痛刺激引起的认知加工会卷入个体更多的心理资源。因此, 自闭特质组比控制组对疼痛图片进行吸引力判断任务时诱发的 P3波幅更大, 说明相比控制组, 自闭特质个体在不注意疼痛线索时需要将更多的认知资源投入到疼痛刺激加工进程中, 需要更为复杂的认知评价和认知控制加工。
这可能是由于, 一方面, 相比控制组, 自闭特质个体在注意指向面孔刺激时可能对面孔信息产生了回避, 从而更多地将注意资源分配到了疼痛刺激上, 使得P3波幅增大。因为以往研究发现ASD个体存在对他人面孔信息的回避(Chita-Tegmark,2016; Key & Stone, 2012)。从这个角度该研究结果为自闭特质个体回避面孔信息提供了证据。
另一方面, P3波幅增大可能是因为自闭特质个体加工面孔刺激时比控制组更容易受到疼痛刺激的干扰。根据认知负荷理论(Lavie, Hirst, de Fockert,& Viding, 2004), 高认知负荷下干扰刺激会得到更多的加工, 以往研究也发现个体在高认知负荷下会更多的加工他人的疼痛刺激(Cheng, Luo, & Cui,2017)。自闭特质个体在注意不指向疼痛线索时的共情能力减弱, 以及异常的面孔加工可能使其加工疼痛刺激需要投入更多的认知资源, 从而产生了更大的认知负荷, 使得在要求注意面孔时可能更容易对疼痛这一干扰刺激进行加工, 导致 P3波幅大于控制组。总之, 不注意疼痛线索时, 自闭特质个体和控制组在 P3成分上的差异说明了自闭特质个体存在对他人疼痛面孔特异的共情认知神经反应, 并且会受到对疼痛线索自上而下注意的调节。
综上所述, 本研究采用疼痛面孔图片, 通过让自闭特质组和控制组分别完成疼痛判断任务和吸引力判断任务, 使他们分别将注意指向疼痛线索和不指向疼痛线索, 来考察注意线索对自闭特质个体面孔疼痛共情的影响。最终发现对视觉疼痛线索自上而下的注意可能会调节自闭特质个体疼痛共情的晚期加工进程, 这样的影响体现在当自闭特质个体注意疼痛线索时与控制组有相似的共情神经反应; 而当自闭特质个体不注意疼痛线索时, 对他人疼痛面孔的加工程度更高。这一结果也说明自闭特质个体可能存在对他人面孔信息的回避, 以及为自闭特质个体的共情缺损提供了证据。上述结果可以帮助我们进一步了解 ASD个体疼痛共情的特点和影响因素, 以及ASD个体对面孔的加工机制。
本研究也存在几处问题和局限。首先, 研究中采用的是单一合成的图片, 以后研究可以制作更贴近实际生活情境的图片, 提高研究的外部效度。其次, 本研究选取的被试年龄跨度较小, 可能导致得出的结论解释性有限。最后, Jemel等认为ASD个体对整体面孔的识别能力并不存在缺陷(Jemel,Mottron, & Dawson, 2006), 未来研究可以结合注意线索进一步探讨ASD个体对整体和局部(眼部或嘴部)面孔的加工机制。
猜你喜欢
辽河(2022年4期)2022-06-09 01:56:03
癫痫与神经电生理学杂志(2022年6期)2022-02-10 13:55:46
智慧少年·故事叮当(2020年2期)2020-03-08 14:19:16
青少年科技博览(中学版)(2019年7期)2019-10-11 04:13:55
人与自然(2019年4期)2019-07-26 12:52:37
文教资料(2019年31期)2019-01-14 02:32:05
价值工程(2018年25期)2018-09-26 10:14:36
校园英语·中旬(2016年8期)2016-07-09 21:25:53
西南交通大学学报(2016年4期)2016-06-15 20:29:36
华北电力大学学报(自然科学版)(2016年3期)2016-04-25 05:06:56
