
小兴安岭阔叶混交林距道路边缘不同距离地表节肢动物多样性*
2020-03-08 08:09:10高梅香李晓坤黄金龙
哈尔滨师范大学自然科学学报 2020年5期
李 佳,张 蕊,高梅香,李晓坤,黄金龙
(1.哈尔滨师范大学; 2.宁波大学; 3.黑龙江省丰林国家级自然保护区管理局)
0 引言
自然保护区中的道路对森林管理工作具有重要作用,虽然与公路相比部分林区道路更窄且没有铺设路面,但已有研究表明林区道路同样会对土壤侵蚀和沉积、植被和动物造成一定程度的影响[1-5].地表节肢动物作为土壤生态系统的重要组成类群,在生态系统物质循环和能量转换中发挥着积极作用,与生态系统多方面信息密切相关[6].国外学者侧重于探究道路改变原有生境对于地表节肢动物群落的影响,如揭示道路邻近度对陆生节肢动物的生境特异性影响[7];阐明道路作为森林系统开放空间对植被和地表节肢动物群落的影响[8].另外,国内已有学者关注到道路对土壤动物多样性的影响[9-10],探讨了公路路域植被演替的不同阶段土壤动物群落结构特征[11-13]以及土壤动物对路域植被恢复的指示作用[14]等,但大多选择在高速公路两侧.而自然保护区除了能保护自然生态系统[15]、维持生物多样性外[16], 还能提供与人类自身需求直接相关的多种生态系统服务[17],因此探讨保护区内道路边缘地表节肢动物群落特征对于生物多样性保护和森林生态系统管理具有重要的现实意义.
丰林自然保护区内阔叶混交林受人为干扰较少, 林分群落结构稳定且保持了自然演替的所有特征,具有很高的代表性[18].该实验选择在丰林自然保护区未铺设路面的林区道路旁进行,在距道路边缘不同距离设置三条样线(分别距道路边缘0m、5m、20m),于2018年6月12~23日采用陷阱法对地表节肢动物群落进行调查. 该实验欲阐明以下科学问题:(1)距道路边缘不同距离地表节肢动物群落组成特征;(2)距道路边缘不同距离地表节肢动物多样性特征;(3)距道路边缘不同距离地表节肢动物beta多样性特征,最终阐明小兴安岭阔叶混交林距道路边缘不同距离地表节肢动物多样性,为生物多样性保护和森林生态系统管理提供基础数据和理论支撑.
1 研究地区与研究方法
1.1 研究地区
研究区为丰林国家级自然保护区(48°02'~48°12' N,128°58'~129°15' E),该区是目前世界上保存较为完整的北温带原始阔叶红松林分布区.位于小兴安岭南坡北段,海拔280~683 m,属坡状的低山丘陵地貌,土壤主要为山地棕色森林土.气候属北温带大陆性季风气候,夏季高温多雨而冬季寒冷干燥,年均温-0.5 ℃,年降水量650 mm左右.该实验在保护区36林班内进行,该林班植被类型是1931年自然火灾形成的次生白桦林逐步演替形成的阔叶混交林,主要乔木为白桦(Betulaplatyphylla)、红皮云杉(Piceakoraiensis)、臭冷杉(Abiesnephrolepis)、色木槭(A.mono)、春榆(Ulmuspropinqua)、香杨(Populuskoreana)等;灌木为毛榛子(Corylusmandshurica)、忍冬(Lonicerajaponica)等;草本为蕨类(Pteridophyta)、苔草(Carextristachya)等[19].
1.2 样地设置和样品采集
实验样地设置在丰林自然保护区36林班内,该林班的乔木林地为国家一级公益林,实验样地邻近未铺设路面的林区道路(宽约5m).该实验在林区道路南侧阔叶混交林中设置三条和道路平行的样线,这三条样线分别距道路边缘0、5和20 m(分别标记为Dis_0、Dis_5、Dis_20).在每条样线上以40m为间隔分别设置5个10 m×5 m的样方,其长轴为沿道路方向,分别记为Plot_1、Plot_2、Plot_3 、Plot_4、Plot_5.该实验沿着3条样线分别在15个样方内,使用陷阱法捕捉地表节肢动物.在每个样方内,将三个陷阱随机布置在样线上,首先使用土钻(内径7cm)挖取一个约深15cm的柱状土坑,使用塑料杯(高14cm、内径7cm)作为陷阱,将杯子放入打好的土坑中,使杯口与地面齐平,杯中倒入饱和NaCl溶液(大概至杯子三分之二处),为了防止雨水、凋落物和林冠节肢动物等进入陷阱,在杯口上方约10 cm处覆盖由四根筷子支撑的一次性塑料透明餐盒.由于6月份为雨季,为了防止陷阱内雨水过多,该实验于2018年6月12日至23日连续12 d内进行五次样品采集,后将五个样品合到一起进行整体分析.
在室内将挑拣出来的所有地表节肢动物放于95%医用酒精中保存.主要依据《中国土壤动物检索图鉴》[20]、《原色中国东北土壤甲虫图鉴——步行虫类》[21]、《原色中国东北土壤甲虫图鉴——隐翅虫类拟步甲类》[22]、《中国东北的葬甲科研究》[23]等鉴定标本.步甲科和葬甲科鉴定到种,其它地表节肢动物鉴定到科或目.样品中的成虫与幼虫分别计数,仅将成虫用于后续处理分析.
1.3 数据处理分析
1.3.1 多度等级划分和多样性指数
多度等级划分:按照个体数占总捕获量的百分比来划分多度等级,个体数占总捕获量的 10%以上、1%~10%、1%以下分别为优势类群(物种)、常见类群(物种)和稀有类群(物种).采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)和Simpson优势度指数(C)对全部地表节肢动物、步甲科和葬甲科群落多样性进行分析,其中以地表节肢动物类群数(物种数)来代表丰富度.计算公式如下:
(1)
J=H′/lnS
(2)
(3)
式中,Pi=ni/N,ni为第i类群(物种)个体数,N为群落个体总数,S为群落类群数(物种数).
1.3.2 方差分析
为了保证数据的正态分布,首先对地表节肢动物群落个体数和类群数(物种数)及多样性指数进行正态分布检验,对不符合正态分布的数据进行对数转换.采用单因素方差分析说明距道路边缘不同距离内全部地表节肢动物、步甲科和葬甲科个体数和类群数(物种数)的差异显著性.
1.3.3 Beta多样性
Beta多样性是指同一区域内沿环境梯度不同生物群落之间物种组成的差异性[24-26].通过比较不同样线上地表节肢动物beta多样性的不同,来描述距道路边缘不同距离地表节肢动物群落结构特征.该文采用 PERMDISP2[27]方法分别计算不同样线上全部地表节肢动物、步甲科和葬甲科群落的beta多样性,并使用置换检验分别评估不同样线间全部地表节肢动物、步甲科和葬甲科群落beta多样性的差异显著性.基于 Sørensen 非相似性计算beta多样性(βSOR),再将其分解为基于多地点的空间物种转换(βSIM)和物种集群镶嵌(βSNE)[28].计算公式如下:
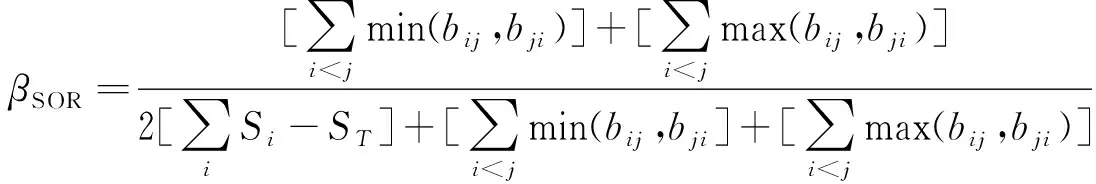
(4)
(5)
βSNE=βSOR-βSIM=
(6)
式中,Si为样点i的全部类群数(步甲、葬甲物种数),ST为全部调查样点的全部类群数(步甲、葬甲物种数),bij和bji分别为群落i和群落j各自所特有的类群数(步甲、葬甲物种数).βSOR为群落Sørensen非相似性,βSIM为群落Sørensen非相似性的空间物种转换部分(即Simpson非相似性),βSNE为群落Sørensen非相似性物种集群镶嵌部分.由于空间物种转换和物种集群镶嵌是两个分离的可加部分,所以全部beta多样性表达式为βSOR=βSIM+βSNE[29].
在Microsoft Excel 2010中对原始数据进行基本的整理分析和预处理.通过R(3.4.3)软件的vegan程序包中的shapiro.test函数进行正态分布检验、aov函数进行方差分析、diversity函数计算多样性指数、betadisper函数计算beta多样性、permutest函数进行检验(置换999次).
2 结果与分析
2.1 道路边缘地表节肢动物群落组成特征
该研究共采集地表节肢动物2662只,隶属于13目15科,其中步甲科、蚁科和蜘蛛目为优势类群,占地表节肢动物总个体数的71.90%.葬甲科、隐翅虫科、金龟子科、马陆目、蜈蚣目、盲蛛目、同翅目为常见类群,占地表节肢动物总个体数的24.83%.叶甲科、天牛科、叩甲科、球蕈甲科、拟天牛科等13个稀有类群,占地表节肢动物总个体数的3.08% .
从Dis_0到Dis_20,地表节肢动物个体数逐渐增多,而类群数先增多后减少,但单因素方差分析表明全部地表节肢动物个体数和类群数在三条样线间差异均不显著.步甲科、蚁科和蜘蛛目在距道路边缘不同距离三条样线上均为优势类群,而马陆目仅在Dis_0为优势类群,在其他两条样线上为常见类群;金龟子科仅在Dis_0为常见类群,在其它两条样线上为稀有类群;球蕈甲科、直翅目仅在Dis_20为常见类群,在其它两条样线上为稀有类群(见表1).但单因素方差分析结果显示每一类群个体数在三条样线间差异均不显著.

表1 距道路边缘不同距离地表节肢动物群落组成
该实验共采集步甲科335只22种,其中Pterostichusadstrictus和Pterostichusmaoershansis为优势种,占步甲科总个体数的69.24%.Bembidionlissonotum、Platynusezoanus、Morphocarabushummelismaragdulus、Morphocarabushummliwangxingensis、Megodontusvietinghoffibowringi、Carabusbillergisedakovi、Aulonocarabuscanaliculatus为常见种,占步甲科总个体数的25.04%.Pterostichussulcitarsis、Morphocarabusvenustuschenpengi、Elaphruspunctatus等13个稀有物种占步甲科总个体数的5.57%.从Dis_0到Dis_20,步甲科个体数先减少后增多,而物种数先增多后减少,但单因素方差分析表明步甲科个体数和物种数在三条样线间差异均不显著.Carabusbillergisedakovi仅在Dis_0为优势种,在其他两条样线上为常见种;Aulonocarabuscanaliculatus仅在Dis_20为优势种,在其他两条样线为常见种.Chlaeniuscostiger、Lebiabifenestrata、Carabusgranulatustelluris、Acoptolabrusschrenchizhangensis、Morphocarabusvenustuschenpengi仅在Dis_0出现;Elaphruspunctatus、Morphocarabuswulfiusijilinensis、Colpodeselainus、Acoptolabrusconstricticollisheilongjiangensis、Lechnolebiacribricollis仅在Dis_5出现;Brachininaesp仅在Dis_20出现(见表2).
共采集葬甲科71只3种,其中Silphaperforataperforata为优势种,占葬甲科总个体数的88.73%,该物种在Dis_0和Dis_20均为优势物种,但在Dis_5为稀有物种.Nicrophorusvespilloides和Dendroxenasexcarinata为常见物种,占葬甲科总个体数的11.27%(表2).从Dis_0到Dis_20,葬甲科个体数先减少后大幅度增加,物种数则逐渐增加,单因素方差分析表明三条样线间葬甲科个体数存在极显著差异,但物种数差异不显著.
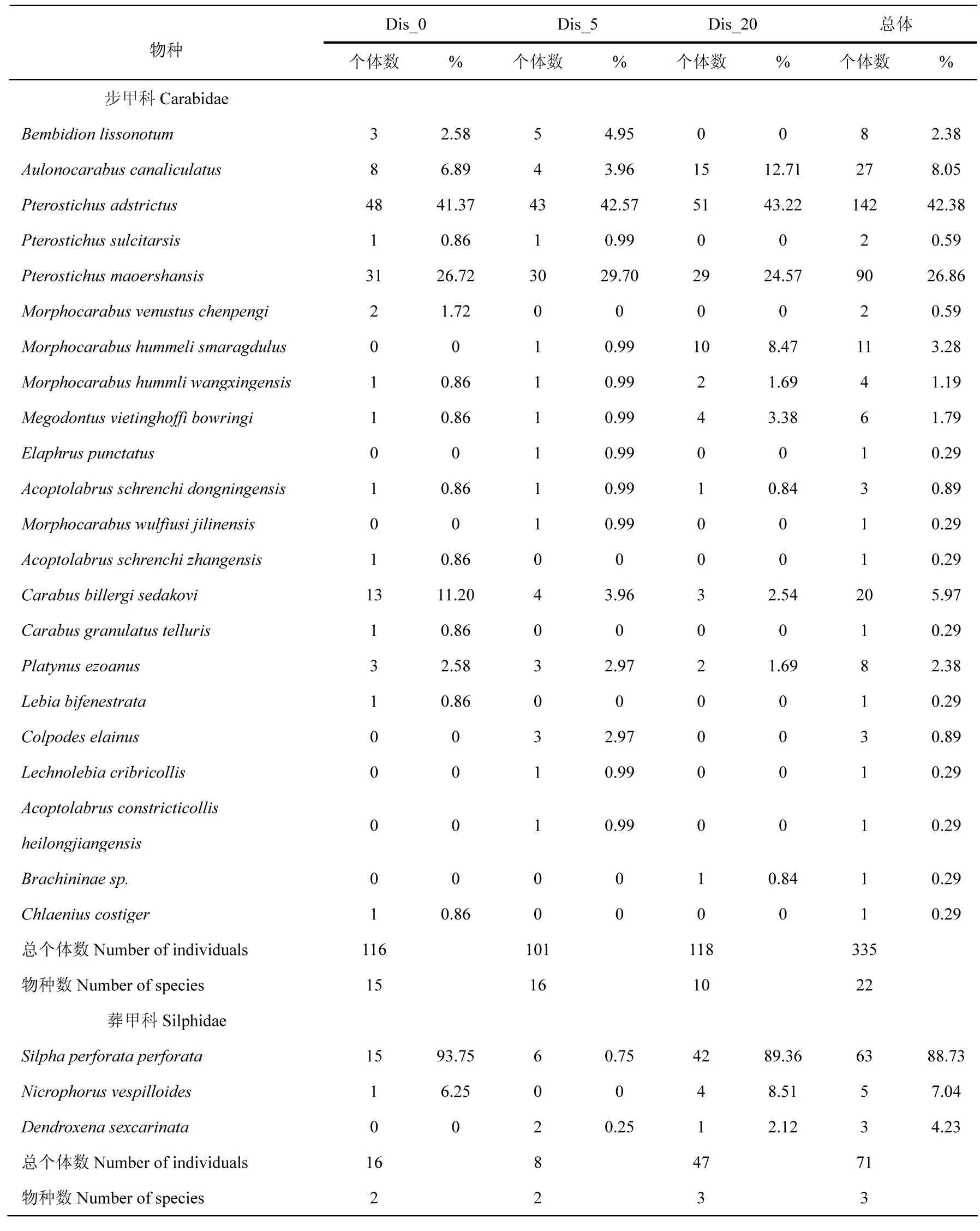
表2 距道路边缘不同距离步甲科、葬甲科物种组成及其个体数
2.2 距道路边缘不同距离地表节肢动物群落多样性特征
不同样线上地表节肢动物多样性指数H′和均匀度指数J值由大到小依次为:Dis_0> Dis_20>Dis_5;优势度指数C的大小次序则与二者相反.但在Dis_20的多样性指数H′仍低于Dis_0的指数.结果表明在距道路边缘较近的样线上(Dis_0附近)地表节肢动物多样性最高,向森林内部距道路边缘较远的样线上(Dis_20)地表节肢动物多样性反而有降低的趋势(如图1所示).

图1 距道路边缘不同距离地表节肢动物多样性注:H′: 多样性指数 J: 均匀度指数 C: 优势度指数
单因素方差分析结果显示(见表3),全部地表节肢动物多样性指数(H′)、均匀度指数(J)、优势度指数(C)在距离道路边缘不同距离样线间差异均不显著.
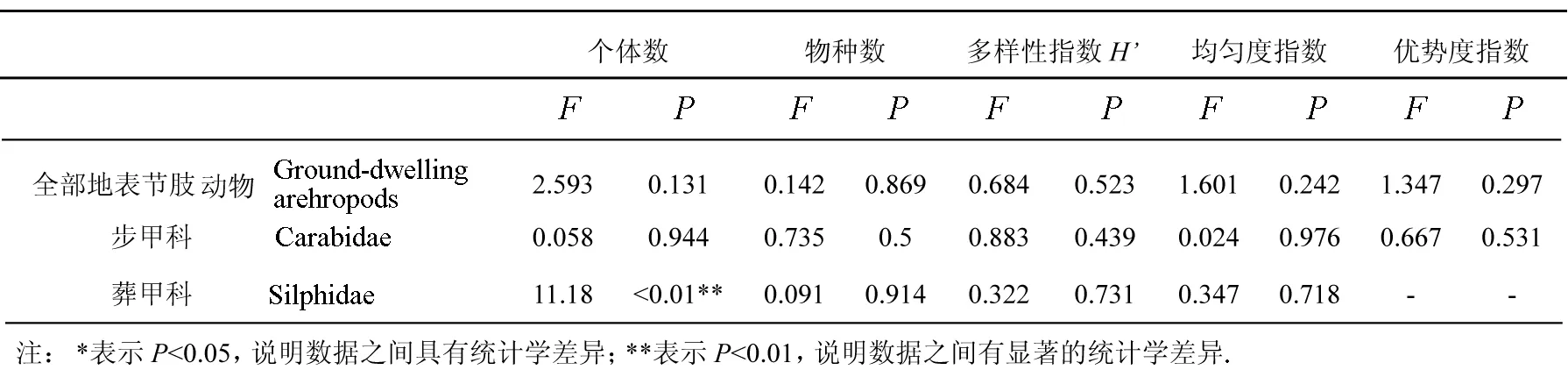
表3 距道路边缘不同距离样线上对地表节肢动物群落多样性的差异性
2.3 道路边缘不同距离地表节肢动物的Beta多样性特征
根据beta多样性结果可以看出(如图2所示),全部地表节肢动物在Dis_0样线上群落物种组成差异性最小,在Dis_20样线上群落物种组成差异性最大;步甲科则呈现相反特征,在Dis_0样线上群落物种组成差异性最大,在Dis_20样线上群落物种组成差异性最小;而葬甲科由于捕获数量较少,其群落物种组成差异性在三条样线上具有一致性.置换检验结果表明在距道路边缘不同距离样线间全部地表节肢动物、步甲科和葬甲科的 beta 多样性均具有显著差异(P<0.05).
全部beta多样性被分解之后,在距道路边缘不同距离样线上除了Dis_20的步甲科和全部葬甲科外,全部地表节肢动物及Dis_0和Dis_5的步甲科的空间物种转换(βSIM)均大于0.4,而物种集群镶嵌(βSNE)均小于0.2(如图2所示).
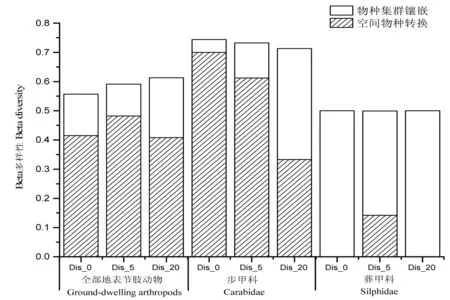
图2 距道路边缘不同距离地表节肢动物的空间物种转换和物种集群镶嵌
3 讨论
3.1 群落组成
地表节肢动物在距道路边缘不同距离样线上的共有优势类群为步甲科、蚁科、蜘蛛目,共有常见类群为隐翅虫科、马陆目、蜈蚣目、盲蛛目.但其中马陆目在Dis_0为优势类群,在其它两条样线上为常见类群,这可能由于马陆同化率与温度呈正相关且具有一定的依赖性[30],而距道路边缘最近的区域植被覆盖率较低,光照时间长温度较高[31],为马陆提供了良好的生长、繁殖条件,从而导致其群落丰富度略高于相对密闭的森林环境.
全部地表节肢动物个体数从Dis_0到Dis_20逐渐增加,但全部地表节肢动物及每个类群(物种)个体数在距道路边缘不同距离样线间均不存在显著差异.从共有优势类群个体数在距道路边缘不同距离样线间的变化特征来看,步甲科个体数从Dis_0到Dis_20逐渐减少,由于道路对地表甲虫具有向导作用,可能引导其沿路边移动[32],所以路边捕获量可能高估了其真正活动密度,从而表现出与毗邻森林内部数量无明显差异.而蚁科个体数从Dis_0到Dis_20逐渐增加,但无显著差异,有研究发现杂食性蚁科作为广布种无栖息地偏好且敏感度较低[33-35].蜘蛛目个体数从Dis_0到Dis_20也逐渐增加,同样无显著差异,Downie等研究了英格兰北部牧场交错带上的蜘蛛,认为微气候和潜在的竞争效应比栖息地结构对蜘蛛分布的影响更重要[36].有研究同样发现蜘蛛的个体数在距林间道路不同距离间无显著变化,但作者认为产生这种现象的原因目前很难解释[37].
3.2 群落多样性
在该研究中全部地表节肢动物Shannon-winner多样性指数在Dis_0最高,在Dis_5最低,在 Dis_20指数低于Dis_0但却高于Dis_5,即多样性指数向距离道路较远的样线反而略有升高,均匀度指数具有相同变化,优势度指数则呈相反的变化趋势.Dis_20所处地势相对较低,邻近调查期间存有积水的区域,这可能限制了地表节肢动物进一步向森林内部的扩散[38],从而导致地表节肢动物群落物种多样性在距离道路较远样线反而升高.虽然有研究表明,未铺设的道路对爬行动物的分布会产生深远的影响[39],但不同等级和不同交通状况的道路经过不同区域对自然生态系统产生的影响程度、范围、持续时间也有所差异[40-41],且确定某些生境特征对动物分布的影响程度是困难的,因为它们与其他非生物和生物因子之间存在着各种协同作用[39].由于该实验环境因子调查不足,因此不能较好的解释道路边缘地表节肢动物多样性受哪些因子调控,且应加强对其他月份的调查.
3.3 Beta多样性
Beta多样性不仅可以说明不同地段之间物种组成的差异,也可以直接表征环境的异质性[42],该研究中全部地表节肢动物在Dis_0样线上beta多样性最小,在Dis_20样线上群落beta多样性最大;步甲科则呈现相反特征;而葬甲科由于捕获数量较少,其beta多样性在三条样线上具有一致性.在距道路边缘不同距离样线间全部地表节肢动物、步甲科和葬甲科的 beta 多样性均具有显著差异(P<0.05).该文并未分析地形因子和空间过程等对地表节肢动物beta多样性的调控机制,相关研究还有待于进一步开展.分解beta多样性后得到全部地表节肢动物和步甲科群落的beta多样性均主要由空间物种转换组成,物种集群镶嵌的贡献很小,与高梅香等对丰林地表甲虫beta多样性分析结果一致[43],而葬甲科正相反.表明全部地表节肢动物的群落组成变异、步甲科的物种组成变异几乎都是由物种替换引起,即主要由本地类群(物种)之间较高的转换或替换引起[44].物种集群镶嵌都非常小,说明三条样线间物种缺失并不显著,这在一定程度上排除了道路及限制扩散对这些集群的重要调控作用,而葬甲科由于捕获个体数、物种数极少,从而导致beta多样性无显著变化且主要由物种集群镶嵌组成.由于该实验研究路段极少有车辆通过,因此不存在道路交通造成的空气污染、噪音污染及重金属污染等影响[45-46],但未铺设路面的道路足以改变森林内部结构且构成气候差异[47].而地表节肢动物具有个体小、数量大、种类多等特点,因此受土壤环境变化的影响较大[48-49],相关研究有待进一步开展.
4 结论
该研究共捕获地表节肢动物2662只,23类群,包括步甲科335只22种,葬甲科71只3种,其中步甲科、蚁科和蜘蛛目为优势类群.地表节肢动物个体数、类群数和多样性指数在距离道路边缘0~20 m空间范围内差异不显著,但随着距离道路边缘越远其总体呈升高趋势,Beta多样性结果进一步表明距道路边缘不同距离间地表节肢动物有较强的空间物种转换特征.基于这些结果可以推测该研究中未铺设路面的林区道路对其边缘地表节肢动物群落未造成太大影响.
猜你喜欢
大自然探索(2024年1期)2024-03-19 19:01:03
区域治理(2023年6期)2023-03-02 14:34:54
中国农学通报(2022年32期)2022-11-28 05:01:16
科学大众(中学)(2021年3期)2021-05-17 17:07:01
——以北长山岛为例
海洋开发与管理(2020年9期)2020-10-13 13:31:22
现代农业科技(2020年10期)2020-06-04 09:13:31
江苏农业科学(2019年5期)2019-09-02 14:01:46
课外生活(小学1-3年级)(2018年10期)2018-11-19 07:26:10
广东农业科学(2017年5期)2017-08-29 10:37:31
西南农业学报(2016年4期)2016-05-17 05:42:09
