
沼泽红假单胞菌作为微生物细胞工厂的应用
2020-03-12 02:51李美洁夏青青HARWOOD杨建明
生物加工过程 2020年1期
李美洁,夏青青,HARWOOD C S,2,杨建明
(1.青岛农业大学生命科学学院光合固碳产能中心,山东青岛266109;2.华盛顿大学微生物学系,美国西雅图98195-7240)
沼泽红假单胞菌(Rhodopseudomonaspalustris)是一种紫色非硫细菌(purple non-sulfur bacterium,PNSB),属于变形菌门、红螺菌科、红假单胞菌属[1]。沼泽红假单胞菌在自然界中分布很广泛,主要分布在光照充足的厌氧水环境中,在湖泊、土壤、沼泽、大海等均有分布[2]。沼泽红假单胞菌新陈代谢方式多样,可以通过自然界存在的4种代谢模式进行生长,包括光合自养、光合异养、化学自养和化学异养(图1)[3]。

图1 沼泽红假单胞菌的4种新陈代谢模式[3]Fig.1 The four types metabolism of R.palustris[3]
目前,沼泽红假单胞菌主要应用于废水处理和水产养殖领域(图2)。沼泽红假单胞菌富含生物活性蛋白质、维生素、多糖、泛酸、叶酸等,可作为饲料添加剂[4];沼泽红假单胞菌能够提高养殖水的含氧量,稳定pH,净化养殖水环境[5];沼泽红假单胞菌还能够提高水生生物的免疫能力,预防疾病。而且,沼泽红假单胞菌具有很强的生物降解性能,能够降解动植物废弃物中和工业废弃物中的多种成分,包括木质素单体、含氮化合物、氯化物、芳香族化合物等[6]。对沼泽红假单胞菌的全基因组进行测序,发现其基因组中含有大量的降解基因[3]。总之,在废水处理和水产养殖行业,沼泽红假单胞菌有很大的研究价值,也是目前研究的重点。

图2 沼泽红假单胞菌的应用Fig.2 Application of Rhodopseudomonas palustris
除了以上应用,近年来,沼泽红假单胞菌在其他方面也有应用,包括产氢气(H2)[7]、产甲烷(CH4)[8]等燃气和燃料,产类胡萝卜素等萜烯化合物[9],在微生物燃料电池(MFC)[10]、微生物电合成[11]以及光催化合成[12]等领域(图2)。
沼泽红假单胞菌表现出了构建微生物细胞工厂的巨大潜力和应用价值。本文中,笔者通过3个方面对其作为微生物细胞工厂的研究应用进行探讨:碳源多样性;基因工程策略;沼泽红假单胞菌作为微生物细胞工厂的应用领域。另外,对其目前研究面临的问题和进一步的研究方向进行展望。
1 碳源多样性
不同于其他常用的微生物细胞工厂,沼泽红假单胞菌能够利用存在于废物废气中的碳源,包括有小分子机酸、醇类、无机盐和芳香族化合物等(表1)。沼泽红假单胞菌以光作为能量来源,产ATP。因此,作为微生物细胞工厂,沼泽红假单胞菌在能量来源和碳源方面具有很大的优势。
目前,已有多种碳源被报道可以用于沼泽红假单胞菌的生长。首先,沼泽红假单胞菌可以分解利用农业和工业废水中大量存在的小分子有机酸,包括乙酸、丁酸和乳酸等,在进行废水处理的同时生产目标产物。在不同有机酸条件下,沼泽红假单胞菌的生长不同,产目标产物的能力不同。在产CH4的沼泽红假单胞菌中,在培养基中分别加入7种不同的有机酸盐,选取延胡索酸时,其CH4产量最高[8]。碳源的不同浓度也能够影响目标产物的产量。在产H2的沼泽红假单胞菌培养基中,加入不同浓度(2~10 mmol/L)的乳酸,H2的产量不同[13]。
其次,沼泽红假单胞菌还可以利用乙醇、粗甘油、丁醇等醇类物质作为碳源。粗甘油是生物柴油生产过程中主要的副产物,因此一般作为工业废料进行处理。基于环境和经济效益的考虑,如何利用粗甘油是工业生产上的突破点。研究表明,沼泽红假单胞菌可以利用粗甘油作为碳源合成H2[14]。
再次,沼泽红假单胞菌还可以利用无机盐作为碳源。在产甲烷的沼泽红假单胞菌中,以无机盐NaHCO3作为碳源,在碳酸酐酶的作用下分解为CO2,硫代硫酸盐提供电子,部分CO2通过卡尔文-本森-巴沙姆循环(CBB cycle)参与细胞生长代谢,部分CO2与电子在固氮酶的作用下合成CH4[8]。
最后,沼泽红假单胞菌还可以利用多种芳香族化合物作为单独的碳源。以木质纤维素水解产物作为微生物细胞生长的能量来源是现在研究的热点,但是水解产物中存在大量的芳香族化合物,能够抑制微生物细胞的生长。在沼泽红假单胞菌中,存在苯甲酰辅酶A途径(benzoyl-CoA pathway),这个途径可以分解代谢多种芳香族化合物[15-17]。目前对芳香族化合物库玛酸盐的研究比较多,库玛酸是一种木质素单体,以它作为碳源时,沼泽红假单胞菌的固氮效率和H2产率都比较高,是乙酸为碳源时的2~5倍[18]。

表1 沼泽红假单胞菌可利用的碳源
综上所述,由于沼泽红假单胞菌可利用的碳源种类比较广泛,而且很多碳源是工业和农业生产上废水废气处理中比较难以去除的化合物,因此从碳源利用的角度来说,沼泽红假单胞菌作为微生物细胞工厂具有很大的优势。
2 基因工程策略
沼泽红假单胞菌作为微生物细胞工厂,通过基因工程策略,对其进行基因敲除、基因表达等是非常必要的。但是,目前对沼泽红假单胞菌的基因编辑工具研究得并不多。为了方便基因操作,Inui等[25]从400个菌中筛选获得能够在紫色非硫细菌中复制的质粒pMG101(15 kb),研究发现:pMG101来源于其中的沼泽红假单胞菌,以pMG101为基础,获得了沼泽红假单胞菌-大肠杆菌穿梭质粒pMG103(5.68 kb)和pMG105(5.68 kb);在非选择压力条件下,这些质粒能够在沼泽红假单胞菌中稳定地复制传代[25]。Xu等[26]以质粒pMG103作为载体,在沼泽红假单胞菌中过表达crtE、hpnD和dxs基因,以提高角鲨烯产量至野生型菌株的112倍,达到15.8 mg/g。
另外一种宿主广泛的穿梭质粒pBBR1 MCS-5在沼泽红假单胞菌能够用于载体构建和外源基因表达。Huang等[27]将fix基因簇克隆至pBBR1 MCS-5质粒上的多克隆位点,将重组质粒转化沼泽红假单胞菌,使得fix基因簇外源表达,除了pBBR1 MCS-5外,pBBR1 MCS系列的pBBR1 MCS-1、pBBR1 MCS-2、pBBR1 MCS-3、pBBR1 MCS-4和pBBR1 MCS_START等都能应用于沼泽红假单胞菌,可以根据实际需要进行选择[28]。
在沼泽红假单胞菌中,除了基因表达外,通过自杀质粒进行基因敲除、基因替换的研究也有报道。pJQ200KS是一种在革兰氏阴性菌中应用很广泛的自杀质粒,含有P15A进行质粒复制,通过庆大霉素抗性进行筛选,含有来自于枯草芽孢杆菌的sacB基因(编码蔗糖-6-果糖基转移酶),能够催化蔗糖水解合成高分子果聚糖,对革兰氏阴性菌产生致死作用[29]。沼泽红假单胞菌是一种革兰氏阴性菌,Rey等[18]利用自杀质粒pJQ200KS将突变的nifA*571和nifA*574片段转入沼泽红假单胞菌的染色体上,以研究固氮酶的产H2功能。另外,也通过pJQ200KS将突变的nifDV75AH201Q转入染色体上,以研究固氮酶产甲烷的功能[8]。
总之,目前应用于沼泽红假单胞菌的基因编辑工具很少,为了将沼泽红假单胞菌更好地应用于微生物细胞工厂的构建,研究探索其基因编辑工具是非常有必要的,也是非常迫切的。
3 沼泽红假单胞菌作为微生物细胞工厂的应用
3.1 利用沼泽红假单胞菌产燃气燃料
目前,人类社会对石油、煤炭等化石燃料具有很大依赖性,化石能源不可再生,而且其生产应用过程引起很大的环境污染,寻找清洁能源迫在眉睫。
沼泽红假单胞菌的产H2能力被广泛地研究。在沼泽红假单胞菌中,固氮酶能够将N2转化为NH3,同时产生H2,而且在没有N2时,固氮酶也能催化H2的合成,见式(1)[7]。其中,H2的合成过程:有机物或者无机物底物提供电子,通过光合电子传递链传递给铁氧还原蛋白,光能通过光合电子传递链转化为ATP,ATP和带电子的铁氧还原蛋白在固氮酶的作用下产生H2[7]。与藻类不同的是,沼泽红假单胞菌的光合系统无法分解H2O,因此不产生O2,更利于H2的生成,这种光合细菌产H2速率更快[7,30]。
天然的固氮酶将75%的铁氧还原蛋白用于N2的固定,合成H2效率较低[18]。而且,另外一个产物NH3能够在转录水平上和翻译后水平上抑制固氮酶的活性[31]。Rey等[18]通过筛选获得了突变菌株,使固氮酶NifA亚基的转录调控因子发生点突变,从而减弱NH3对固氮酶活性的抑制,最终提高H2产量。目前对于沼泽红假单胞菌产H2的研究主要集中于不同光照强度、不同碳源等的考察,而在基因水平上提高其产量却鲜有报道,这可能是因为对其产H2的代谢途径不清楚所导致的。
钼依赖型固氮酶,其反应为

(1)
钼依赖型固氮酶突变体,其反应为
(2)
铁依赖型固氮,其反应为
(3)
近年来,沼泽红假单胞菌的产甲烷性能也被报道。Harwood团队的Lixen等[8]发现钼依赖性的固氮酶突变体(NifDV75AH201Q)能够催化CO2合成CH4,同时合成H2,替换其原有的固氮功能见式(2)。另外,钒依赖性的固氮酶发生同样的突变之后(NifDV57AH180Q)也能够催化CH4的合成[21]。而天然的铁依赖性固氮酶同时具有合成CH4的能力和固氮的能力,可以同时合成CH4、NH3和H2,但是CH4的比例比较低见式(3)[21]。如何通过代谢工程和合成生物学手段提高固氮酶合成CH4的能力是进一步研究的方向。
另外,以沼泽红假单胞菌作为底盘细胞产丁醇也有报道,与乙醇相比,丁醇是一种高密度、不易挥发、与传统燃料不相溶的燃料。在沼泽红假单胞菌中通过质粒过表达来自于丙酮丁醇梭菌(Clostridiumacetobutylicum)的adhE2基因,能够催化底物丁酸盐合成丁醇,其产率为0.03 g/(L·d),转化率达到了40%,接近于其理论转化率45%[32]。但是由于沼泽红假单胞菌生长代谢比较慢,影响其丁醇的产率,如何提高其生长代谢是研究的重点和难点。
3.2 利用沼泽红假单胞菌产萜烯化合物
沼泽红假单胞菌富含不同种类的萜烯化合物,其中类胡萝卜素含量很高。类胡萝卜素在细胞膜上通过非共价键连接到跨膜蛋白上,一方面能够稳定跨膜蛋白,另一方面能够捕获光线,具有光保护作用[33]。番茄红素和β-胡萝卜素等类胡萝卜素在食品和药品等方面具有很大的应用价值,目前主要是从番茄等植物中提取。利用沼泽红假单胞菌提取番茄红素和胡萝卜素是一种替代方法。沼泽红假单胞菌细胞结构简单,提取相对容易,因此作为类胡萝卜素的细胞工厂其性能优于植物和微藻等宿主。目前针对沼泽红假单胞菌产类胡萝卜素的研究,主要是在发酵水平改变培养条件,提高类胡萝卜素产量这些方面。崔艺久等[9]对产类胡萝卜素的发酵条件(碳源、氮源和pH等)进行优化,提高了类胡萝卜素的累积量。Kuo等[34]以及Muzziotti等[35]研究发现,在不同的光照强度下,类胡萝卜素的含量和种类是不同的。而且在沼泽红假单胞菌的类胡萝卜素中,番茄红素含量最高,在一定的光照条件下,可以达到类胡萝卜素总量的79%。因此,沼泽红假单胞菌作为宿主产番茄红素是非常有潜力的。但是,目前从基因水平对其代谢途径进行改造优化还没有研究报道。通过沼泽红假单胞菌的全基因组测序结果,找到了其代谢途径——2-C-甲基-D-赤藓糖醇-4-磷酸盐(MEP)途径的基因和下游合成类胡萝卜素的基因(图3)[3]。对沼泽红假单胞菌在基因水平上进行研究,将能够进一步提高其产量。

图3 沼泽红假单胞菌基因组上类胡萝卜素合成途径基因的分布Fig.3 Genes distribution of the carotenoids synthetic pathway in R.palustris genome
以沼泽红假单胞菌作为微生物细胞工厂,通过代谢工程合成另一种萜烯化合物——角鲨烯,通过敲除下游角鲨烯-藿烯环化酶基因(shc),将代谢途径的2个连续的酶CrtE和HpnD进行融合,过表达上游MEP途径的dxs基因,最终角鲨烯产量达到15.8 mg/g,是野生菌株的112倍[26]。在野生型的沼泽红假单胞菌中,角鲨烯的下游产物藿烷类化合物(hopanoids)的产量能够达到30 mg/g[36]。因此,沼泽红假单胞菌的角鲨烯产量还有进一步的提升潜力。以其他底盘细胞酿酒酵母[37]和聚球藻[38]产角鲨烯的研究结果,将对以沼泽红假单胞菌作为底盘细胞的研究具有指导意义。
以沼泽红假单胞菌作为微生物细胞工厂产其他萜烯化合物的研究目前还没有报道,但是由于其自身产类胡萝卜素和藿烷类化合物的能力比较突出,通过代谢工程和基因工程手段探索其产萜烯化合物能力具有开发前景。
3.3 沼泽红假单胞菌在微生物燃料电池的应用
MFC是一种特殊的燃料电池,微生物在阳极区能够氧化有机底物,释放电子和质子,电子和质子转移至阴极与O2反应生成H2O,最终将有机物的化学能转化电能(图4)[39]。当利用光合微生物应用于MFC时,还可以同时将光能转化为电能(图4(a))[40]。目前,MFC主要应用于废水处理,以废水中大量的有机物为底物,在分解有机物的同时产生电能。Xing等[41]研究发现,沼泽红假单胞菌(R.palustrisDX-1)能够用作MFC,以乙酸盐作为底物时,其功率密度能够达到(2 720±60) mW/m2,高于混合菌的功率密度。另外,Lai等[10]研究发现,沼泽红假单胞菌的多磷酸盐代谢与其产电能力相关:在光照条件下,沼泽红假单胞菌在停滞生长期能够累积多磷酸盐,通过多磷酸盐储存能量;在生长条件受限制的情况下,沼泽红假单胞菌能够利用储存的多磷酸盐维持生长,同时产生电能。由此可见,沼泽红假单胞菌应用于MFC是非常有潜力的。在MFC领域,目前的研究主要通过电极材料和反应器等方面的优化来提高其污水处理效率和产电能力[42],而对产电微生物产电能力的研究还比较少。在基因水平上对沼泽红假单胞菌等产电微生物进行改造,提高其产电能力是进一步研究的方向。
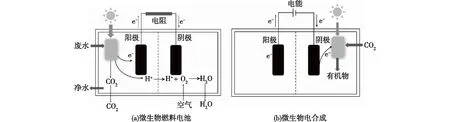
图4 以光合微生物(沼泽红假单胞菌)构建的微生物燃料电池和微生物电合成示意Fig.4 Illustration of the microbial fuel cell and microbial electrosynthesis using R.palustris
3.4 沼泽红假单胞菌在微生物电合成的应用
沼泽红假单胞菌与固体电极能够发生双向电子转移,除了能够向阳极释放电子应用于微生物燃料电池领域,还能够从阴极吸收电子,将CO2等底物固定,合成有机物,应用于微生物电合成领域(MES)(图4(b))。Rengasamy等[11]利用沼泽红假单胞菌(R.palustrisTIE-1)以铁基氧化还原介质Prussian Blue作为阴极材料提供电子、CO2作为碳源、合成聚羟基丁酸酯(PHB),一种可降解生物塑料。Guzman等[43]对沼泽红假单胞菌的MES电子传递机制进行研究发现,电子从阴极传递至光合系统的电子传递链,从而参与沼泽红假单胞菌的体内生物合成途径(例如CO2的固定)。除了PHB的合成,其他常见的生物基化学品,例如乙酸、1,3-丙二醇等都可以通过微生物电合成。但是,目前微生物电合成属于新型研究方向,沼泽红假单胞菌应用于微生物电合成也是近几年才开始研究的领域,电子传递效率是制约其产物合成的关键因素,如何提高胞外胞内电子传递,如何改造沼泽红假单胞菌提高其代谢流,是今后需要研究解决的问题。
3.5 沼泽红假单胞菌合成纳米材料及其在光催化合成领域的应用
沼泽红假单胞菌能够将电能转化为目标产物,即微生物电合成,还能够不通过其光合传递系统而将太阳能直接转化为目标产物,实现光催化合成。附着于细胞表面的或者细胞内的纳米颗粒硫化镉(CdS)能够吸收太阳能,释放电子,参与细胞内各种生物合成途径(图5)。Wang等[12]研究发现,Cd2+和半胱氨酸的S2-在沼泽红假单胞菌表面形成的CdS纳米颗粒,在太阳光的刺激下释放电子,产生还原力,产生ATP,参与固定CO2的CBB循环,合成甘油醛3-磷酸,参与体内生物合成。在CdS纳米颗粒的作用下,沼泽红假单胞菌的生物量、类胡萝卜素产量和PHB产量分别提高至148%、122%和147%[12]。CdS纳米颗粒还可以在沼泽红假单胞菌胞内合成,吸收太阳能,参与固氮酶的N2固定作用[44]。目前,将沼泽红假单胞菌与纳米颗粒结合进行光催化合成还在发展的起步阶段,对不同的纳米材料进行研究,针对目标产物对沼泽红假单胞菌进行基因改造,有望提高其催化合成效率,最终实现工业化生产。

图5 沼泽红假单胞菌与纳米材料构建光催化合成系统Fig.5 Illustration of the photocatalytic CdS R.palustris system
3.6 以沼泽红假单胞菌构建微生物细胞工厂存在的问题以及解决方法
虽然沼泽红假单胞菌作为微生物细胞工厂应用的潜力很大,但是目前还处于发展的初级阶段,有一些问题还需要解决。首先,沼泽红假单胞菌利用多种碳源的代谢效率比较低,生长缓慢,影响产物的合成。通过随机突变筛选和定向驯化等策略有望提高其代谢效率。其次,目前由于对其在基因水平的研究较少,可利用的基因操作工具并不多。但是作为一种紫色非硫细菌,很多适用于其他紫色非硫细菌的基因操作工具也适用于沼泽红假单胞菌。再次,一些新型的基因操作工具(例如CRISPR-Cas、反义RNA等),也为沼泽红假单胞菌的基因编辑提供了便利。因此,随着基因操作工具研究应用的增多,获得大量适合于沼泽红假单胞菌的基因操作工具指日可待。最后,目前对其产燃气燃料、产萜烯化合物和产电的研究集中在发酵水平,如改变其光照、碳源、氮源等培养条件或工艺的研究,然而在基因水平的研究比较少。通过代谢工程和合成生物学手段,对其合成目标产物的代谢途径进行改造,通过提高代谢流最终提高产量,是进一步研究的方向。
4 结论和展望
以沼泽红假单胞菌为研究对象,对其在构建微生物细胞工厂方面的应用研究进行了总结,它可以利用多种碳源,可通过多种基因工程策略进行改造,同时在产燃气燃料、产萜烯化合物和产电等领域都有重要应用。作为微生物细胞工厂,沼泽红假单胞菌有合成其他高附加值化合物如库玛酸、粘康酸等的潜力,可实现木质素到芳香族化合物的直接合成。总之,沼泽红假单胞菌作为微生物细胞工厂具有很大的研究应用价值,虽然目前发展仍处于初级阶段,但是潜力巨大。
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05
中国土壤与肥料(2021年5期)2021-12-02
昆钢科技(2021年6期)2021-03-09
天津科技(2020年4期)2020-05-09
新农村(浙江)(2019年5期)2019-05-29
下一代英才(酷炫少年)(2017年9期)2017-11-27
剑南文学(2016年11期)2016-08-22
中国酿造(2016年12期)2016-03-01
中国果菜(2016年9期)2016-03-01
时代英语·高三(2014年5期)2014-08-26
