
漓江大型底栖动物功能摄食类群时空分布及生态效应
2020-03-13 03:15朱晨曦莫康乐林育青陈求稳
生态学报 2020年1期
朱晨曦,莫康乐,唐 磊,吴 焱,李 婷,林育青,陈求稳
南京水利科学研究院, 南京 210029
河流生态系统由生物和生境两部分组成,两者一直处于动态变化的过程中。其中,生物是河流的生命系统,生境是河流生物的生命支持系统[1]。河流大型底栖动物是河流生态系统食物链结构中的重要环节,对河流生态系统的物质循环、能量流动有着积极作用[2]。同时,由于大型底栖动物的活动能力弱、活动范围小,对于外来污染具有较小的规避能力,并且对外界胁迫较为敏感,因此能有效的指示河流生态系统的健康,其种类组成及时空分布更能体现生境变化对于生物的影响[3-4]。
大型底栖动物依据取食的不同划分为5个功能摄食类群(Functional feeding groups, FFGs):捕食者、撕食者、刮食者、收集者和滤食者[5- 8]。大型底栖动物功能摄食类群与传统大型底栖动物形态分类方法不同,它主要依据底栖动物的食物资源类型及在获取食物过程中形态学的适应机制,来反映生境变化对大型底栖动物群落的影响,揭示出大型底栖动物的群落结构特征和生境适应性特征[9]。大型底栖动物功能摄食类群能较好的反映人类活动对河流生态系统的影响及河流生态系统的受损情况,因此近年来受到了研究者们的广泛关注[9- 11]。
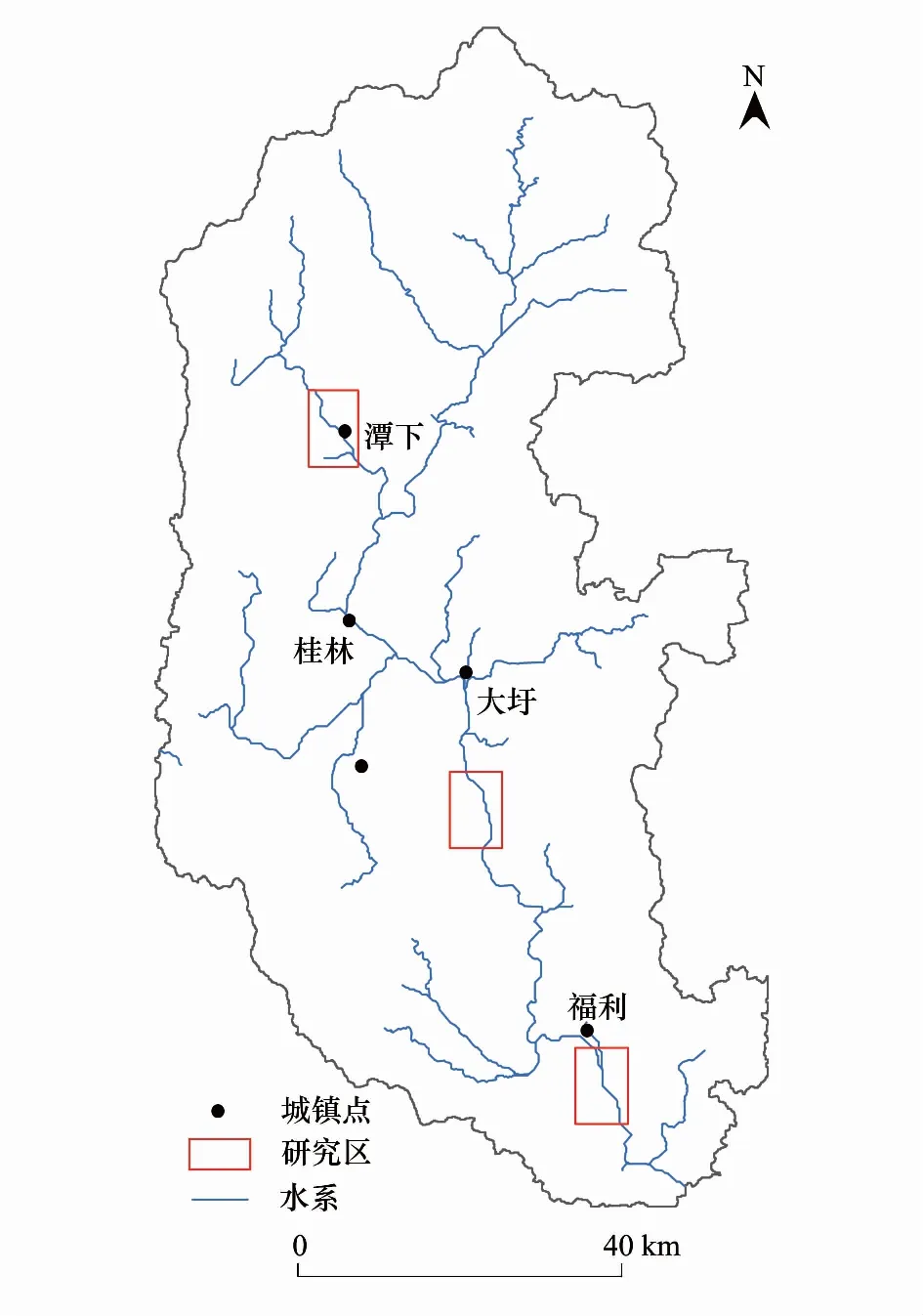
图1 漓江研究区域分布Fig.1 The study reas in Lijiang River
漓江作为中国唯一一条入选的全球最美河流,是桂林山水的灵魂,保护漓江河流生态系统健康在维持漓江山水自然风景方面有着重要的意义[12]。以往应用大型底栖动物对漓江进行评价的研究,大多基于传统的形态分类方法,考虑其与水环境、底质类型、沉水植物之间的关系[12-14],鲜有从功能摄食类群角度去评价漓江河流生态系统。本文以漓江为对象,研究大型底栖动物功能摄食类群的时空分布特征,并分析其与环境因子的相关性,采用基于功能摄食类群的生态参数评价漓江河流生态系统健康状态,为漓江生态环境保护提供依据。
1 材料与方法
1.1 区域概况及采样区设置
漓江属于珠江流域西江水系,是支流桂江上游河段的通称,位于中国的南部(110°18′—111°18′E,23°23′—25°59′N),发源于桂江源头越城岭猫儿山。漓江上游为从猫儿山到桂林河段、中游为从桂林到阳朔河段、下游为从阳朔到平乐河段,其中桂林到阳朔河段为著名的桂林漓江风景区。漓江流域总面积为12285 km2,主河道长214 km,主要包括甘棠江、黄柏江、川江、小溶江等几条支流。
本研究分别在漓江上、中、下游选取典型江段进行研究(图1)。其中,上游甘棠江研究江段长7 km,底质以细沙和淤泥为主;中游大圩研究江段长6 km,底质以卵石为主;下游福利研究江段长7 km,两岸植被稀少,主要为人工栽植的凤尾竹,底质以砾石为主。本研究在不同季节对长约20 km的研究江段进行大型底栖动物的采样调查,调查时段分别为2009年秋季(9月)、2010年春季(4月)、2012年冬季(1月)和夏季(8月)。4次调查的样本数分别为150个(上游54个,中游48个,下游48个)、150个(上游54个,中游41个,下游55个)、209个(上游55个,中游78个,下游76个)和165个(上游26个,中游75个,下游64个),并同步对部分水体理化指标及河流生境状况进行了测定和记录。
1.2 大型底栖无脊椎动物的采集与鉴定
本研究大型底栖动物的采集一共使用了两种采样工具,包括直径30 cm的40目D型网和面积为1/20 m2的Peterson改良式采泥器。在现场采集时,于每个采样点断面的两岸可涉水区域使用直径30 cm的40目D型网采集底栖动物样品共6个样方,采集面积1.8 m2;断面中间不可涉水区域使用面积为1/20 m2的Peterson改良式采泥器采集底栖动物样品共4个样方,采集面积0.2 m2。将每个断面采集的10个样方混合成一个样品,采集面积共计2 m2。在现场用40目分样筛筛选样品,挑出大型底栖无脊椎动物后放入100 mL标本瓶中,然后用10%的甲醛溶液保存并带回实验室进行种类鉴定、个体计数、称重。在鉴定时,所有样品尽量鉴定到最低分类单元[15-17]。
1.3 水体理化指标测定
在采集现场使用便携式多参数水质监测分析仪(YSI6600,美国金泉仪器)测定水温(Temp)、pH、溶解氧(DO)、电导率(EC)、浊度(Tur)、盐度(Salt),并在每个样点采集2瓶550 mL水样,其中一瓶现场加硫酸调整pH值至小于2,低温保存运回实验室后分别采用GB3838—2002[18]中规定的钼酸铵分光光度法、碱性过硫酸钾消解紫外分光光度法和高锰酸盐法测定总磷(TP)、总氮(TN)、化学需氧量(COD)。
1.4 大型底栖无脊椎动物功能摄食类群及生态系统属性划分
根据河流大型底栖动物的食性类型,参照Cummins等、Barbour等和Bode等的划分规则[5-8],将漓江大型底栖动物分为以下5类功能群(FFGs):(1)收集者(GC),主要以河底的各种有机物颗粒为食;(2)滤食者(FC),主要以水流中的细有机颗粒物为食(0.45 mm<粒径<1 mm);(3)捕食者(PR),主要以捕食其他水生动物为食;(4)撕食者(SH),主要以各种凋落物和粗有机质颗粒为食(粒径>1 mm);(5)刮食者(SC),主要以各种营固着生活的生物类群为食。
参考Yoshimura等及王博涵等的研究[9,19],本文根据大型底栖动物功能摄食类群的参数,从物质循环、物质纵向输送能力、沿岸物质输入等方面对漓江生态系统进行评价(表1)。

表1 大型底栖动物功能摄食类群所表征的生态系统属性
1.5 数据处理与分析
采用物种优势度指数(Y)来表示大型底栖动物各功能摄食类群中某一物种在其中所占优势的程度,公式如下:
Y=(ni/N)fi
式中,N为各功能摄食类群大型底栖动物的总密度,ni为第i种的密度,fi为第i种出现的频率。优势度指数Y>0.02的种类确定为本次调查各功能摄食类群的优势种[20]。
基础数据统计均在Excel软件中完成,前向筛选和冗余分析(RDA)在R软件中完成(version 3.3.2,R development Core Team, http://www.r-project.org/)。
2 结果
2.1 大型底栖无脊椎动物物种组成及功能摄食类群组成
本研究4次采样共采集到64个大型底栖无脊椎动物分类单元,隶属于3门8纲16目38科。其中昆虫纲28种、腹足纲25种、甲壳纲3种、瓣鳃纲3种、蛭纲2种、多毛纲1种、寡毛纲1种、蛛形纲1种。对漓江大型底栖无脊椎动物功能摄食类群进行分析,结果显示,刮食者在漓江占显著优势,相对丰度为73.26%;其次为滤食者和收集者,相对丰度分别为15.12%和8.85%;捕食者和撕食者相对丰度较小,分别为2.72%和0.06%。由表2可知收集者、滤食者、捕食者、撕食者、刮食者的优势物种,滤食者中的河蚬优势度最大为0.367,除撕食者外其他功能摄食类群均存在两种优势物种。
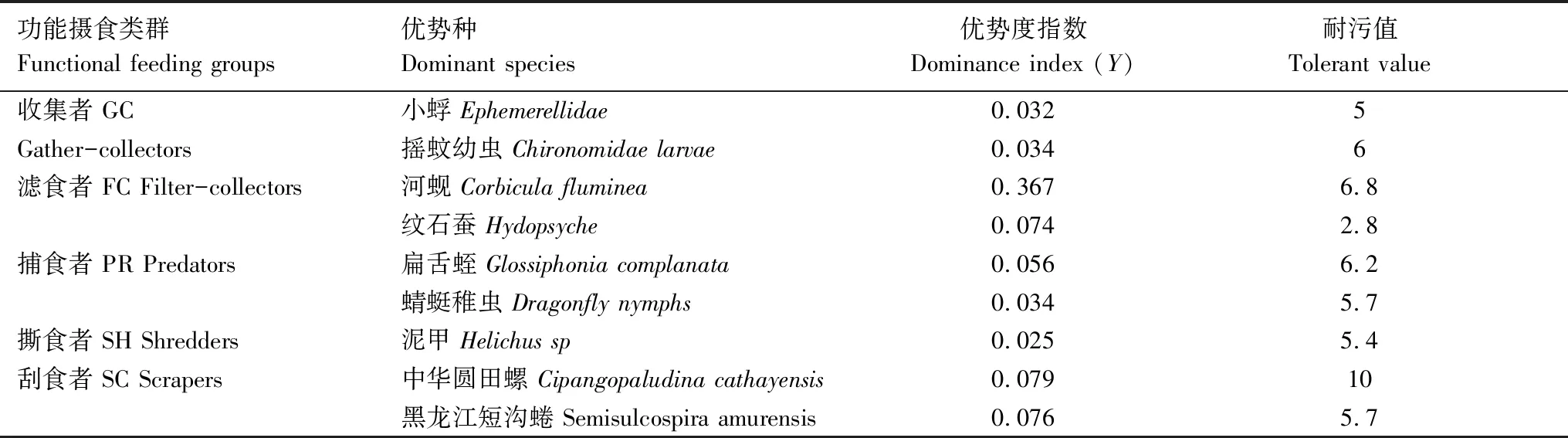
表2 漓江大型底栖无脊椎动物各功能摄食类群优势种
2.2 大型底栖无脊椎动物功能摄食类群空间分布特征
2.2.1相对丰度空间分布特征
漓江大型底栖无脊椎动物功能摄食类群的相对丰度空间分布见图2。优势类群刮食者的最大相对丰度出现在夏季中游江段,为93.48%,最小出现在夏季上游江段,为25.27%;捕食者的最大相对丰度出现在冬季上游江段,为8.88%,最小出现在夏季中游江段,为0.75%;滤食者的最大相对丰度出现在夏季上游江段,为55.05%,最小出现在冬季中游江段,为3.79%;收集者的最大相对丰度出现在春季上游江段,为29.36%,最小出现在秋季下游江段,为0.62%;撕食者相对丰度在各江段都很小,只在夏季上游江段和秋季上游江段相对较大,分别为0.53%、0.42%。
2.2.2密度的空间分布特征
漓江大型底栖无脊椎动物功能摄食类群的密度空间分布如图3所示。优势类群刮食者的最大密度出现在夏季中游江段,为2354个/m2,最小出现在夏季上游江段,为73个/m2;捕食者的最大密度出现在春季下游江段,为84个/m2,最小出现在秋季上游江段,为4个/m2;滤食者的最大密度出现在秋季中游江段,为436个/m2,最小出现在冬季中游江段,为49个/m2;收集者的最大密度出现在春季中游江段,为208个/m2,最小出现在秋季下游江段,为5个/m2;撕食者密度在各江段都很小,只在春季、夏季、秋季的上游江段以及秋季的中游江段相对较大,分别为1、2、2、2个/m2。
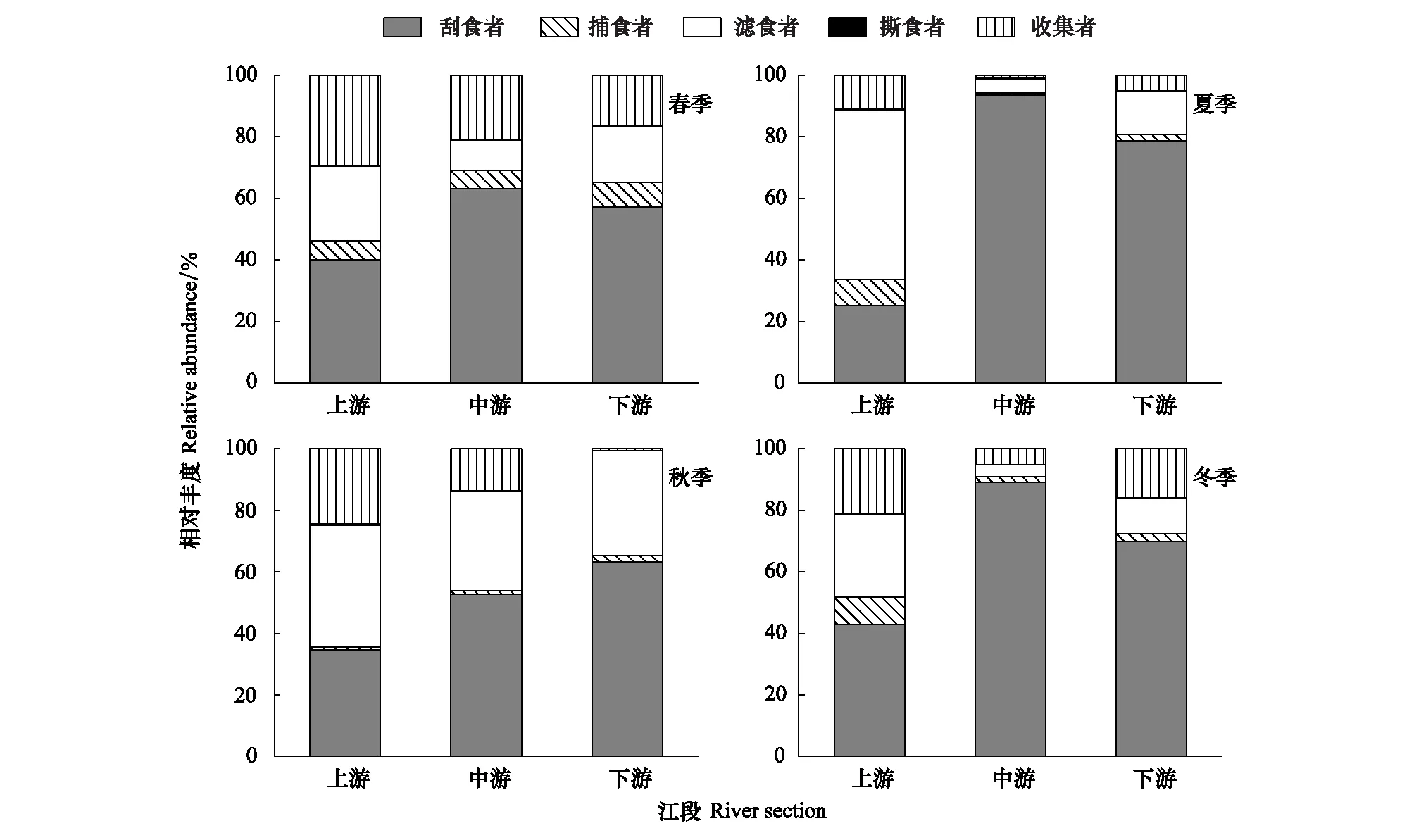
图2 各季节漓江上中下游大型底栖动物功能摄食类群相对丰度Fig.2 The relative abundance of macroinvertebrate FFGs in Lijiang River among different river section in different seasonsFFGs: 功能摄食类群Functional feeding groups; SC:刮食者 Scrapers;PR:捕食者 Predators;FC:滤食者 Filter-collectors;SH:撕食者 Shredders;GC:收集者 Gather-collectors
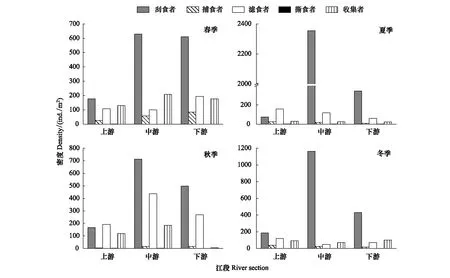
图3 各季节漓江上中下游大型底栖动物功能摄食类群密度Fig.3 The density of macroinvertebrate FFGs in Lijiang River among different river section in different seasons
2.3 大型底栖无脊椎物功能摄食类群季节变化特征
2.3.1相对丰度季节变化特征
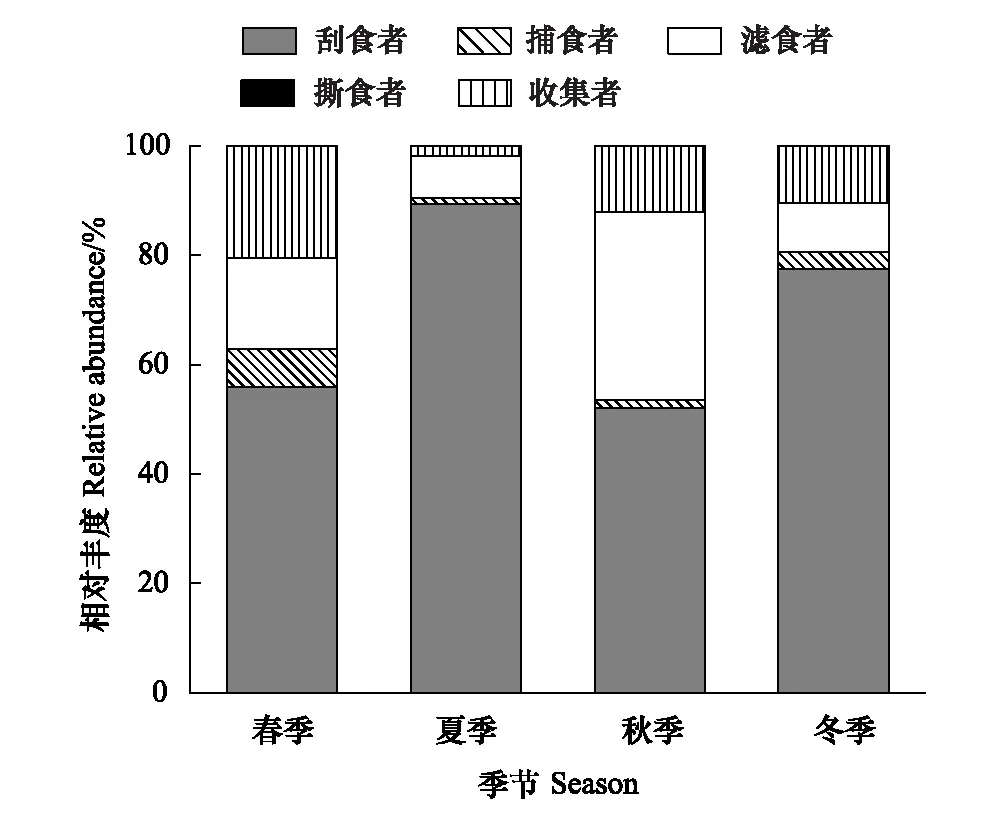
图4 漓江大型底栖动物各功能摄食类群相对丰度的季节动态 Fig.4 The relative abundance of macroinvertebrate FFGs in different seasons in Lijiang River
图4显示了功能摄食类群相对丰度的季节变化。各季节刮食者相对丰度明显高于其他功能摄食类群,一直处于优势地位,最大值出现在夏季,为89.37%,最小值出现在秋季,为52.12%;撕食者相对丰度明显低于其他功能摄食类群,一直处于劣势地位,最大值出现在秋季,为0.17%,最小值出现在冬季,为0.01%;滤食者相对丰度最大值出现在秋季,为34.31%,最小值出现在夏季,为7.54%;收集者相对丰度最大值出现在春季,为20.46%,最小值出现在夏季,为1.86%;捕食者相对丰度最大值出现在春季,为6.86%,最小值出现在夏季,为1.16%。
2.3.2密度季节变化特征
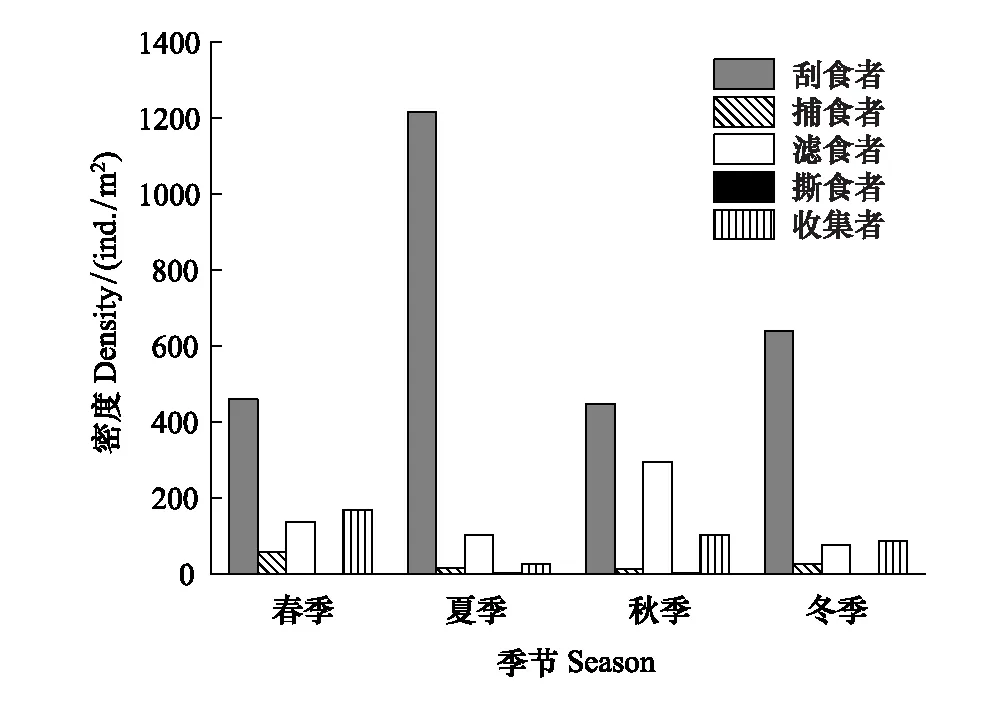
图5 漓江大型底栖动物功能摄食类群密度的季节动态 Fig.5 The density of macroinvertebrate FFGs in different seasons in Lijiang River
功能摄食类群密度的季节变化见图5。各季节刮食者密度明显高于其他功能摄食类群,优势地位明显,最大值出现在夏季,为1215个/m2,最小值出现在秋季,为447个/m2;撕食者密度明显低于其他功能摄食类群,劣势地位明显,最大值出现在秋季,为2个/m2,最小值出现在冬季,为0个/m2;滤食者密度最大值出现在秋季,为294个/m2,最小值出现在冬季,为75个/m2;收集者密度最大值出现在春季,为168个/m2,最小值出现在夏季,为25个/m2;捕食者密度最大值出现在春季,为56个/m2,最小值出现在秋季,为12个/m2。
2.4 基于大型底栖无脊椎动物功能摄食类群参数的漓江生态系统评价
基于功能摄食类群的各项参数进行时空差异性分析表明(表3):在物质循环方面,底栖动物初级生产力(F1)在漓江中游大于其余区域,夏季明显大于其他季节;自养/异养(F2)在中游大于其余区域,夏季明显大于其他季节;底栖动物分解能力(F3)在中游大于其余区域、夏季小于其他季节;底栖动物次级生产力(F4)从上游到下游逐渐增高。在物质纵向输送能力方面,纵向输送能力(F5)和相对纵向输送能力(F6)中下游较高、夏季明显较大。在沿岸物质输入方面,沿岸物质输入量(F7)和相对物质输入量(F8)均为上游高于中下游,秋季高于其他季节。在其他方面,捕食者的下行效应(F9)在河流空间分布上差异性不大、四季变化也不大,但参数水平普遍较低;生境稳定性(F10)表现为从河流上游到下游逐渐增强。
综合上述分析,对漓江生态系统进行评价。结果表明:漓江生态系统在空间上从上游到下游大型无脊椎底栖动物的生物量逐渐增加且生境的稳定性越来越强;在时间上表现为夏季漓江生态系统各项参数水平均较高,生境稳定性也最强。但从参数F9可以看出漓江生态系统内底栖动物的高营养级对低营养级控制力不强[21]。
2.5 大型底栖无脊椎动物功能摄食类群与环境因子的相关性分析
为了探究采样期间不同环境因子对漓江大型底栖动物功能摄食类群的影响,对功能摄食类群与环境因子DO、pH、EC、Temp、Tur、Salt、TN、TP和COD进行冗余分析(RDA)(图6),通过前向选择(forward selection)排除掉贡献较小的环境因子(P>0.05)。分析结果显示,漓江四季对大型底栖动物功能摄食类群影响较大的环境因子有所不同,春季COD(P=0.001)和DO(P=0.005)两个环境因子与漓江大型底栖动物功能摄食类群相关性较强;夏季EC(P=0.001)、pH(P=0.002)和DO(P=0.002)等3个环境因子与功能摄食类群相关性较强;秋季EC(P=0.001)和TN(P=0.024)与功能摄食类群相关性较强;冬季Temp(P=0.001)和Tur(P=0.001)与功能摄食类群相关性较强。
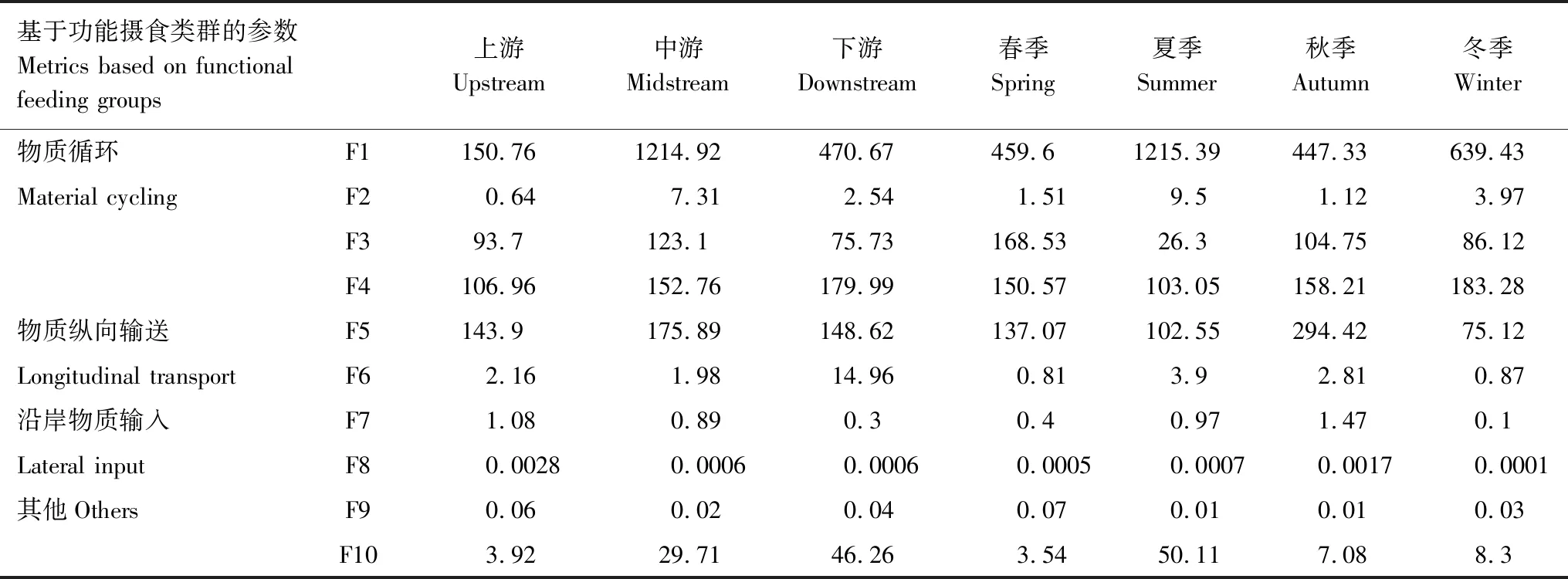
表3 漓江大型底栖无脊椎动物功能摄食类群参数的时空变化
F1:刮食者密度 Density of scrapers;F2:刮食者密度与滤食者、收集者之和的比率 Ratio of scrapers to filterers and gathering-collectors;F3:撕食者密度和收集者密度之和 Density of shredders and gathering-collectors;F4:生物量 Biomass;F5:滤食者密度 Density of filterers;F6:滤食者与撕食者和收集者之和的比率 Ratio of filterers to shredders and gather-collectors;F7:撕食者密度 Density of shredders;F8:撕食者密度与总密度之比 Ratio of shredders to total density;F9:捕食者密度与总密度之比 Ratio of predators to total density;F10:刮食者和滤食者之和与撕食者和收集者之和的比率 Ratio of scrapers and filterers to total shredders and gather-collectors
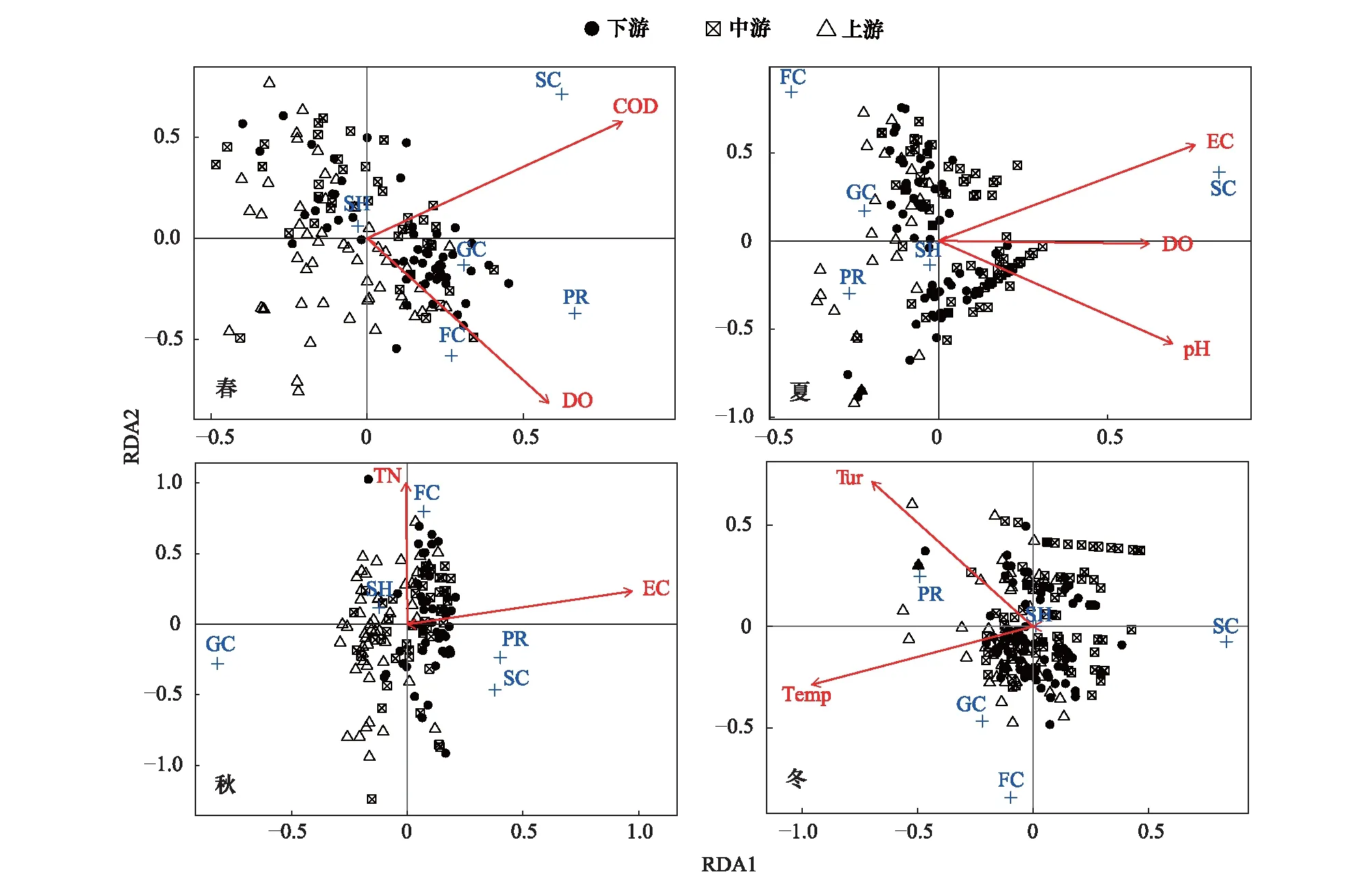
图6 漓江各季节大型底栖动物功能摄食类群与环境因子的冗余分析(RDA)Fig.6 Relationships of macroinvertebrate FFGs and environmental factors obtained by Redundancy analysis (RDA) in different seasons in Lijiang RiverRDA1:冗余分析第一轴 Redundancy analysis first axis;RDA2:冗余分析第二轴 Redundancy analysis second axis;COD:化学需氧量 Chemical oxygen demand;DO:溶解氧 Dissolved oxygen;EC:电导率 Electrical conductivity;TN:总氮 Ttotal nitrogen;Tur:浊度 Turbidity;Temp:温度 Temperature
3 讨论
3.1 漓江大型底栖无脊椎动物功能摄食类群的组成与时空分布
大型底栖动物功能摄食类群的组成与生境密切相关。漓江大型底栖动物以刮食者占据绝对优势,撕食者处于劣势,这一结果与国内其他研究区域如香溪河[22]、昌江[23]等地区的研究结果存在差异。这是由于漓江流域属于喀斯特地貌,河道底质多为沙卵石加细沙,水浅流急,四季清澈见底,适宜沉水植物生长。漓江茂盛的沉水植物[24]为螺类等刮食者提供了适宜的生境[25],使得刮食者在组成上占据优势。撕食者数量少的原因是漓江两岸多为河漫滩与深切河谷,灌乔木稀少,无法给撕食者提供枯枝落叶和荫蔽的条件,进而撕食者难以生存[26]。
河流连续性概念认为功能摄食类群的空间分布与自然生境中有机物沿河流的运输、利用和存储有密切的关系[27],但人类活动对其空间上的分布也产生了较大的影响。按照经典的河流连续性概念预测,对溪流来讲,上游河流中撕食者的相对丰度较大[28],捕食者在河流上中下游空间上变化不大[26],刮食者在中游河段的相对丰度最大[29]。本研究观测结果与上述情况基本吻合。在自然河流中,细颗粒有机物在很大程度上是由粗颗粒有机物分解形成的,其比例自上游至下游逐渐增加,按河流连续性概念预测滤食者和收集者的相对丰度也应自上游至下游逐渐增加[30]。但在本研究中,这两者的相对丰度表现为上游最大、中下游符合上述规律,这与预测情况偏差较大,原因是本研究所观测到的滤食者和收集者的优势种摇蚊幼虫、小蜉和河蚬均为耐污种,而上游研究区位于漓江支流甘棠江的下游河段,该河段流经村镇,大量使用的化肥、村庄的生活污水这些有机物输入到河流中,为一些滤食者和收集者的生长和繁荣创造了条件[29],并致使耐污种比例增高;中下游研究区域均为旅游观光区,两岸基本无污染排放。这也与功能摄食类群参数F10相对应,可推断村镇居民生活对漓江生态坏境的影响大于旅游观光的影响。
底栖动物功能摄食类群的季节性分布主要是由于季节变化改变了岸边植被状态及河流水文条件,进而使底栖动物的生境发生了微变化,最终导致其时间分布上的差异性。撕食者的分布能够反映树叶在河道底部的堆积情况[31]。在本研究中,撕食者的相对丰度和密度在秋季明显高于其他3个季节,主要是因为树木的落叶期主要集中在秋季。捕食者的相对丰度和密度在春季高于其他季节,是因为春季其他类群密度较大,为捕食者提供了丰富的食物[9]。刮食者的相对丰度和密度在夏季明显高于其他季节,主要是因为在夏季繁殖旺盛的中华圆田螺[32]为该功能类群的优势种,夏季沉水植物的生长也为其提供了适宜的生境。收集者主要以沉积的有机颗粒为食,在流速低的区域有机颗粒较易沉积,因而收集者偏爱流速较低的生境[33]。本研究中收集者的相对丰度和密度在春、秋和冬季无太大差异,在夏季却很低,这主要是因为夏季是汛期,河流流速增大,不适宜收集者进食。滤食者以悬浮的细有机颗粒物为食,一般都存在一个最适滤食流速[22],在本研究中滤食者相对丰度和密度在秋季明显高于其他季节,是因为滤食者密度主要受到水流流速及食物影响,秋季处于洪水刚过的时期,水面趋于平稳、流速缓慢,而且撕食者的优势季节也处在这一时期,其取食粗颗粒有机质后为滤食者提供了更多的细颗粒有机质[26]。
3.2 大型底栖动物无脊椎功能摄食类群与水环境因子之间的关系
在河流生态学研究中确定影响大型底栖动物功能摄食类群的关键环境因子对分析河流生态健康起着重要作用[34]。研究指出,环境因子对底栖动物的影响在不同区域不同季节各有差异,较为复杂[34-35]。本研究中RDA分析得出7个环境因子(DO、pH、EC、Temp、Tur、COD、TN)对漓江大型底栖动物功能摄食类群组成与分布具有关键影响,主要反映了水体的有机污染物和物理因素。春季COD与刮食者呈显著的正相关性,这主要是因为COD反映了水中有机质的含量或者说有机物污染程度,而本研究中刮食者是以中华园田螺等耐污种为优势种,这与其他研究中COD与刮食者的响应关系结论相同[36]。春季和夏季DO与漓江大型底栖动物功能摄食类群相关性较强,这是因为春季和夏季是底栖动物生长和繁殖的季节,其新陈代谢增强,摄食活动会大量消耗水中的溶解氧,所以影响了底栖动物的组成与分布[37]。夏季pH与底栖动物功能摄食类群有较强的相关性,是因为绝大部分底栖动物适宜的pH值为6—8[37-38],而本研究中夏季pH值达到最大为8.49,超出了适宜范围,对底栖动物的生存产生了影响。天然水体中的pH主要取决于游离二氧化碳的含量及碳酸平衡,夏季植物生长旺盛光合作用强烈致使溶于水体中的二氧化碳减少导致pH值变大。夏季和秋季EC与漓江底栖动物功能摄食类群相关性较强,这是由于电导率是度量水环境总溶解离子量的参数,而离子主要通过影响细胞膜渗透压的平衡对大型底栖动物产生了影响[39-40]。已有研究指出氮元素与底栖动物功能摄食类群有较好相关性[23,41],本研究中秋季TN与滤食者呈显著正相关,这与吴东浩等在西苕溪上游得到TN与滤食者呈负相关的结论相反[42],这是因为漓江的滤食者中优势种河蚬为耐污种。冬季Temp与底栖动物功能摄食类群有较强的相关性,这是因为冬季温度过低致使底栖动物的活动减弱[37],使得温度成为冬季限制底栖动物的主要环境因子。不同时期不同区域对底栖动物功能摄食类群起主要影响的环境因子不尽相同,需要充分探究底栖动物的生活习性与生境状况之间的响应关系。
4 结论
漓江大型底栖无脊椎动物功能摄食类群中以刮食者占据绝对优势地位,撕食者和捕食者处于劣势地位。基于大型底栖无脊椎动物功能摄食类群的时空分布特征分析可知,各功能摄食类群的时空分布可反映漓江生境在时空上的微变化及人类活动对生境的干扰,其分布特征明显受食物资源时空变化的限制。在春季,对功能摄食类群有较强影响的环境因子为COD和DO,夏季为EC、DO和pH,秋季为TN和EC,而在冬季为Tur和Temp。基于功能摄食类群参数对漓江生态系统评价,可知漓江中下游生境稳定性明显高于上游,村镇居民生活对漓江生态坏境的影响大于旅游观光的影响。
猜你喜欢
科学之谜(2021年4期)2021-07-09
生态学报(2021年3期)2021-03-31
中国水利(2020年23期)2020-12-24
西湖(2020年8期)2020-08-13
科学导报(2020年27期)2020-05-13
绿色科技(2019年14期)2019-11-19
简帛(2019年2期)2019-11-03
江苏农业科学(2019年5期)2019-09-02
广东农业科学(2017年5期)2017-08-29
科学之友(2017年3期)2017-03-24
