
PRRSV GP2a胞外区的原核表达及免疫原性分析
2020-03-12 00:40:42扣莉云张改平史西保张小转车志芬周景明祁艳华王爱萍郑州大学生命科学学院
中国兽医学报 2020年2期
扣莉云,张改平,2,史西保,张小转,车志芬,周景明,祁艳华,王爱萍∗ (.郑州大学 生命科学学院,
河南 郑州450001;2.河南农业大学 牧医工程学院,河南 郑州450002;3.河南师范大学 生命科学学院,河南 新乡
453007)
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)是由猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)引起的一种高度传染性疾病,是造成养猪业重大经济损失的主要疾病之一。PRRSV 基因组全长约15.1 kb,具有10个开放阅读框。其中,GP2a是由ORF2编码的跨膜蛋白,是病毒囊膜外凸的主要组成部分,研究表明GP2a具有抑制细胞凋亡[1]和引起T 淋巴细胞增殖[2]的作用。GP2a蛋白能与PRRSV 的宿主受体CD163 结合,作为病毒吸附蛋白介导病毒进入宿主细胞[3]。一旦PRRSV 缺失GP2a,便不能在MARC-145细胞中生长,但GP2a的C末端缺失10个氨基酸并不影响其在MARC-145细胞中的生长,也不改变重组病毒的中和抗原表位[4],因此,GP2a可能在PRRSV 感染与调控宿主免疫中发挥重要作用。因为PRRSV 的GP2a蛋白的信号肽序列和疏水性跨膜区使得GP2a的体外表达十分困难[5],所以限制了对GP2a的研究。
目前,刘金霞等[6]使用原核表达载体pGEX-6p-1实现了PRRSV CH-1a毒株GP2a的38-208aa的表达;刘明莉等[7]使用原核表达载体p GEX-6p-1实现了PRRSV Hn-1/06毒株GP2a的41-256aa的表达,但对缺失N 端和C 端的GP2a胞外区41-208aa的原核表达还未见报道。因为昆虫杆状病毒表达系统和真核表达系统存在表达效率低和难以纯化等缺点[8],及原核表达具有快速、简便、成本低且具有利于对蛋白的理化性质、结构及功能进行研究等优点[9],所以本研究使用带有可溶标签的原核表达载体p ET-32a对PRRSV BJ-4毒株的GP2a胞外区进行原核表达与纯化,并对重组蛋白的免疫原性进行研究,为深入研究PRRSV GP2a蛋白的相关功能奠定理论基础。
1 材料与方法
1.1 材料
1.1.1 质粒载体与菌株 pET-32a、pcDNA3.1-ORF2由河南省动物免疫学重点实验室提供;大肠杆菌DH5α、Rosetta(DE3)由郑州大学生命科学学院动物分子免疫学实验室保存。
1.1.2 主要试剂和酶 PCR 引物由生工生物工程(上海)股份有限公司合成;Ex Taq 酶、Bam HⅠ限制性内切酶、SalⅠ限制性内切酶、T4DNA 连接酶、Ta KaRa MiniBEST Plasmid Purification Kit、Ta KaRa MiniBEST Agarose Gel DNA Extraction Kit Ver.4.0等购自Ta KaRa宝生物工程(大连)有限公司;异丙基-β-D-硫代半乳糖苷(IPTG)、氨苄青霉素(ampicillin)、氯霉素(chloromycetin)购自Solarbio公司;HRP标记的羊抗鼠IgG 和抗His标签的单克隆抗体购自郑州益康生物有限公司。
1.1.3 主要仪器 普通离心机购自上海湘仪有限公司;Gel Doc XR 凝胶成像系统、PowerPac 3000电泳仪、半干转膜仪购自美国Bio-Rad;DK-3D 电热恒温水浴锅购自金坛市晶玻实验仪器厂;超声波破碎仪购自美国SONIC 公司;低温冷冻离心机购自德国EPPENDORF公司;Barnstead Nanopure超纯水仪购自美国Thermo公司。
1.2 方法
1.2.1 引物设计 根据真核质粒pc DNA3.1-ORF2的基因序列,使用Primer Premier 5.0 设计针对GP2a的41~208氨基酸的特异性引物(上游引物序列:5′-AAGGATCCTCACCATCGCCGGTTGGCTG-3′;下游引物序列:5′-AAGTCGACTTATTGCTGAAAATCATGAAGCTTTGGCC-3′,其中加粗碱基为保护碱基,倾斜碱基为限制性内切酶序列,划线碱基为所添加的终止子)由生工生物工程(上海)有限公司合成。
1.2.2 PCR 扩增目的基因 PCR 反应体系:Ex Taq酶25μL,上、下游引物各1.5μL,pcDNA3.1-ORF2重组质粒1μL,加入去离子水21μL 补足50μL。PCR 反应条件:95℃预变性180 s;94℃变性45 s,56℃退火45 s,72℃延伸90 s,经过34个循环;72℃延伸420 s,终止后4℃保存。经1% 核酸胶鉴定后,切胶回收。
1.2.3 目的基因与载体连接 将回收的目的基因与载体进行双酶切,体系:Bam HⅠ酶和SalⅠ酶各1.5μL,10×T Buffer 6μL,目的基因或质粒载体6μg,加去离子水补足到总体积60μL。其中目的基因37℃酶切4 h,载体37℃酶切1 h,酶切后,切胶回收。将双酶切的目的基因和载体连接,连接体系:T4DNA 连接酶1μL,10×T4DNA Ligase Buffer 2μL,DNA 片段约1 pmol,质粒载体约0.1 pmol,加去离子水补足到20μL。16℃过夜连接。连接产物通过化学转化的方法转化到感受态细胞DH5α中,均匀涂布于含氨苄青霉素的LB固体培养基上,倒置,37℃培养12~16 h。
1.2.4 重组质粒的鉴定 挑取平板上的单克隆菌落,接种于5 m L 含氨苄青霉素的LB 液体培养基中,37℃、220 r/min,摇菌8 h后,冻存菌种,其余菌液继续过夜摇菌,用于提取质粒和双酶切鉴定。质粒PCR 和双酶切过程同1.2.2和1.2.3。经上海生工测序正确的重组质粒命名为p ET-32a-eORF2。
1.2.5 重组蛋白的表达及Western blot鉴定 重组质粒p ET-32a-eORF2 转化到Rosetta(DE3)感受态细胞,挑取单克隆,在含有氨苄青霉素与氯霉素的LB液体培养基中,37℃、220 r/min摇菌16 h;以1∶100的比例接种于含有氨苄青霉素与氯霉素的LB液体培养基,继续摇菌约1.5 h;当D600值为0.6~0.8(取出1 m L 作为未诱导样品)加入IPTG(终浓度为1 mmol/L),分别在诱导2,4,6,8 h时,取300μL 菌液,进行SDS-PAGE凝胶电泳鉴定。
将菌液蛋白经SDS-PAGE 凝胶分离后,在15 V,2 h的条件下转至硝酸纤维膜(NC)上,用5%的脱脂奶过夜封闭;使用PBST 以1∶1 000稀释的鼠抗His的单抗37℃孵育1 h后,PBST 洗NC膜3次,然后使用PBST 以1∶5 000稀释的HRP 标记的羊抗鼠IgG(H+L)为二抗,37℃孵育1 h;经PBST 洗3次,去离子水洗3次后,进行增强化学发光(ECL)显色反应曝光。
1.2.6 重组蛋白的可溶性分析 将2瓶100 m L菌液,在37℃、220 r/min摇菌1.5 h后,加入终浓度为1 mmol/L的IPTG 分别在37℃或16℃的摇床上诱导表达8 h和18 h后,分别转入100 m L离心管中,4℃、8 000 r/min离心5 min;弃上清液,加入10 m L PBS(p H 为7.4),在超声波破碎仪下超声20 min(工作5 s,间歇5 s),使得液体均一透明,然后4℃、8 000 r/min 离心15 min,上清液与沉淀分别进行SDS-PAGE鉴定。
1.2.7 包涵体蛋白的变性与复性 包涵体变性过程:将200 m L 诱 导 菌 液,经8 000 r/min 离 心20 min后弃上清,加入20 m L 裂解缓冲液重悬沉淀并混匀,超声破碎10 min,8 000 r/min离心20 min;弃上清液,加入洗涤缓冲液重悬沉淀后超声破碎2 min,8 000 r/min 离心15 min;弃上清液,再用2 mol/L 尿素洗涤液重悬沉淀后超声破碎2 min,8 000 r/min 离心15 min;弃去上清,加入10 m L 8 mol/L 尿素变性液变性包涵体,冰上放置30 min后,8 000 r/min离心15 min;收集上清,即为变性包涵体,用紫外分光光度计测定蛋白浓度。
包涵体稀释复性过程:使用稀释液(Tris-HCl 0.02 mol/L,EDTA ·2Na 0.002 mol/L,尿 素8 mol/L,GSH 1 mmol/L,GSSG 0.1 mmol/L,p H为8.0)调整变性包涵体的浓度,使蛋白终质量浓度约为100 mg/L,尿素浓度为1 mol/L 以下,然后溶液4℃静置20 h;经浓缩管浓缩至蛋白质量浓度约为400 mg/L时,将浓缩的蛋白液装入透析袋;将透析袋放入透析液(20 mmol/L Tris-HCl,p H 为8.0)中,4℃磁力搅拌透析,每2 h换1次透析液,换液6次,收集透析袋中的蛋白,蛋白经0.45μm 的滤膜过滤,使用Bradford 法(PC0010)测定蛋白浓度后,-80℃保存。
1.2.8 重组蛋白的活性检测 用0.05 mol/L CBS将复性重组蛋白稀释为5 mg/L,每孔100μL 包被酶标板,4℃过夜;用250μL/孔的5%脱脂奶封闭酶标板,37℃孵育1 h后,再用100μL 不同稀释梯度的PRRSV 阳性血清和阴性血清于37℃孵育1 h;使用PBST 以1∶1 000稀释的HRP-羊抗猪IgG 抗体100μL加入到各孔中,37℃孵育1 h;PBST 洗涤6次后,甩干,加入100μL 显色液,5 min后加50μL 10.85%浓硫酸终止,15 min内使用酶标仪读数。
1.2.9 免疫后小鼠的血清效价检测 以50μg蛋白或蛋白稀释液(20 mmol/L Tris-HCl)与弗氏完全佐剂1∶1混合,首免6~8周龄BALB/c鼠,每间隔2周进行二免、三免(50μg蛋白或蛋白稀释液与弗氏不完全佐剂1∶1混合),三免后小鼠剪尾采血,用100μL 不同稀释梯度的免疫血清作一抗,使用PBST 以1∶5 000稀释的HRP标记的羊抗鼠IgG(H+L)为二抗。具体步骤与1.2.8相同。
2 结果
2.1 目的基因的扩增以真核质粒pcDNA3.1-ORF2为模板,经PCR 扩增GP2a胞外区,目的基因大小为523 bp,与预期基因大小一致(图1)。
2.2 重组质粒p ET-32a-eORF2的菌液PCR 与双酶切鉴定将GP2a胞外区连接到p ET-32a构建重组质粒p ET-32a-eORF2,挑选单克隆菌进行质粒PCR,PCR 结果如图2所示,然后提取重组质粒,重组质粒经Bam HⅠ与SalⅠ双酶切,切出大小约为5 900 bp 的质粒载体pET-32a和大小约为520 bp的目的基因(图3)。
2.3 重组蛋白的表达及Western blot鉴定重组菌种p ET-32a-eORF2-Rosetta(DE3)在37℃转速为220 r/min的摇床中诱导培养不同时间后,SDSPAGE结果表明GP2a胞外区成功表达,重组蛋白相对分子质量大小约36 000,且在诱导8 h时,表达量最多(图4)。Western blot鉴定显示未诱导的p ET-32a-eORF2-Rosetta(DE3)未显色,而诱导的p ET-32a-eORF2-Rosetta(DE3)在 约36 000 处 显色,对照p ET-32a-Rosetta(DE3)在约20 000 处显色,表明GP2a胞外区被成功表达(图5)。

图1 pcDNA3.1-ORF2 的 PCR 结 果 M.DL2000 DNA
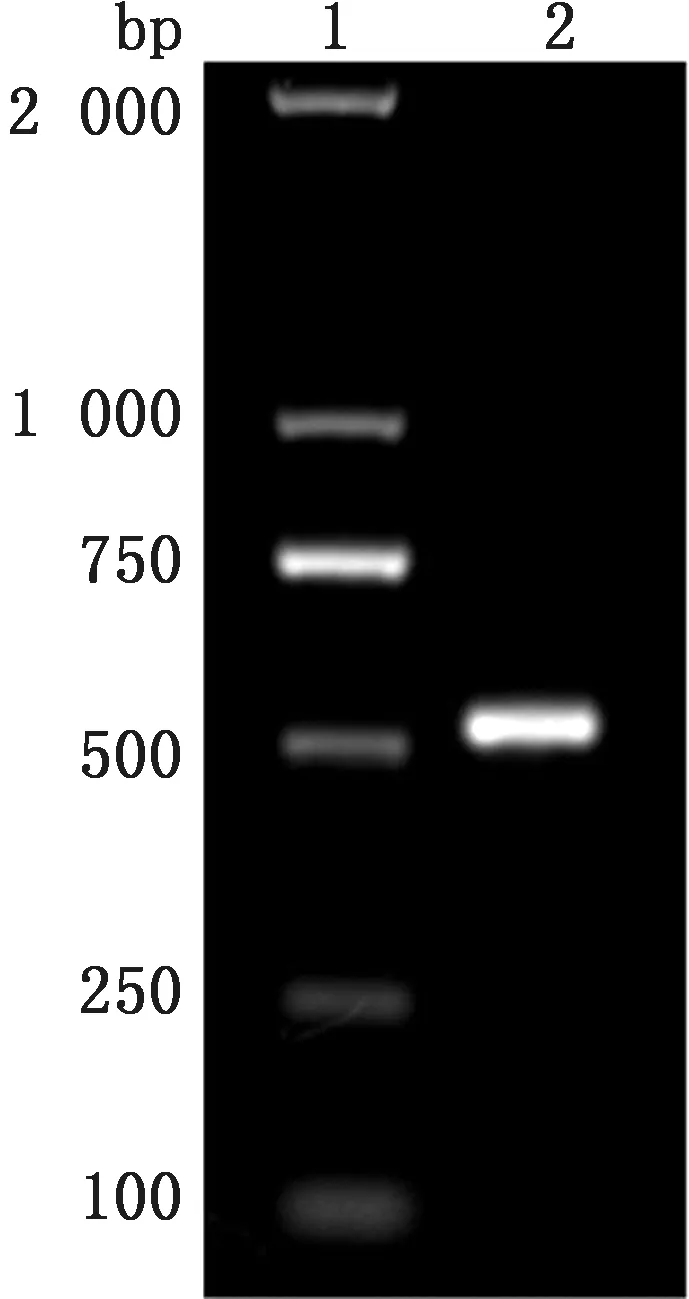
图2 重组质粒p ET-32a-eORF2的PCR 结果 M.DL2000 DNA Marker;1.PCR 扩增产物
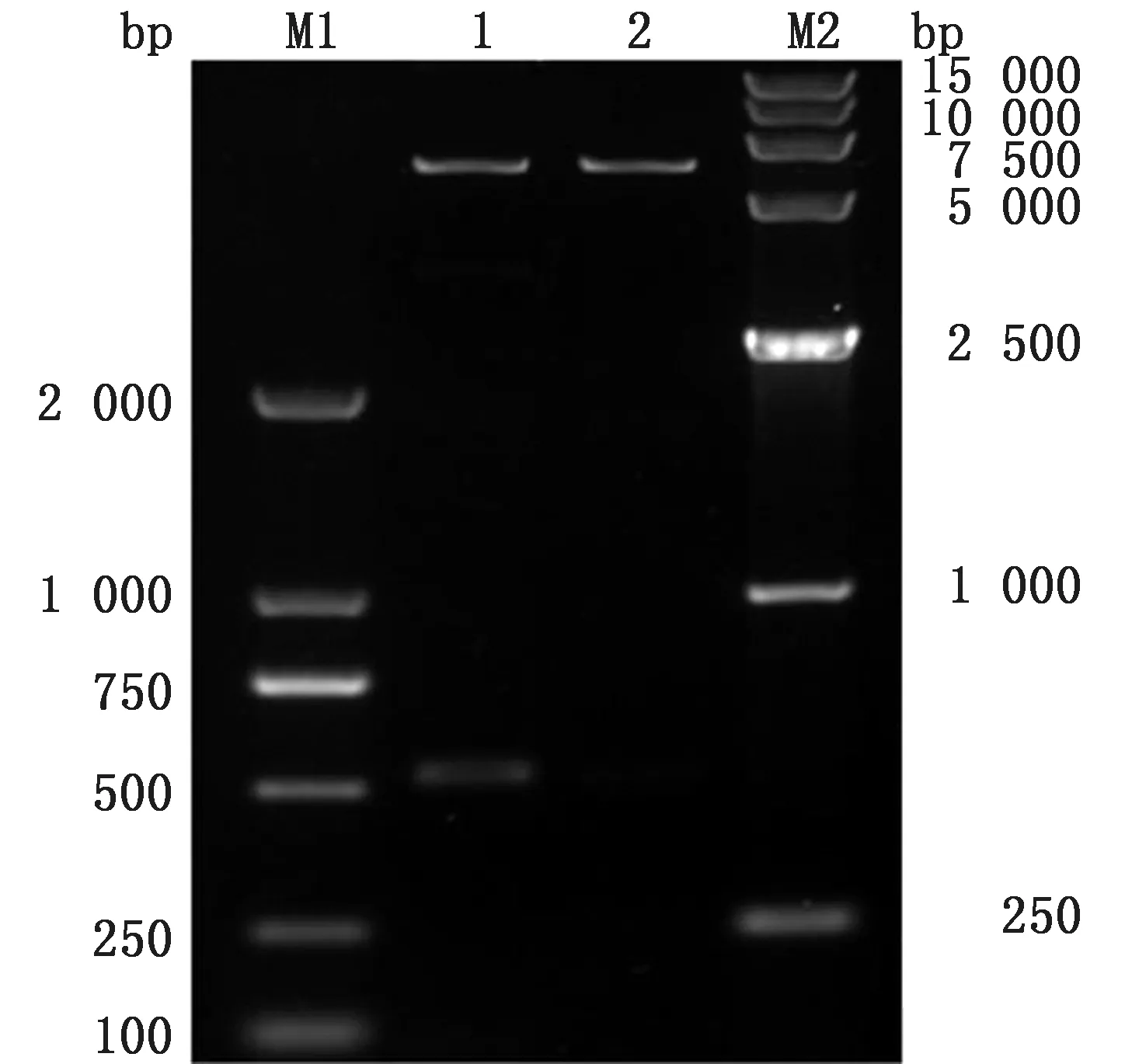
图3 重组质粒p ET-32a-eORF2双酶切鉴定 M1.DL2000 DNA Marker;1.p ET-32a-eORF2 酶 切 产 物;2.p ET-32a酶切产物;M2.DL15000 DNA Marker
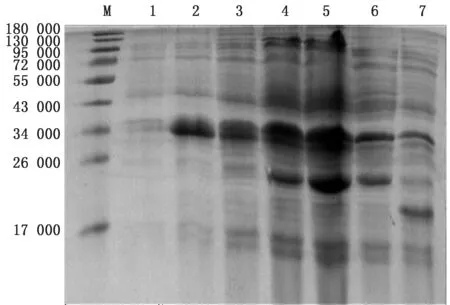
图4 重组蛋白的SDS-PAGE 分析 M.蛋白Marker;1.p ET-32a-eORF2-Rosetta(DE3)未诱导产物;2~5.p ET-32a-eORF2-Rosetta(DE3)分别为诱导2,4,6,8 h 产 物;6.p ET-32a-Rosetta(DE3)的 未 诱 导 产 物;7.p ET-32a-Rosetta(DE3)的诱导产物
2.4 重组蛋白的可溶性分析重组菌种p ET-32aeORF2-Rosetta(DE3)分别在37,16℃诱导8,18 h,经SDS-PAGE分析,表明重组的GP2a胞外区蛋白主要以包涵体的形式表达(图6)。
2.5 重组蛋白的变性与复性经尿素变性和稀释复性,得到蛋白纯度达到90%以上的变性重组蛋白和复性重组蛋白,SDS-PAGE 分析结果如图7和图8所示。
2.6 Bradford法测定蛋白浓度将透析后的蛋白进行蛋白质量浓度测定,D570值为0.273。根据试剂盒说明书,测定的牛血清白蛋白(bovine albumin,BSA)蛋白标准曲线如图9,计算得出透析后蛋白质的质量浓度为0.23 g/L。
2.7 重组蛋白的活性检测结果见表1。ELISA结果显示重组蛋白与猪PRRSV 阳性血清反应的D450值高于与猪PRRSV 阴性血清,而且阳性血清值与阴性值之比(P/N)大于2.1,表明重组蛋白能够被PRRSV 阳性血清所识别,具有一定免疫原性。
2.8 免疫后小鼠的血清效价检测用重组蛋白三免BALB/c小鼠2周后,断尾采血,间接ELISA 检测血清效价,当阳性血清稀释到1∶102 400 时,D450值接近于0.5,且阳性与阴性比值均大于2.1,这说明抗体效价可达到1∶102 400(表2)。
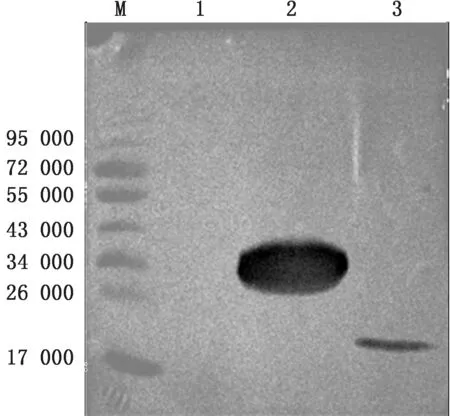
图5 重组蛋白的Western blot鉴定 M.蛋白Marker;1.p ET-32a-eORF2-Rosetta(DE3)的未诱导产物;2.p ET-32a-eORF2-Rosetta(DE3)的诱导表达产物;3.p ET-32a-Rosetta(DE3)的诱导表达产物
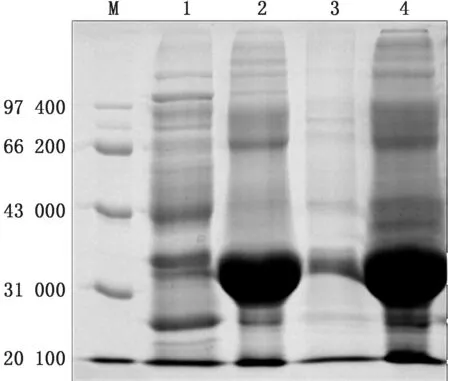
图6 重组蛋白的可溶性分析 M.蛋白Marker;1.37℃诱导上清液;2.37℃诱导沉淀;3.16℃诱导上清液;4.16℃诱导沉淀
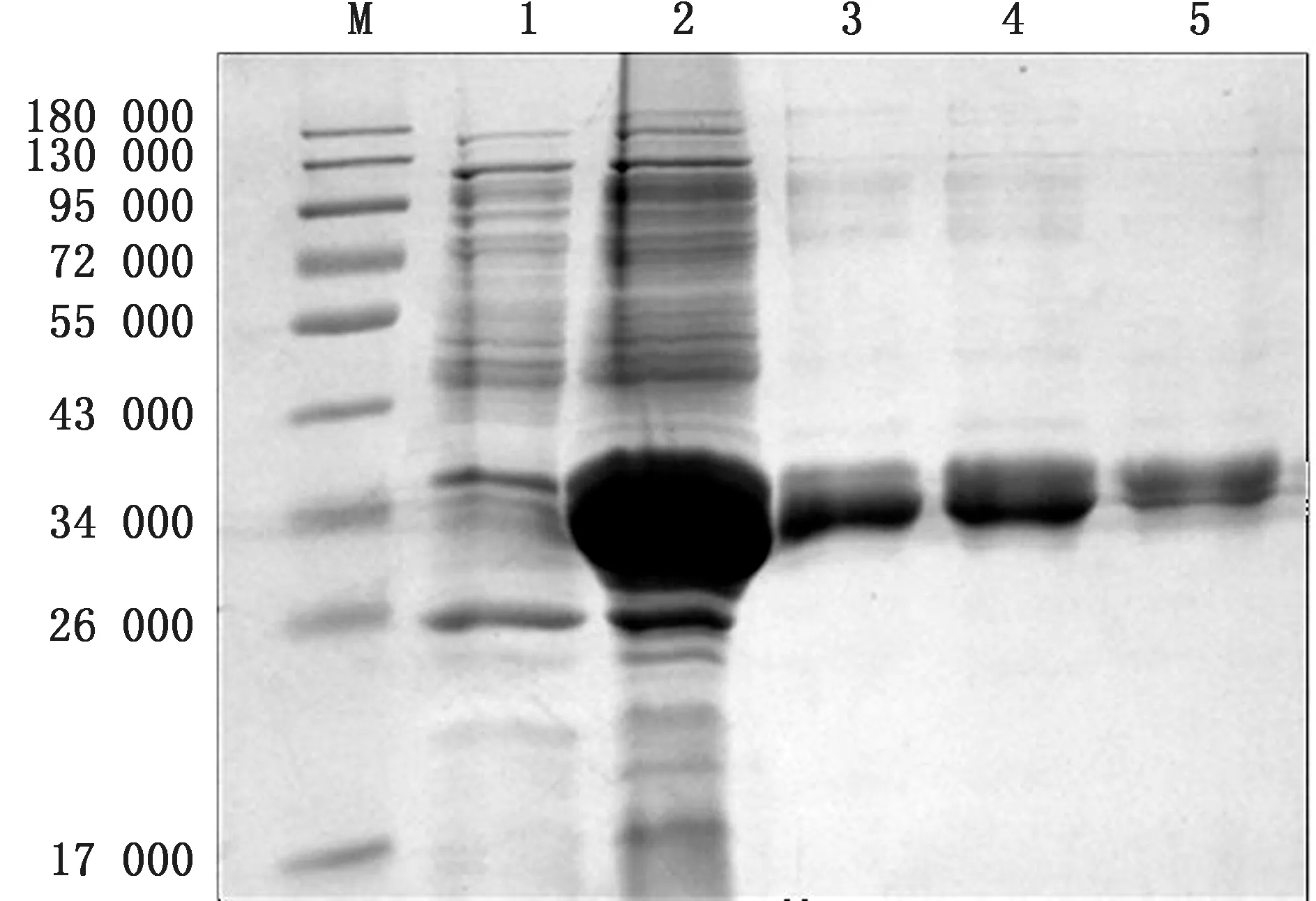
图7 重组蛋白包涵体的洗涤和变性 M.蛋白Marker;1.裂 解 上 清 液;2.裂 解 沉 淀;3.洗 涤 沉 淀;4.2 mol/L 尿素沉淀;5.8 mol/L尿素变性蛋白
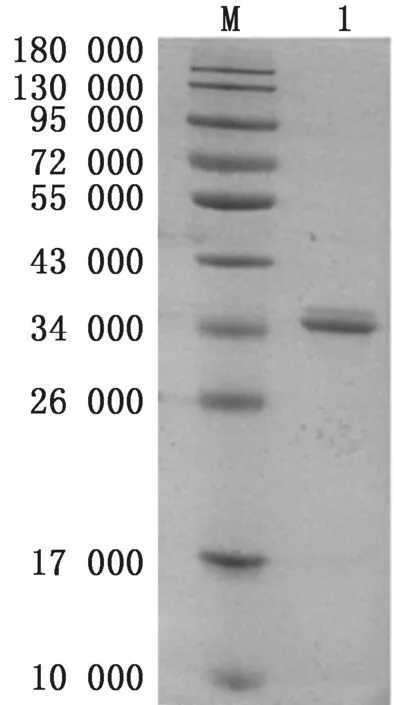
图8 复性后重组蛋白的SDS-PAGE分析 M.蛋白Marker;1.复性后重组蛋白
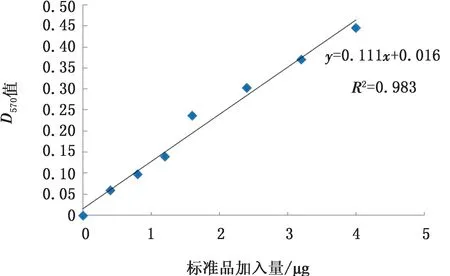
图9 Bradford BSA 蛋白标准曲线
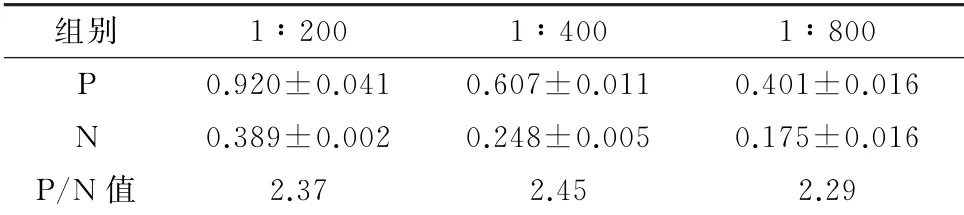
表1 猪阳性血清的ELISA 检测结果 D 450值
3 讨论
本试验探讨了PRRSV BJ-4毒株GP2a胞外区的原核表达,并对其免疫原性加以研究。由于GP2a蛋白N 端(信号肽序列)与C端均具有跨膜区,所以原核表达系统不易表达该蛋白或表达的重组蛋白以包涵体的形式表达。因此,本研究选择p ET-32a为载体,成功构建了GP2a胞外区的重组质粒p ET-32a-eORF2,并在大肠杆菌Rosetta(DE3)中得到表达,表达量随着时间的延长而增多。可溶性分析结果显示,使用带有较大可溶性标签的p ET-32a作为表达载体,有很小部分重组蛋白以可溶的形式表达,但由于可溶蛋白含量低且重组蛋白p H 偏酸(p H 为6.76),未能通过镍柱亲和层析法纯化出单一可溶的重组蛋白。
包涵体蛋白具有防止蛋白酶水解与产率大的优点,但会发生缺乏生物学活性的缺点[10],所以要对变性的包涵体进行复性。本试验选择稀释复性的方法对包涵体蛋白复性,在稀释液中加入氧化型与还原型谷胱甘肽,辅助蛋白质恢复正常的折叠状态;由于稀释复性使得蛋白质浓度降低,为了保证复性后的蛋白质浓度,本试验使用浓缩管对稀释后的蛋白进行反复浓缩,从而达到试验所需的浓度,这避免了在透析复性过程中因蛋白浓度过大造成的蛋白聚沉。经PRRSV 的猪阳性血清和重组蛋白免疫后的小鼠血清的ELISA 检测,表明复性后的重组蛋白具有良好的免疫活性。
目前,PRRSV 感染猪只后,已报道了N 蛋白、M 蛋白、GP5、NSP1、NSP2、NSP4、NSP7、NSP8 的抗体动态变化[11-12],而针对GP2a蛋白的抗体动态水平还不清楚。本研究表达的GP2a胞外区蛋白,可作为检测原,检测GP2a抗体的动态变化;同时,为深入研究PRRSV GP2a蛋白的相关生物学功能奠定基础。
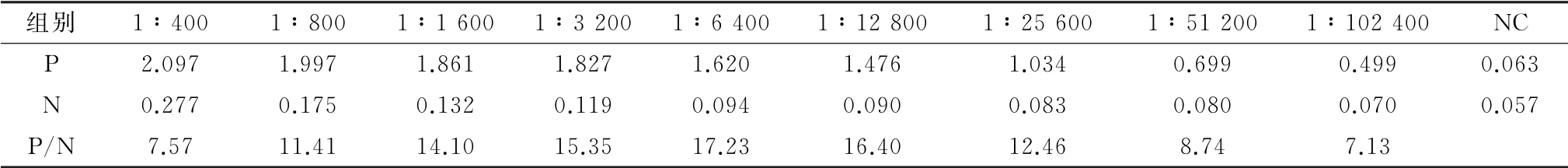
表2 免疫小鼠血清的ELISA 检测结果 D 450值
猜你喜欢
中国造纸(2022年9期)2022-11-25 02:24:54
环球时报(2019-04-03)2019-04-03 04:15:14
食品科学(2018年10期)2018-05-23 01:27:28
幸福·健康版(2016年10期)2016-11-17 11:21:46
西藏科技(2015年10期)2015-09-26 12:10:16
西南医科大学学报(2015年1期)2015-08-22 13:01:46
实用手外科杂志(2015年3期)2015-04-03 12:19:54
中国当代医药(2015年9期)2015-03-01 02:01:59
西南军医(2015年6期)2015-01-23 01:25:50
湖北大学学报(哲学社会科学版)(2014年4期)2014-03-20 13:56:45
