
转TaPK-R1基因小麦的农艺性状评价
2020-03-12 08:38周淼平张增艳姚金保王化敦杨学明余桂红马鸿翔
江苏农业科学 2020年1期
周淼平, 张增艳, 姚金保, 王化敦, 杨学明, 张 鹏, 余桂红, 马鸿翔
(1.江苏省农业科学院粮食作物研究所,江苏南京 210014; 2.中国农业科学院作物科学研究所,北京 10081)
小麦纹枯病是一类重要的世界性小麦病害,主要由禾谷丝核菌等侵染小麦茎秆基部引起,造成病苗死苗、花秆烂茎,严重时产生枯白穗,导致减产,该病害在我国长江中下游麦区和黄淮麦区频繁发生,每年造成数亿元的经济损失[1],培育和应用抗纹枯病小麦品种无疑是防治该病害最经济有效的途径。常规抗病品种的培育需要有抗性稳定的抗源用于配制杂交组合,近30年来,我国研究人员对3 000余份小麦种质材料进行了纹枯病抗性鉴定,但未筛选到免疫和高抗材料[2]。研究发现,小麦对纹枯病的抗性是一数量性状,目前已对部分抗源的抗病数量性状座位(quantitative trait locus,QTL)进行分子定位,但这些QTL对纹枯病抗性的解释率低,要明显提高小麦的纹枯病抗性须累积多个抗性QTL,因而对小麦抗纹枯病的分子标记辅助聚合育种带来诸多困难[3]。
植物基因工程技术为提高小麦的纹枯病抗性提供了新的途径,通过引入外源抗病基因或提高内源抗病基因的表达等方法可以增强小麦对纹枯病菌的抵御能力。目前,采用该方法已经获得一批抗纹枯病的种质材料,进一步拓宽了小麦纹枯病抗源的范围。中国农业科学院作物科学研究所张增艳实验室采用基因芯片分析技术发现了参与小麦抵御纹枯病菌侵染的重要基因TaAGC1(TaPK-R1),该基因属于AGC蛋白激酶,将该基因连接组成型启动子并导入大面积推广品种扬麦20中,发现转基因T2代植株的纹枯病抗性较受体对照扬麦20有显著提高[4-5]。但该基因的组成型表达对小麦其他农艺性状如生育期、产量以及籽粒性状等有无影响尚不清楚。本研究对筛选获得的4个稳定的T7代转基因株系在大田环境下与受体对照扬麦20的农艺性状进行比较,评价TaPK-R1基因的组成型表达对转基因株系生长发育的影响。
1 材料与方法
1.1 材料
1.1.1 植物材料 转TaPK-R1基因株系PK372、PK394、PK465和PK495由中国农业科学院作物科学研究所张增艳研究员提供;受体对照扬麦20由江苏省农业科学院粮食作物研究所麦类作物研究室提供。
1.1.2 试剂与仪器 Karroten植物基因组DNA提取试剂盒,购自南京翼飞雪生物科技有限公司;目的基因PCR检测试剂、RNA提取试剂盒、反转录试剂盒以及荧光定量分析试剂盒,购自宝生物工程(大连)有限公司;PCR引物由生工生物工程(上海)股份有限公司合成。试验所用TP350PCR仪以及TP960荧光定量PCR仪,购自宝生物工程(大连)有限公司。
1.2 试验方法
1.2.1TaPK-R1基因的检测 在拔节期对4个转基因株系和对照扬麦20各取5株叶片,分别混样,冷冻干燥磨碎后参照植物基因组DNA提取试剂盒操作手册提取DNA,导入的TaPK-R1基因采用嵌合引物进行扩增,正向引物序列 PK-3F为5′-CATCTAAGGCGGGTCCAT-3′,反向引物序列NOS-2R为5′-AACCCATCTCATAAATAACG-3′。PCR反应体系总体积为20 μL,含10×buffer 2 μL、25 mmol/L MgCl21.2 μL、2.5 mmol/L dNTPs 1.6 μL、10 μmol/L引物1.0 μL、50 ng模板DNA、1 UTagDNA聚合酶(TaKaRa)。PCR扩增条件为:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火40 s,72 ℃ 延伸45 s,35个循环;72 ℃延伸10 min。PCR产物采用1.0%琼脂糖凝胶电泳分离。
1.2.2TaPK-R1基因的表达分析 在拔节期对4个转基因株系和对照各取5株叶片,分别混样,液氮冷冻磨碎,采用TaKaRa MiniBEST Universal RNA Extraction Kit提取RNA,用PrimeScript RT reagent Kit with gDNA Eraser试剂盒进行反转录,按照SYBR®Premix ExTaqⅡ(Tli RNaseH Plus)试剂盒推荐方法进行荧光定量PCR。Actin基因为内参基因,其特异引物对为TaActin-F(5′-C A C T G G A A T G G T C A A G G C T G-3′)和TaActin-R(5′-C T C C A T G T C A T C C C A G T T G-3′);TaPK-R1基因特异引物对为TaPK-R1-Q-F(5′-T T T G G A G A G G T G C G T C T T T G T-3′)和TaPK-R1-Q-R(5′-A G G A G G T T T C T T T C G G C T T T G-3′)。荧光定量PCR在TaKaRa TP960荧光定量PCR仪上进行,PCR扩增条件为:95 ℃预变性30 s;95 ℃ 变性5 s,60 ℃ 退火30 s,40个循环。根据荧光定量PCR结果计算相对表达量。
1.2.3 转基因株系的农艺性状分析 供试材料于2017年11月28日在江苏省农业科学院六合转基因基地(118°38′47″E、32°28′26″N)播种,对照和每个转基因株系按随机小区种植,每个小区5行,行长 5 m,行距0.25 m,3次重复。播种后按正常小麦管理。
田间试验主要对转基因株系的生长发育和形态特征等主要农艺性状进行观察记载,包括播种期、出苗期、抽穗期、开花期和成熟期等生育期;基本苗、高峰苗、有效穗数、株高、产量等重要农艺性状以及转基因株系对白粉病、纹枯病和赤霉病等病害的抗性。成熟时,考察主穗穗长、小穗数、不孕小穗数以及每穗粒数的变化。收获后,测量籽粒大小、千粒质量、籽粒蛋白含量和籽粒硬度等籽粒性状。籽粒大小测量方法:每小区随机取100粒籽粒,照相后采用SmartGrain软件根据像素计算籽粒长度与宽度;籽粒蛋白含量采用Perten DA7200近红外仪测定;籽粒硬度测量方法:每小区随机取200粒籽粒,采用Perten SKCS4100单粒籽粒硬度测定仪测定,计算平均值。
1.3 数据处理
所有数据的方差分析和检验分析均采用Excel 2016软件进行。
2 结果与分析
2.1 转基因株系TaPK-R1基因的检测结果
此次种植的转基因株系已经是T7代,在拔节期取转基因株系和对照叶片,提取DNA,采用嵌合引物对目的基因TaPK-R1进行PCR检测,结果(图1)表明转基因株系均能扩增出886 bp的目的条带,说明导入的TaPK-R1基因已经整合进小麦基因组并能稳定遗传。
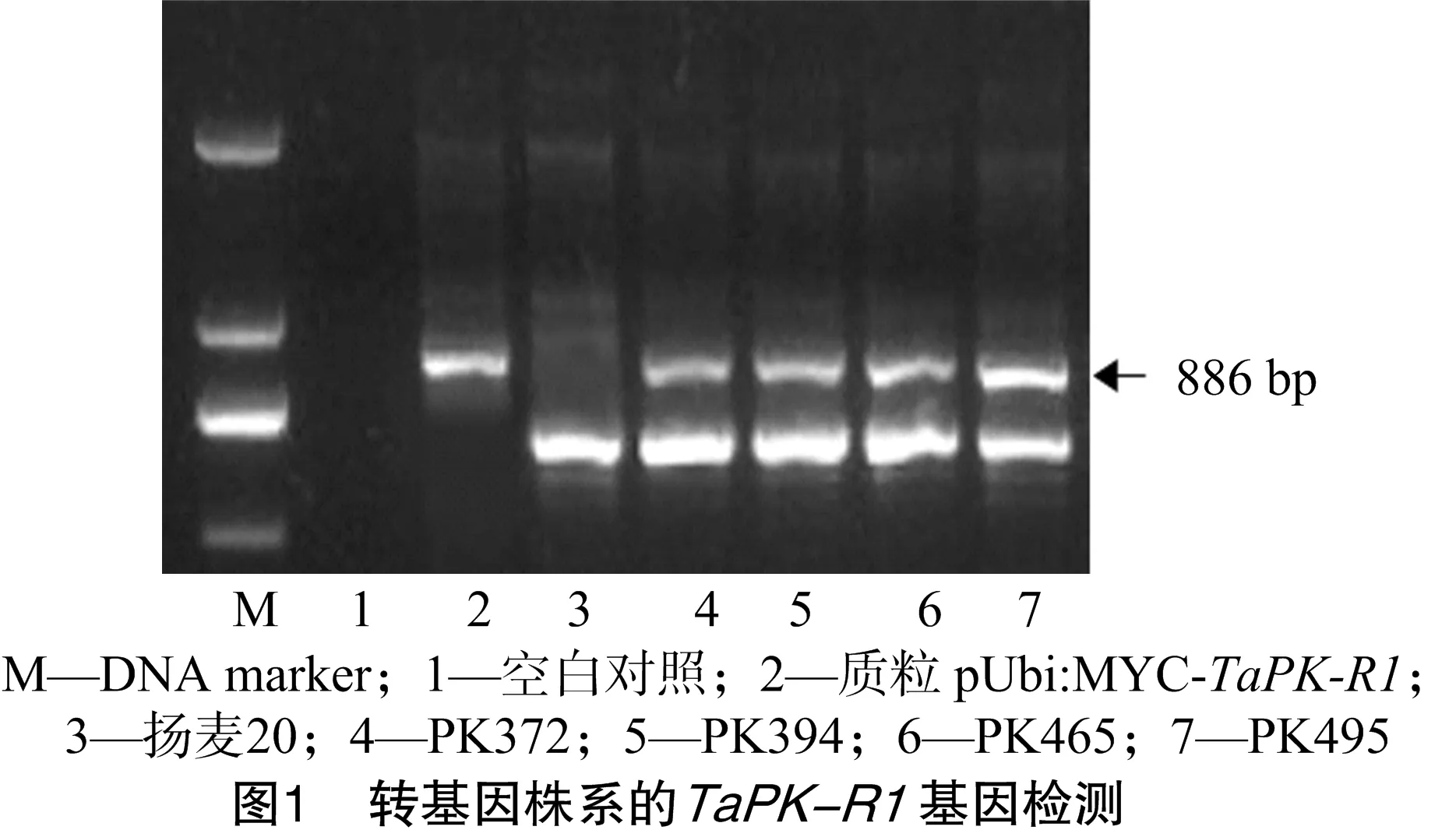
2.2 转基因株系TaPK-R1基因的表达分析结果
在拔节期同时取转基因株系和对照叶片,提取RNA并反转录成cDNA,再采用荧光定量PCR检测TaPK-R1基因的表达,以受体对照扬麦20为基准,计算相对表达量,结果(图2)表明转基因株系PK372、PK394、PK465、PK495的TaPK-R1基因表达量分别是对照的9.0、8.9、9.2、5.9倍,说明导入的目的基因不仅可以正常表达,而且表达量较对照有较大提高。
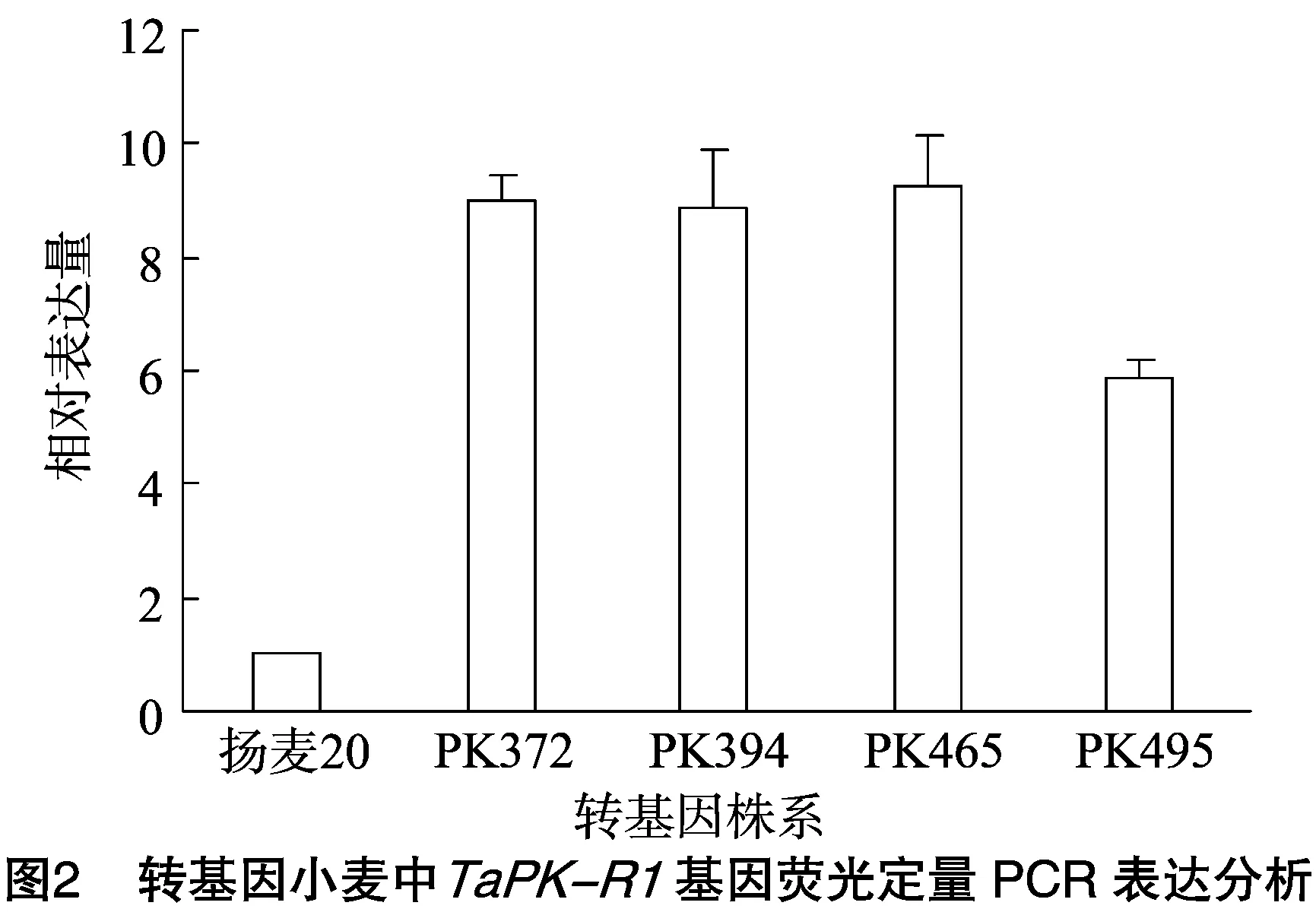
2.3 转基因株系农艺性状评价
2.3.1 生长发育部分阶段的物候期比较 由于播种时间较正常播种期有所推迟,从播种到出苗的时间延长,但转基因株系与对照没有差别,说明TaPK-R1基因的组成型表达没有影响种子的萌发能力。抽穗后气温较高,生育期进程加快,转基因株系与对照相似,全生育期有所缩短,两者几乎没有区别(表1)。
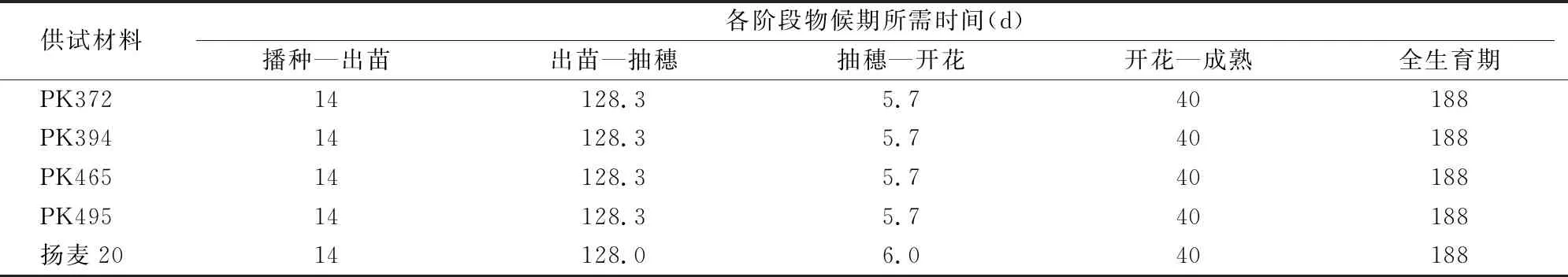
表1 转基因小麦生长发育各阶段物候期的比较
2.3.2 苗情和产量的比较 与受体对照扬麦20相似,转基因株系也属春性小麦,幼苗半直立,株型松散。在基本苗基本一致的情况下,转基因株系的高峰苗数和有效穗数与对照差异不显著,说明TaPK-R1基因的组成型表达没有对小麦的繁茂性和成穗率产生影响。4个转基因株系的产量均较对照有所降低,但差异没有达到显著水平。由于播种推迟,生长期缩短,转基因株系和对照的株高较往年有较大降低,转基因株系与对照也未出现显著差异(表2)。
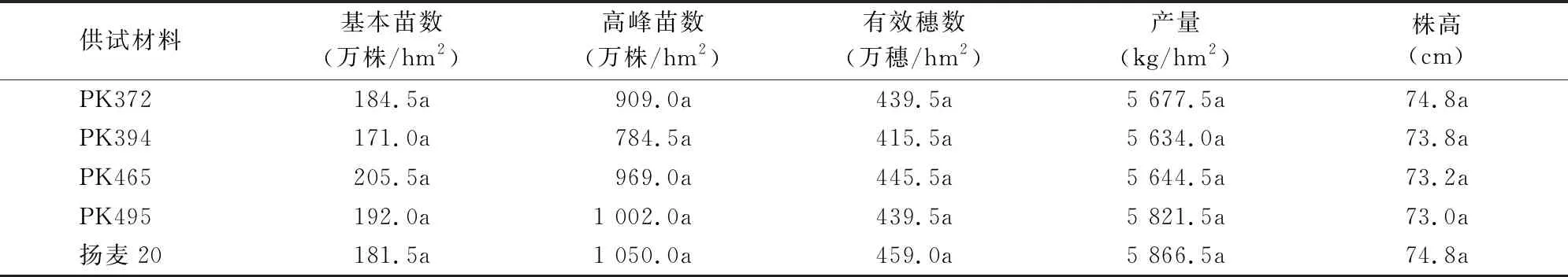
表2 转基因小麦部分重要农艺性状调查结果
注:同列数据后不同小写字母表示在0.05水平差异显著。表3、表4、表5同。
2.3.3 主穗性状的比较 成熟时每个小区随机取10株主穗,考察主穗农艺性状,结果(表3)表明转基因株系的主穗长度、小穗数、不孕小穗数和粒数与受体对照相比均无显著差异。
2.3.4 籽粒性状的比较 籽粒收获晒干后,每小区随机取100粒测定籽粒长度和宽度,结果显示,转基因株系籽粒的长宽虽有所变化,但与对照相比差异不显著。采用近红外测定籽粒蛋白含量发现,转基因株系籽粒蛋白含量较对照有所提高,但未达到显著差异水平。籽粒硬度测定结果与之相仿,转基因株系籽粒硬度比对照稍高,但也没有显著差异,同属于软质麦。千粒质量的分析结果也表明,转基因株系与对照差异不显著(表4)。
对转基因株系和对照的病害调查显示,在整个生育期内小麦黄花叶病没有发生,白粉病和锈病只有轻微发生,两者差异不明显。但后期纹枯病和赤霉病的危害情况有显著差异,扬麦20平均纹枯病病情指数达到50.7%,而转基因株系只有30.7%~36.7%;赤霉病方面,转基因株系的病级要低于对照,普遍率也是同样的趋势(表5),说明TaPK-R1基因的组成型表达不仅提高了小麦对纹枯病的抗性,对赤霉病的抗性也有所提高,可能TaPK-R1是激酶基因,该基因参与了小麦抗病的信号传导,诱导了下游抗病基因的表达,从而提高了小麦对病害的基础抗性。
3 讨论与结论
序列和结构分析表明,TaPK-R1,属于AGC蛋白激酶家族,该家族是植物蛋白激酶的一个亚家族,家族内的激酶均包含C端的丝氨酸/苏氨酸特异性的催化结构域。在哺乳动物中,AGC蛋白激酶是细胞生长、新陈代谢和细胞死亡的重要调节因子,但在植物中AGC蛋白激酶的功能还不是十分清楚。在拟南芥中已经发现39个AGC蛋白激酶成员,根据其激酶催化结构域的变化可将它们分为6个亚族,但只有少数几个AGC蛋白激酶基因的功能得到解析,分别参与植物生长激素的调节、根毛发育以及蓝光信号和活性氧信号的传导。
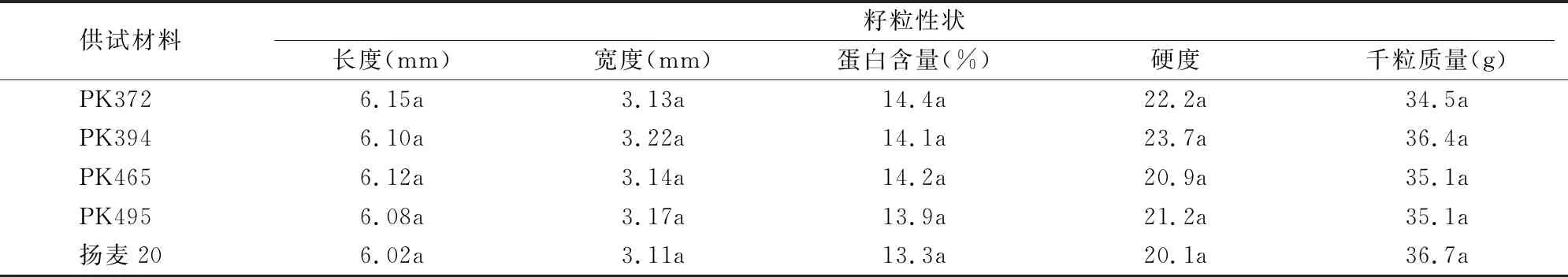
表4 转基因小麦籽粒性状的调查结果
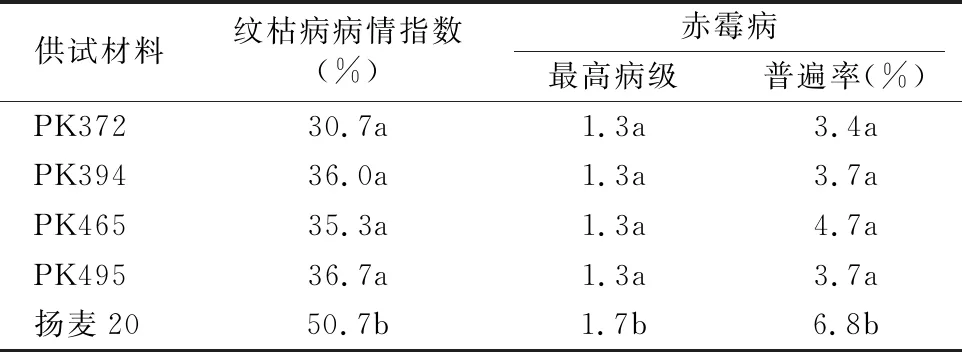
表5 转基因小麦纹枯病和赤霉病抗性表现
目前参与植物病害防御的AGC蛋白激酶基因除了TaPK-R1(TaAGC1)基因外,报道的只有3个基因分别在拟南芥、水稻和番茄中的功能被解析。如拟南芥的OXIDATIVESIGNAL-INDUCIBLE1(AtOXI1)蛋白激酶基因对模式触发免疫(PTI,PAMP triggered immunity)和效应蛋白触发免疫(ETI,effector triggered immunity)等植物免疫反应均有正调控作用[6]。在水稻中,Matsui等发现了与AtOXI1蛋白激酶基因同源的OsOxi1基因,研究结果表明该基因对水稻病害抗性起正调控作用,过表达OsOxi1基因可以增强水稻对稻瘟病(Magnaportheoryzae)的抗性,OsOxi1基因主要通过对OsPti1a的磷酸化,解除OsPti1a对水稻基础抗性的负调控作用,从而提高水稻的基础抗性[7]。番茄的细菌性斑点病主要由番茄细菌性斑点病菌(Pseudomonassyringaepv. Tomato)侵染引起,番茄的Pto激酶基因可以识别病原菌菌株种类,从而产生AvrPto效应物,加强番茄对病原菌侵染的抵抗。Devarenne等研究发现,一种属于AGC Ⅷa的蛋白激酶AvrPto-dependentPto-interactingprotein3(Adi3)可能参与Pto基因的抗病过程,Adi3基因对宿主细胞的程序性死亡起负调控作用,Pto/AvrPto可与Adi3互作,造成病原菌侵染的宿主细胞死亡,从而增加宿主对病原菌的抗性[8]。本研究进一步证实TaPK-R1基因可以提高小麦的抗病性,组成型表达TaPK-R1转基因小麦的纹枯病和赤霉病抗性都得到明显提高。
研究表明,在转基因小麦中组成型表达TaPK-R1基因没有对小麦的生育期、生长习性、产量性状、穗部性状、株高和籽粒性状产生明显的影响。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
科学(2020年2期)2020-08-24
四川农业科技(2019年5期)2019-07-01
西南农业学报(2016年6期)2016-04-16
浙江柑橘(2016年1期)2016-03-11
现代农业(2016年5期)2016-02-28
乡村科技(2016年30期)2016-02-19
吉林大学学报(医学版)(2015年1期)2015-12-17
医学研究杂志(2015年3期)2015-06-10
中国烟草学报(2012年4期)2012-04-09
