
斑鞘豆叶甲触角感器的超微结构
2020-03-09 02:50樊瑞冬伍名渊董亚南史树森
环境昆虫学报 2020年1期
徐 伟,樊瑞冬,伍名渊,董亚南,史树森
(吉林农业大学植物保护学院,长春130118)
昆虫触角的主要功能是嗅觉和触觉,其上着生有多种类型的感器,不同类型感器的外部形态、数量和分布不同,在功能上也存在明显的差异。国内外对鞘翅目许多昆虫触角感器进行了研究,如油菜蓝跳甲Psylliodeschrysocephala(Bartletetal., 1999)、暗黑叩头甲Agriotesobscurus(Meriveeetal., 1997)、锥须步甲Bembidionlampros(Meriveeetal., 2000)和红缘吉丁虫Buprestisfairmairei(刘玉双等,2005)等,发现了10余种触角感器类型,常见有毛形感器、刺形感器和锥形感器等,即使同一类型的感器在不同种类昆虫中形态也有差异,可见感器超微结构的复杂多样(Bruce and Cork, 2001)。这些感器内有特异性神经受体细胞用来探测环境,并将有关信息传递给中枢神经系统(Shields and Hildebrand, 2001),具有嗅觉、触觉和感受温、湿度等功能,使昆虫能够感知环境中的各种信息,如寄主植物、配偶和捕食者发出的化学信息(Knolhoff and Heckel, 2014),并对所处环境进行判断做出正确反应。
斑鞘豆叶甲ColposcelissignataMotschulsky属鞘翅目Coleoptera,叶甲科Chrysomelidae,是东北春大豆苗期的主要害虫之一,主要以成虫为害大豆的叶片、茎、子叶和胚芽,导致大豆叶片出现缺刻、孔洞,由于发生在苗期,在田间呈聚集分布,易造成缺苗断垅,严重影响大豆产量,造成经济损失(沙洪林等,2013)。目前,对斑鞘豆叶甲的寄主选择和聚集行为调控机制等研究较少,通过触角感器的形态和类型探索其生态学功能具有重要意义。本文采用扫描电镜技术对斑鞘豆叶甲触角感器进行观察,明确触角感器形态、类型、数量及分布特点,分析探讨各类触角感器可能的功能和作用,为利用斑鞘豆叶甲的通讯交流机制,进行绿色生态防控提供理论依据。
1 材料与方法
1.1 供试虫源
5月中旬于吉林农业大学大豆试验田采集斑鞘豆叶甲成虫,将成虫分雌、雄收集于扎孔的离心管中,每10头一管,以新鲜大豆叶片在光温培养箱内饲养。环境条件设置为25℃±1℃,RH 50%±5%,L ∶D=16 h ∶8 h。
1.2 触角材料的准备
选取健康斑鞘豆叶甲雌、雄虫各30头,在体视显微镜下用手术刀切取其完整触角,并用小毛笔将触角表面的杂质去除。
1.3 扫描电镜样品的制备与观察
将切下的斑鞘豆叶甲触角放入加入洗洁精的蒸馏水中超声清洗2 s,再用蒸馏水超声清洗3遍。然后用浓度70%、80%、90%和100%的乙醇进行梯度脱水,每个浓度脱水5 min,再将样品放入干燥器内。干燥好的触角用导电胶固定于铜质样品台上,置于JFC-1600型离子溅射仪内进行真空喷金处理,在JSM-6700F型冷场发射电子显微镜下对触角及感器进行扫描观察和拍照,并记录数据。
1.4 数据处理
应用Photoshop CS5软件测量触角柄节、梗节和各鞭亚节长度,以及触角感器的长度和基部直径,应用Excel计算触角不同亚节的长度,及触角感器长度、基部直径和数量的平均值和标准误,采用SPSS 16.0软件以t检验对雌雄触角进行差异显著性分析。采取Schneider命名法对感器进行分类和描述(Schneider, 1964)。
2 结果与分析
2.1 触角的一般形态
斑鞘豆叶甲触角为棕色,丝状,包括柄节(Sc)、梗节(Pe)和鞭节,鞭节有9个亚节构成(F1~F9),雄虫与雌虫触角节数相同(图1;图2-A),但雄虫触角长于雌虫,其中柄节和部分鞭亚节显著长于雌虫。柄节和梗节表皮上均有波纹状隆起,着生感器数量较少(图2-B, C)。第一鞭亚节较短且细,雌虫更加短小纤细,后面亚节逐渐变长变粗。除末亚节呈子弹形,其余各鞭亚节近似长梯形,雄虫鞭亚节基部向端部逐渐变粗,

图1 班鞘豆叶甲雌雄触角长度比较Fig.1 Comparison of antennal length of male and female of Colposecelis signata注:Sc,柄节;Pe,梗节;F1,第1鞭亚节;F2,第2鞭亚节;F3,第3鞭亚节;F4,第4鞭亚节;F5,第5鞭亚节;F6,第6鞭亚节;F7,第7鞭亚节;F8,第8鞭亚节;F9,鞭节末亚节。*表示雌、雄虫间差异显著(P < 0.01)。Note: Sc, scape; Pe, pedicel; F1, the first flagellomere; F2, the second flagellomere; F3, the third flagellomere; F4, the fourth flagellomere; F5, the fifth flagellomere; F6, the sixth flagellomere; F7, the seventh flagellomere; F8, the eighth flagellomere; F9, lateral flagellomere. *Indicate significant difference between male and female (P<0.01).

图2 斑鞘豆叶甲触角的形态及感器分布Fig.2 Antennal segments and sensilla distribution of Colposcelis signata注:A,雄虫触角;B,雌虫触角柄节背面;C,雄虫触角柄节腹侧面;D,雌虫触角中部鞭节;E,雌虫触角末节背面;F,雄虫触角末节背面。Note: A, male antennae; B, dorsal view of scape of female antenna; C, ventral view of scape of male antenna; D, flagellomere of middle of female antenna; E, lateral flagellomere of female; F, dorsal view of lateral flagellomere of female.
前后亚节相互套叠,呈丝状(图2-A);雌虫自第3鞭亚节开始基部明显变细呈环节状,其后部突然加粗,导致分节明显,触角近似念珠状(图2-D, E)。鞭节表皮呈鱼鳞状隆起,腹面感器较背面密集,自第1亚节向后感器数量呈增加趋势,雄虫自第2鞭亚节,雌虫自第3鞭亚节端部感器呈带状区域密集分布,区域内感器类型较多(图2-D),雌虫末亚节2/3处在背面和腹面形成凹陷区,其内感器密集,而雄虫末亚节背面无感器密集区域,感器分布较均匀(图2-E, F)。
2.2 触角感器的类型、特征及分布
通过扫描电镜对斑鞘豆叶甲雌、雄虫触角进行观察,共发现8种类型感器,分别为:毛形感器(sensilla trichodea, ST)、刺形感器(sensilla chaetica, SC)、锥形感器(sensilla basiconica, SB)、指形感器(sensilla finger, SF)、腔锥形感器(sensilla coeloclnica, SCo)、栓锥形感器(sensilla styloconica, SS)、钟形感器(Sensilla Campaniformia, SCa)和Böhm氏鬃毛(Böhm bristles, BB)。
2.2.1毛形感器(Sensilla trichodea, ST)
毛形感器是斑鞘豆叶甲触角上数量最多、分布最广的一类感器,细长如毛发,着生于触角表皮内陷的凹窝内。感器向触角端部伸出,其基部稍粗,向顶端逐渐变尖细,根据形态与长短分为毛形感器Ⅰ型(STⅠ)和毛形感器Ⅱ型(STⅡ)两种亚型。
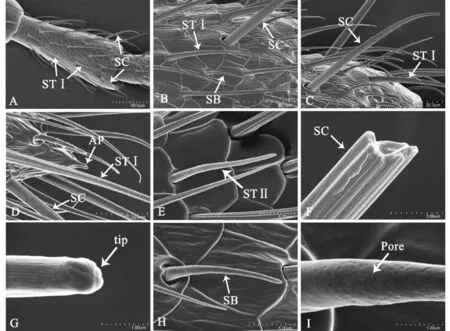
图3 毛形、刺形和锥形感器的形态与分布Fig.3 Morphology and distribution of sensilla of trichodea, chaetica and basiconica注:A,第一鞭亚节的毛形感器Ⅰ型(STⅠ)和刺形感器(SC);B,中间鞭亚节上的毛形感器Ⅰ型(STⅠ),刺形感器(SC)和锥形感器(SB);C,毛形感器Ⅰ型(STⅠ)和刺形感器(SC);D,末节毛形感器Ⅰ型(STⅠ),刺形感器(SC)和末端,AP 3个栓锥形感器;E,毛形感器Ⅱ型,STⅡ表皮纹理;F,刺形感器(SC)表面纹理;G,雄虫刺形感器(SC)顶端;H,锥形感器(SB);I,锥形感器(SB)表皮小孔。Note: A,Sensilla trichodeaⅠ( STⅠ)and Sensilla chaetica(SC) distributed on the first flagellomere; B, Sensilla trichodeaⅠ( STⅠ), Sensilla chaetica(SC) and Sensilla basiconica(SB) distributed on the middle flagellomere; C, Sensilla trichodeaⅠ( STⅠ)and Sensilla chaetica(SC); D, Sensilla trichodeaⅠ(STⅠ),Sensilla chaetica(SC) and three styloconic sensilla on AP, the apical part of the lateral flagellomere; E, Surface texture of sensillum trichodeaⅡ( STⅡ); F, Surface texture of sensilla chaetica(SC); G, Tip of sensilla chaetica(SC) of male; H, Sensilla basiconica(SB); I, Pores of sensilla basiconica(SB).
毛形感器Ⅰ型粗且长,与触角表面呈20°~30°夹角,在二分之一处稍弯曲,呈弧线形(图3-A, B)。感器表面有明显的纵脊,纵脊上有平行的纵纹(图3-C, D)。感器长39.64~60.94 μm,基部直径1.70~4.72 μm,在各节均有分布(图3-A, B)。
毛形感器Ⅱ型稍短,向端部逐渐变细,表皮上无纵脊和纵纹,隐约可见小孔。感器在端部稍弯曲。感器长18.16~32.53 μm,基部直径1.12~2. 50 μm。分布在鞭节,多着位于触角的腹侧面(图3-E)。
2.2.2刺形感器( Sensilla chaetica, SC)
刺形感器刚直坚硬如刺,有规律地着生于开阔的臼窝内,可活动,臼窝边缘突出明显(图3-A, B, C)。感器与触角表面呈60°左右的夹角,明显长且高于其它感器,表面具明显粗大的纵脊,每条纵脊两侧面具斜向对称的纵纹(图3-F)。刺形感器基部较粗向端部渐细,雄虫的近端部稍弯曲,端部稍膨大,顶端呈花瓣状聚拢(图3-G)。雌虫的端部弯曲不明显,端部稍纤细。刺形感器长49.14~67.16 μm,基部直径3.23~4.88 μm。每节3~12个,柄节、梗节、第1、2鞭亚节位于近端部,第3~8亚节位于前中部和近端部,末节则在基部也有分布。雌虫分布于鞭亚节前中部和基部的刺形感器与触角表面贴近,夹角明显小于雄虫。
2.2.3锥形感器(Sensilla basiconica, SB)
锥形感器呈短锥形,着生于触角表皮内陷的开阔凹窝内,分布于2~11鞭亚节,散生于毛形感器之间(图3-B, H)。感器基部稍粗,凹窝边缘具隆起明显,较毛形感器短,向端部明显变细,顶端稍钝圆,表皮具明显稍浅的小孔(图3-I)。感器长10.41~14.01 μm,基部直径1.17~1.78 μm。每鞭亚节有16~30个。
2.2.4指形感器(Sensilla finger, SF)
指形、腔锥形、栓锥形和钟形感器均分布于鞭亚节端后部感器密集条带区(图4-A, B),着生于触角表皮隆起圆台上的浅窝内,另外,在末亚节凹陷区内指形感器密集分布。该感器柱状细长,形如手指,雄虫始于第2鞭亚节,雌虫始于第3鞭亚节出现。指形感器在开始出现的亚节仅有2个,向后逐渐增多,并成群出现。感器粗细均匀,基部有延伸到圆台上的明显树状突起,端部稍尖,表皮具明显较深的漏斗形小孔(图4-B, C)。感器长12.46~22.67 μm,基部直径1.31~2.21 μm。
2.2.5腔锥形感器(Sensilla coeloconica, SCo)
腔锥形感器呈锥体状,雄虫自第2鞭亚节,雌虫自第3鞭亚节出现(图4-A, D)。感器基部稍粗,同样有明显树状突起;端部呈指状聚拢,高倍镜下隐约可见有浅的小孔,指间具缝隙(图4-E)。感器基部直径1.62~2.03 μm,锥状体高4.96~6.67 μm。每节2~5个。
2.2.6栓锥形感器(Sensilla styloconica, SS)
栓锥形感器数量较少,呈柱状,上面有一较粗的类似栓钉状的锥状体,粗细均匀。雄虫自第2鞭亚节出现,雌虫自第3鞭亚节出现,每亚节1~3个。根据栓锥形感器柱状体和其上方锥状体表皮形态,分为栓锥形感器Ⅰ型(SSⅠ)和栓锥形感器Ⅱ型(SSⅡ)两个亚型。
栓锥形感器Ⅰ型锥状体表面具网状突起,顶端中部稍凹陷,下面的柱状体表面有明显较粗的树状突起一直延伸到圆台边缘(图4-F);栓锥形感器Ⅱ型锥状体表面光滑,顶端圆顿,下面柱状体基部有树突状脊纹终止于圆台凹窝边缘(图4-G, H)。栓锥形感器Ⅰ型柱状体基部直径0.98~1.79 μm,锥状体高2.72~4.10 μm,锥状体基部直径0.45~0.67 μm。栓锥形感器Ⅱ型柱状体基部直径2.08~2.66 μm,锥状体高2.75~3.55 μm,锥状体直径0.58~0.78 μm。
2.2.7钟形感器(Sensilla Campaniformia, SCa)
钟形感器呈心形隆起,边缘是一圈褶状凹陷,形似纽扣。仅在雄虫触角第4鞭亚节发现该感器,心形隆起直径1.50~1.61 μm(图4-I)。
2.2.8Böhm氏鬃毛(Böhm bristles, BB)
Böhm氏鬃毛短小,呈刺状,表面光滑无孔,着生于触角表皮的凹陷内,成簇分布于触角柄节前缘背面、腹面凹陷两侧(图5-A),梗节端部与柄节连接处(图5-B)。感器长6.06~7.52 μm,基部直径1.73~2.28 μm。
2.2.9表皮孔(Cuticular pore, CP)
在触角表面具有许多不规则小孔,各节均有分布,鞭节较多,梗节最少,多在毛形和刺形感器周围单独出现,个别刺形感器周围有多个小孔成群出现(图5-C, D),也有的在触角表皮上远离感器(图5-B)。
2.3 雌、雄虫触角感器的数量和大小
斑鞘豆叶甲雄虫触角感器数量多于雌虫,着生更加密集,其中雄虫毛形感器Ⅰ型、Ⅱ型和刺形感器平均数量分别为670.64±30.27、276.53±14.75和94.65±3.70明显多于雌虫(630.58±25.32、234.83±10.42和85.22±2.11),但指形感器数量雌虫为86.21±2.27明显多于雄虫(70.94±1.88)(表1)。雄虫各类型感器均较雌虫粗壮,其中毛形感器Ⅰ型、Ⅱ型、刺形感器和指形感器均明显长于雌虫,各感器基部直径均长于雌虫。

图4 腔锥形、指形、栓锥形、钟感器的形态与分布Fig.4 Morphology and distribution of sensilla of coeloconica, finger, styloconica and campaniformia注:A,鞭亚节端部感器;B,指形感器(SF);C,指形感器(SF)局部放大;D,腔锥形感器(SCO);E,腔锥形感器(SCO)局部放大;F,栓锥形感器Ⅰ型(SSⅠ);G,栓锥形感器Ⅱ型(SSⅡ);H,栓锥形感器Ⅱ型(SSⅡ);I,钟形感器(SCa)。Note: A, Sensilla on the distal margin of flagellomere; B, Sensilla finger(SF); C, Partial enlarged sensilla finger(SF); D, Sensillum coeloconica(SCO); E, Partial enlarged sensillum coeloconica(SCO); F, Sensilla styloconicaⅠ(SSⅠ); G,Sensilla styloconicaⅡ( SSⅡ); H,Sensilla styloconicaⅡ( SSⅡ); I,Sensilla campaniformia(SCa).

图5 Böhm氏鬃毛和感觉孔的形态与分布Fig.5 Morphology and distribution of Böhm bristles and cuticular pores注:A,柄节腹面Böhm氏鬃毛(BB);B,柄节表皮孔(CP)和梗节前端Böhm氏鬃毛(BB);C,毛形感器(STⅠ)附近的表皮孔(CP);D,刺形感器(SC)周围的表皮孔(CP)。Note: A, Böhm bristles(BB) on the ventral and proximal side of the scape; B, Cuticular pores(CP) on the scape and Böhm bristles(BB) on the proximal pedicel; C, Cuticular pore(CP) near sensilla trichodeaⅠ(STⅠ); D, Cuticular pore(CP) around sensilla chaetica(SC).

表1 斑鞘豆叶甲触角感器的大小、数量与分布
注:数据为平均值±标准差。不同字母表示雌、雄虫间差异显著(P< 0.05)。
Note: The data in the table represents the mean±SD. Different letters indicate significant difference between male and female (P< 0.05).
3 结论与讨论
目前国内外学者对于鞘翅目许多昆虫的触角进行了研究,它们的感器类型复杂多样,不同类群昆虫的触角感器形态变化较大,相同类群的感器类型、形态和分布相似。例如,叶甲科的双斑长跗萤叶甲Monoleptahieroglyphica(张聪等,2012)、油菜蓝跳甲Psylliodeschrysocephala(Bartletetal., 1999)和黄曲条跳甲Phyllotretastriolata等(Zhangetal., 2016;刘晓梅等,2018)与本研究中的斑鞘豆叶甲触角均在鞭亚节的端部和末节凹陷区出现较多的感器类型,且分布密集,这种着生方式有利于增强触角对各种刺激的反应。而毛形感器在这些甲虫的触角各节分布较均匀,是数量最多的感器,具有2~3个亚型,其中长毛亚型感器(毛形感器Ⅰ型)表面均具有明显的纵脊和纵纹。Bartlet(1999)和Merivee等(2000)发现在毛形感器基部具柔性臼窝,下面连有一个关节膜和悬垂纤维,只有一个感觉神经元,树状突外节终止于管状体上,因此认为该感器具有机械感觉功能。对于短毛亚型(毛形感器Ⅱ型)未见双斑萤叶甲和黄曲条跳甲关于表皮孔的描述,斑鞘豆叶甲仅在高倍电镜下隐约可见小孔,Bartlet等(1999)通过透射电镜确定油菜蓝跳甲两种短毛型均为厚壁具孔,有两个感觉神经元,Mustaparta(1975)发现Hylobiusabietis毛形感器具孔对信息素马鞭草醇和寄主植物挥发物α-pinene和β-pinene等有电生理响应,具有嗅觉功能。而蛾类毛形感器除对植物气味有反应,还是性信息素的受体(Binyameenetal., 2012)。斑鞘豆叶甲两种亚型的毛形感器雄虫均明显多于雌虫,这种现象在Tetriguslewisi等种类中也有发现,表明毛形感器雌雄在生理功能上存在差异(Renetal., 2014.),但是否对性信息素有反应在鞘翅目中还未被证实。
除了毛形感器,具有嗅觉功能的还有锥形、指形和腔锥形感器。锥形感器普遍存在于甲虫触角上,内含多个神经细胞,表皮壁分布有选择通透性的微管,是一种能识别植物气味分子的嗅觉感器(Lopesetal., 2002)。斑鞘豆叶甲仅发现一种锥形感器,表皮具清晰小孔,但雌雄数量无明显差异。而在Limoniusaeruginosus(Meriveeetal.,1998)、四纹豆象Callosobruchusmaculatus和绿豆象Callosobruchuschinensis(Huetal., 2009)等鞘翅目昆虫中发现雄性锥形感器的数量明显高于雌性,推测雄性锥形感器除了能够感受寄主气味还能感受雌性个体释放的性信息素。指形感器是斑鞘豆叶甲触角上表皮明显具孔的一种感器,其孔径大于锥形感器,有人也将其作为锥形感器的一个亚型(刘晓梅等,2018)。研究发现不同种类甲虫的指形感器在雌雄虫间的着生数量不同,例如Phyllotretacruciferae等4种叶甲中雌雄数量相当(Ritcey and McIver, 1990),黄曲条跳甲雄虫的指形感器数量明显多于雌虫(刘晓梅等,2018),而斑鞘豆叶甲则雌虫数量较多,这表明指形感器可能在斑鞘豆叶甲寄主植物检测和选择产卵场所中起着重要的作用。鞘翅目昆虫腔锥形感器不同于鳞翅目,一般为无缘毛腔锥形,在不同昆虫中功能不同,有的含有嗅觉受体细胞,能够感受植物挥发物;有的具有感受温湿度变化、水蒸气和二氧化碳的作用(Hallberg, 1982;Zacharuketal., 1985)。斑鞘豆叶甲腔锥形感器中央锥状体呈指状聚拢,指间有缝隙并且发现指状结构上有浅的孔存在,应具有嗅觉感受功能。
昆虫触角的另外一个重要功能是触觉作用,刺形感器和Böhm氏鬃毛则是重要的感受机械刺激的感器。刺形感器一般突出于触角表面,基部有臼窝,雄虫数量多于雌虫,有些种类的刺形感器端部具孔(Isidoroetal., 1998; Sunetal., 2010),Isidoro等(1998)发现油菜蓝跳甲刺形感器内有5~6个感觉神经元,其中一个神经元的树突终止于管状体,而其他神经元的树突延伸到端孔中,这种结构表明同时具有味觉和机械感觉的功能,电生理记录也证实其中的化学感觉细胞对硫代葡萄糖苷和油菜提取物反应敏感。斑鞘豆叶甲刺形感器与其他叶甲类昆虫在着生位置上相同,但雌雄在着生方式和形态上存在差异,雄虫刺形感器粗壮挺立,端部明显膨大呈花瓣状聚拢,而雌虫刺形感器则稍细,位于基部的与触角表面夹角小,感器端部无明显膨大结构,这种差异表明雌雄生理功能不同。而斑鞘豆叶甲雄虫末端膨大的花瓣状结构与鳞翅目的黄刺蛾Monemaflavescens(Yangetal., 2016)触角上的刺形感器末端结构相似,在鞘翅目中未见刺形感器端部有此特化结构的报道,这种分化是否更有利于发挥味觉功能需要进一步研究证实。斑鞘豆叶甲Böhm氏鬃毛与其他甲虫类似成簇分布于柄节和梗节关节连接处,其基部位于表皮凹陷的穴内,易感受机械刺激(刘玉双和石福明,2005;张聪等,2012;刘晓梅等,2018)。Schneider等(1964)研究认为Böhm氏鬃毛能够缓冲机械刺激发生的重力作用,减缓触角位置的下降速度。
在斑鞘豆叶甲触角上还发现了栓锥形感器和钟形感器。栓锥形感器在鞘翅目中较少发现,感器内部有丰富的神经细胞,具有感受温湿度的功能(Bartletetal.,1999)。斑鞘豆叶甲栓锥形感器有两种亚型,一种锥状体光滑,另外一种表皮具网状突起,顶端凹陷,这两种亚型是否在功能上存在差异有待进一步研究。斑鞘豆叶甲钟形感器仅分布于雄虫第4鞭亚节上,数量最少,在其他叶甲科昆虫中也较少发现。在栗山天牛Massicusraddei(魏建荣等,2013)和Platynusassimilis(Mustetal., 2010)等中少量发现,Must等(2010)的研究表明Platynusassimilis可依靠钟形感器选择栖息地,感知环境温度,避免过冷或过热的情况出现,Bembidionlampros钟形感器则被认为是能够感知应力变化的机械感器(Merieeetal., 2000)。另外斑鞘豆叶甲虫触角上发现有表皮孔,雌、雄间数量无显著差异,多分布于刺形感器和毛形感器周围。在Psylloidesaffinis、油菜蓝跳甲和栗山天牛的触角上也发现了表皮孔(Ritcey and McIver, 1990; Bartletetal., 1999; 魏建荣等, 2013),目前对表皮孔的功能研究较少,认为是表皮腺体的开口,在一些昆虫中分泌的物质具有酶活性,降解信息素的分子或寄主植物挥发物以防止它们对触角化学感器过载(Prestwich and Blomquist, 1987; Dickensetal., 1992)。
斑鞘豆叶甲的触角感器丰富,存在明显的性二型现象,刺形、栓锥形和钟形感器与其他同类昆虫也存在差异,表现出种间分化的特性,应通过透射电镜、电生理等试验对斑鞘豆叶甲触角感器的形态和功能进一步探索,明确斑鞘豆叶甲的触角感受和行为机制。
猜你喜欢
广西林业科学(2022年1期)2022-03-03
作物研究(2021年6期)2021-12-25
甘肃农业科技(2021年6期)2021-07-25
环境昆虫学报(2021年1期)2021-03-30
江西农业学报(2019年10期)2019-10-29
延安大学学报(自然科学版)(2019年3期)2019-10-11
科学导报(2019年35期)2019-09-03
河南农业科学(2019年5期)2019-05-28
植物保护(2018年3期)2018-05-14
江苏农业科学(2014年3期)2014-07-16
