
NAC转录因子参与调控竹笋木质化
2020-03-07 03:58单雪萌高志民
世界竹藤通讯 2020年1期
单雪萌 高志民
(国际竹藤中心 北京 100102)
原 文 出 处
随着生活水平的提高,人类对木材的需求迅速增长,然而木材供应却在逐渐减少,因此需要寻找替代资源。与多数木材相比,竹类植物生长速度快、产量高,具有良好的韧性和较高的强度,竹材的开发利用可以有效缓解木材供应紧张和木材数量不足的问题。次生细胞壁的形成和加厚在植物生长发育、木质化及木材形成过程中发挥着重要作用。木质化的次生细胞壁是最环保、最具成本效益的可再生资源,可以提高木质纤维素的生物量。目前,植物次生细胞壁倍受关注,转录调控在次生细胞壁生物合成中起关键作用,许多研究揭示了次生细胞壁生物合成在分子水平上的调控,表明植物具有调节次生细胞壁生物合成的特定转录开关,例如许多转录因子通过调控次生细胞壁成分(例如纤维素、半纤维素和木质素)的合成来参与次生细胞壁的形成。
NAC(NAM,ATAF和CUC)家族是植物特异性转录因子家族之一,其成员在多种生物过程中发挥重要作用,包括衰老、激素信号传导以及次生细胞壁的生物合成。在次生细胞壁的形成过程中,3个NAC转录因子(NST1、NST2和NST3/SND1)作为主要调节开关在纤维素、半纤维素和木质素3个组分的合成中发挥着重要功能。拟南芥(Arabidopsisthaliana)和水稻(Oryzasativa)中的NST1和NST2控制了花药内皮的次生壁增厚,同时许多其他NAC基因在次生细胞壁生物合成中也具有重要作用,例如与维管相关的NAC-DOMAIN1-7(VND1-VND7),VND基因的过表达使得次生细胞壁合成相关基因的表达上调,从而增加了细胞壁的厚度。棉花(Gossypiumhirsutum)NAC基因GhFSN1作为正向调控因子参与次生细胞壁生物合成,已经发现棉花中与纤维相关的NAC基因和次生细胞壁生物合成的基因之间存在着共表达关系。在控制次生细胞壁生物合成的基因调控网络中,除NAC转录因子外,还有MYB转录因子和许多其他转录因子等成员。与次生细胞壁生物合成相关的NAC基因的功能在整个植物界都是相对保守的,然而NAC在竹子中的功能仍不清楚。
竹子是最重要的非木材森林资源之一,被认为是新兴的重要木质纤维素生物质能源。毛竹(Phyllostachysedulis)属于禾本科竹亚科,与世界上大多数生态和经济竹种一样可用于生产竹材、艺术品、纸张,笋还可以食用。毛竹生长速度快,可在一个半月达到20 m左右,其木质化程度随着笋高度的增加而持续增加。在毛竹笋快速生长过程中次生细胞壁变厚和木质化,这对提高竹子的材性性能和应用起着重要作用。在拟南芥、水稻、谷子(Setariaitalica)和棉花中已经鉴定出许多与次生细胞壁合成相关的NAC转录因子基因,并且同源基因在不同物种之间具有不同的基因结构和功能。但是关于毛竹中与次生细胞壁相关的NAC转录因子研究很少。毛竹基因组草图和基因组数据库(BambooGDB)的完成有助于探索毛竹NAC基因的特定结构和功能特征。本研究的目的是对毛竹中NAC转录因子进行全面了解,揭示NAC转录因子参与次生细胞壁生物合成的分子机制。
1 材料与方法
1.1 毛竹NAC基因的鉴定
以模式植物拟南芥和水稻中NAC基因作为种子序列,利用植物转录因子数据库Plant TFDB(http://planttfdb.cbi.pku.edu.cn/index.php)获取NAC基因的同源序列,利用在线保守结构域分析软件逐一分析每条序列的保守结构域,剔除保守结构域不完整的序列,保留具有完整NAM结构域的氨基酸序列,并进行下一步研究分析。
1.2 基因结构、蛋白鉴定和序列分析
利用在线软件ProtParam(http://web.expasy.org/protparam/)对获得的蛋白的分子量和等电点等基本理化性质进行分析,使用Plant-mPLoc在线网站(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)对蛋白的亚细胞定位进行预测,同时利用在线网站psRNATarget(http://plantgrn.noble.org/psRNATarget/analysis)对毛竹中NAC基因的中miRNA的靶点进行预测。使用GSDS 2.0(http://gsds.cbi.pku.edu.cn/)分析NAC家族成员的基因结构,运用在线软件MEME(http://meme.nbcr.-net/meme/intro.html)对毛竹NAC的保守蛋白基序进行分析。
1.3 进化分析和功能预测
从网站TAIR(http://www.arabidopsis.org/)下载AtNACs序列,用于和PeNACs进行序列比对和系统进化分析。使用MEGA 6.0程序(http://www.megasoftware.net/mega.php)中的Clustal W进行多序列比对,并构建系统发育树。根据系统树中PeNACs和已知功能的AtNACs的进化关系预测毛竹中NAC的功能。
1.4 毛竹NAC基因的组织特异性表达分析
为了研究毛竹NAC基因的组织表达特异性,从NCBI Short Read Archive(SRA)下载毛竹叶、鞭、根、20 cm笋、50 cm笋、早花期花序和晚花期花序的转录组数据,获取其中NAC基因的RPKM值,用以表示基因的表达丰度。为方便统计,对每个表达数值取以2为底数的对数(Log2),使用Matrix2png(http://www.chibi.ubc.ca/matrix2png/)绘制基因表达热图。
1.5 毛竹NAC基因的共表达网络和GO分析
使用BambooNET数据库(http://bioinformatics.cau.edu.cn/bamboo/)对参与次生细胞壁生物合成的PeNACs进行共表达分析。转录组数据包括在不同发育阶段的鞭、根、笋、叶鞘和芽。对于共表达列表中的候选基因,利用GO分析来揭示PeNACs在次生细胞壁生物合成中的作用,并通过TBtools进行可视化。随后的GO分析在同一平台上进行,p值和FDR(错误发现率)分别低于0.01和0.05。另外,与PeNACs共表达的PeMYBs序列从PLAZA(https://bioinformatics.psb.ugent.be/plaza/)和BambooNET网站下载。
1.6 植物材料和cDNA合成
实验用毛竹源自江西南昌(28°45′58″N、115°45′39″E,海拔约400.0 m)野外,选择生长良好植株,分别采集不同生长阶段竹笋(笋高1.0、2.0、4.0、6.0和8.0 m)基部为样品,每个样品至少3次重复。样品经液氮处理后,存储于-80 ℃冰箱中,用于提取RNA。另外,取相同样品固定在FAA中保存于4 ℃冰箱,用于树脂切片。
使用TRIzol试剂(Invitrogen,美国)对毛竹笋进行总RNA提取。所有样品至少进行3个生物学重复。重组DNA酶I(Takara,日本)用于去除每个样品中残留的DNA。通过琼脂糖凝胶电泳验证总RNA的完整性,并通过分光光度法测定总RNA的纯度和浓度(Nanodrop 2000)。使用反转录试剂盒(Promega,美国)合成第一链cDNA。使用前将cDNA产物稀释5倍。
1.7 实时定量PCR分析
根据PeNACs与AtNACs之间的系统发育关系,筛选出与次生细胞壁生物合成相关的PeNACs。基于PeNACs和正向共表达的PeMYBs的多重序列比对,使用Primer Premier 5.0软件设计特异性引物。使用Roche LightCycler©480 SYBR Green 1 Master试剂盒(Roche,美国)在qTOWER2.2系统(Analytik Jena,德国)上进行qRT-PCR实验。反应体系(10.0 μL)∶5.0 μL2× SYBR Green 1 Master,0.8 μL cDNA,0.2 μL正反向引物(10 μM)和4.0 μL ddH2O。使用2-ΔΔCT方法,以PeNTB作为内参基因统计基因表达,qRT-PCR实验进行3次生物学重复。
1.8 组织化学分析方法
将FAA固定液中的毛竹笋样品进行包埋处理,经过不同浓度配比的聚乙二醇(Polyethylene glycol, PEG)逐级脱水后,包埋于PEG 4000中,用旋转切片机(Leica RM2165)进行切片,厚度为10 μm,用0.05%的甲苯胺蓝(Toluidine Blue O, TBO)对展片的切片于80 ℃染色3 min,并拍照观察。
1.9 毛竹NAC基因转录自激活活性分析
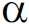
1.10 统计分析
统计分析使用IBM SPSS软件(版本21.0),用3次生物学重复结果进行均值和标准差分析。显著性差异用*(P<0.05)和**(P<0.01)表示。
2 结果与分析
2.1 PeNACs基因的鉴定与蛋白理化性质分析
利用拟南芥和水稻同源基因序列,从毛竹基因组数据库中经BLAST分析,共鉴定获得94个均具有完整保守结构域的NAC家族成员,依次命名为PeNAC1~PeNAC94。通过ProtParam工具,对PeNACs编码蛋白质的长度、分子量及等电点等理化性质进行分析。结果表明,PeNACs的氨基酸长度为217~694 aa,对应分子量为25201.17~76916.46 Da,理论等电点为4.55~10.85,亚细胞定位预测94个PeNACs均被定位在细胞核上。
2.2 PeNACs基因结构及其编码氨基酸的保守基序分析
基因结构分析表明,94个PeNACs聚类在11个分支(S1-S11),每个分支的成员数量为6~11个,内含子数量为0~8个,其中含有2个内含子的PeNACs数量最多有54个,含有3个内含子的有14个,含有4个内含子的有10个。在一个分支上的PeNACs具有相似的外显子—内含子基因结构,每个分支内含子的数量与系统进化关系和分支的分类是一致的,例如S9分支中8个PeNACs均含有8个内含子。然而也有些PeNACs例外,如聚类在S5分支的PeNAC79不含有内含子。PeNACs的基因结构差异明显,具有多样性,具体表现在内含子的大小和位置存在较大差异。
利用MEME软件对PeNACs基序进行分析,共鉴定了8个保守基序,命名为Motif 1~Motif 8,大部分保守基序都位于NAC蛋白的N末端,表明了这些保守基序对PeNACs的功能是非常重要的。聚集在一个分支的PeNACs具有相似的基序组成,例如S10家族的成员主要包含6个基序(motif1、motif2、motif3、motif5、motif6、motif7),S1家族的成员包含8个基序(motif1-motif8),表明了相同家族的PeNACs可能有相似的功能。
2.3 系统进化分析
为了确定PeNACs与拟南芥的NAC蛋白之间的进化关系,对毛竹和拟南芥的NAC氨基酸序列进行了多重比对,并利用MEGA 6.0软件构建了系统发育树。结果显示,NAC成员被分成16分支,毛竹PeNACs聚类在11个分支(C1-C10和C15),每个分支包含1~18个成员,进化树结果表明了一些NAC在毛竹与拟南芥中进化上具有较近的亲缘关系,例如AtNAC97和PeNAC55;AtNAC36、PeNAC51和PeNAC63。聚类在C1和C10家族的AtNACs与木质素的沉积和次生细胞壁的合成相关,推测与之聚类的PeNACs可能具有相似的功能。
2.4 PeNACs基因编码蛋白的功能预测
拟南芥中NAC转录因子的功能已经被证实,通过与AtNACs的比较可以预测PeNACs的功能。结果显示,拟南芥与毛竹NAC成员共聚类在16个亚家族(C1—C16),其中11个为毛竹与拟南芥所共有,5个为拟南芥所特有。有65个PeNACs聚类在6个具有功能注释的亚家族,29个PeNACs聚类在5个功能未知的亚家族,这65个PeNACs聚类在5类功能中,第1类包含2个亚家族(C1和C10),其主要通过木质素的生物合成和沉积来调控次生细胞壁的合成;第2类、第3类和第4类都包含1个亚家族,第2类(C2)与序列特异性DNA结合有关;第3类(C4)在根尖分生组织的形成和器官边界的建立中起着重要作用;第4类(C10)参与了植物调控和信号转导反应;第5类包含2个亚家族(C7和C8),主要参与植物衰老和非生物胁迫,例如高盐和干旱胁迫。这些结果表明了PeNACs功能的多样性,并且可能在毛竹生长发育过程中发挥着重要作用。
2.5 PeNACs组织特异性分析
利用转录组数据对PeNACs在毛竹不同组织中的表达模式进行分析。结果显示:94个PeNACs在毛竹不同组织和不同发育时期均有着不同程度的表达,然而表达丰度存在着明显的差异。94个PeNACs均至少在2个组织中检测到表达,63个PeNACs在所有的组织中均检测到表达,表达丰度为0.028~961.932。一些PeNACs基因在一些特异的组织中具有高的表达量,例如S7、S8和S9家族的基因在叶片和花序中具有较高的表达量,但在其他4个组织中表达量较低,尤其在根中表达量最低。同时,12个PeNACs(S1家族的PeNAC20、PeNAC40和PeNAC84;S3家族的PeNAC41和PeNAC60;S5家族的PeNAC71;S7家族的PeNAC6、PeNAC24和PeNAC34;S8家族的PeNAC70和PeNAC78以及S11家族的PeNAC2)几乎在所有检测到的组织中均有较高的表达量,S3家族的基因具有较高的表达量,但是S6家族的基因在7个组织中的表达量较低。
2.6 共表达网络分析
通过毛竹共表达网站,基于78个组织的转录组数据,对C1和C9亚家族中15个与次生细胞壁合成相关的PeNACs构建基因共表达网络,分别鉴定出与之共表达基因236个和160个。根据GO分析显示,共表达基因的功能和相关的生物途径主要富集在木质素分解代谢和纤维素合成这2个与次生细胞壁合成相关的生物过程,同时C1亚家族的共表达基因与次生细胞壁合成关系更加密切,说明这些基因可能参与次生细胞壁的合成途径。通过对共表达网络的构建,能够预测NAC转录因子可能与其它基因共同协调来参与毛竹次生细胞壁的合成。此外,7个PeMYBs基因与7个PeNACs基因呈现正向共表达关系,这些PeMYBs的上游2.0 kb启动子均含有NAC的结合元件(SNBE),SNBE由(T/A)NN(C/T)(T/C/G)TNNNNNNNA(A/C)GN(A/C/T)(A/T)组成,为19 bp的不完全回文序列,由此表明,这些PeNACs能够通过结合PeMYBs启动子中的SNBE元件来调控PeMYBs的表达。
2.7 PeNACs中miRNA靶点预测
据报道,许多miRNA能够参与次生细胞壁的生物合成,miRNA能够通过与相应的miRNA配对来切割靶基因,起到负调控的作用。用在线网站psRNATarget预测PeNACs中miRNA的靶点,由于水稻与毛竹亲缘关系较近,使用水稻miRNA文库作为参考对调控PeNACs的miRNA进行分析。结果显示,在94个PeNACs中共预测出315种miRNA,具有563个miRNA靶点,其中miRNA类型最多的是PeNAC5(91个),类型最少的是PeNAC25(3个)。另外,每个基因可能受到多种miRNA的调控,而一些PeNACs含有相同的miRNA靶点,例如16个PeNACs具有miR164靶点,11个PeNACs具有miR397靶点。15个与次生细胞壁合成相关的PeNACs中有3个(PeNAC36、PeNAC42和PeNAC45)是miR164的靶基因。由此表明,并不是所有具有相似功能的基因都由相同的miRNA调控,而相同的miRNA也可以调控具有不同功能的多个基因。
2.8 不同高度笋中PeNACs的表达模式分析
为了解15个与次生细胞壁相关的PeNACs的功能,以NTB基因作为内参基因,对PeNACs在不同高度笋中的表达模式进行分析。结果显示15个PeNACs在不同毛竹笋生长高度中均呈现差异表达,且随着笋高度的增加,除了PeNAC37的表达持续上调外,其它基因均不同程度地先上升再下降。相比于1.0 m高笋中的表达量,PeNAC37在8.0 m高笋中的表达量上升了1 000倍。同时PeNACs具有不同的表达模式,PeNAC3、PeNAC32、PeNAC45、PeNAC56和PeNAC85等5个基因在4.0 m高的笋中表达量达到最高值;PeNAC1、PeNAC8、PeNAC36、PeNAC42、PeNAC73、PeNAC76、PeNAC81和PeNAC94等8个基因在6.0 m高的笋中达到最大的表达量;而PeNAC11在2.0 m高的笋中达到了表达量最大。除PeNAC11外,其它PeNACs达到最高基因表达量均超过1.0 m高笋中表达量的2倍,而PeNAC94最高表达为1.0 m高笋中的12 565倍。同时,这些基因达到最大值后下降程度有着一定的差异,9个PeNACs(PeNAC1、PeNAC3、PeNAC8、PeNAC32、PeNAC73、PeNAC76、PeNAC81、PeNAC85和PeNAC94)的表达有一个明显的下降(超过最大值的70%),5个PeNACs(PeNAC11、PeNAC36、PeNAC42、PeNAC45和PeNAC56)仅下降了约20%,而且3个PeNACs(PeNAC36、PeNAC42和PeNAC45)上调的表达模式与miR164的表达模式恰好相反,表明miR164可能在毛竹笋生长发育过程中参与PeNACs的表达调控。此外,qRT-PCR结果显示PeMYBs与其共表达的PeNACs具有相似的表达趋势。另外,毛竹笋木质化程度随着笋高度的增加而逐渐上升,组织化学分析进一步证实了这一点。随着毛竹笋高度的增加,维管束的染色加深区域明显逐渐扩大,说明其木质化逐渐增强。
2.9 转录自激活活性分析
对与次生细胞壁合成相关的PeNAC8、PeNAC36和PeNAC73基因进行转录自激活活性分析,结果显示,所有含有目的基因质粒的酵母细胞均能在SD/-Trp单缺平板上生长并出现可见的白色菌落,在SD/-Trp/-His/-Ade/X-α-Gal选择培养基上只有阳性对照和含有目的基因质粒酵母细胞能够生长,并且变蓝色;相反阴性对照不能够生长。由此表明,PeNAC8、PeNAC36和PeNAC73均具有转录自激活活性。
3 讨论
NAC转录因子在许多生物过程中发挥着重要的作用,例如茎顶端分生组织的形成和维持、激素信号转导、细胞分化以及次生细胞壁的合成,然而毛竹中NAC家族基因并没有详细的研究,尤其是其成员在次生细胞壁形成和沉积过程中的作用尚未见报道。本研究通过对毛竹中NAC家族的基因结构、保守基序、进化关系、组织学分析、组织特异性表达以及转录活性等方面的研究,探讨其中NAC转录因子的调控机制。
3.1 PeNACs保守性和多样性
根据BLAST和Clustal W,所有的NAC都包含NAM结构域。NAC家族成员的数量在不同的物种中存在一定的差异,毛竹中共鉴定出94个PeNACs,比拟南芥、水稻和烟草中的数量都少。在NAC基因家族中,基因长度、预测的蛋白质分子量和蛋白质等电点之间差异很大,但基因结构是相对保守的。同一家族中的NAC成员可能具有相似的外显子/内含子结构和位置,表明这些成员亲缘关系较近并且可能具有相似的功能,这与先前的研究是一致的。然而,同一家族中的成员之间存在一些差异,例如外显子的大小、相对位置和数量,例如S11家族中的PeNAC81具有5个外显子,而其他家族成员具有3~4个外显子,而且其长度明显长于其他成员,这一结果可以通过进化过程中基因片段的剪接或插入从而产生新的基因功能来解释。另外,基序检测证实了同一家族中的大多数NAC蛋白具有相同的基序,这证明这些蛋白具有保守的功能,这与次生细胞壁相关NAC的功能可能在整个植物界中是保守的观点是一致的。
3.2 同源基因具有相似的功能
先前的研究表明,毛竹和水稻具有较近的进化关系,但是水稻中的大多数NAC基因尚未得到功能验证,拟南芥中的NAC基因功能已经得到了很好的研究,因此根据拟南芥的功能可以预测PeNACs的功能。系统进化树显示,毛竹和拟南芥共聚集成16个分支,根据与拟南芥NAC的同源性进行预测,结果显示在一个分支中的PeNACs的功能具有多样性,例如C4分支的AtNAC31、AtNAC92和AtNAC98不仅与植物衰老和顶端分生组织形成有关,而且与器官边界的建立有关。另外,具有相似功能的PeNACs不一定聚类在一个分支,例如与次生细胞壁形成和木质化合成相关的PeNACs分别聚类在C1和C10,也有一些分支由于拟南芥中相关功能没有研究而没有功能注释,这些分支功能需要进一步研究确认。
3.3 PeNACs基因表达模式的多样性
基因的表达是生物学功能的重要表现形式之一。为了进一步验证PeNACs的组织特异性表达,利用毛竹转录组数据分析发现,S3家族的成员在7个组织中均具有较高的表达量,表明它们在毛竹生长和发育过程中发挥着重要的作用;相反,S6家族的成员在7个组织中表达量均较低,可能因为它们在其他未检测到的组织中发挥着功能;一些PeNACs在叶片和花序中检测到较高的表达量,但在20 cm和50 cm高早期的毛竹笋中的表达量较低,说明它们可能主要参与细胞的分化和伸长而不是木质化的过程。与次生细胞壁合成相关的15个PeNACs在不同高度笋中的表达表明,与1.0 m高笋中相比,它们具有相似的表达变化,进一步证明它们在调控次生细胞壁合成过程中的保守性,这与前人的研究是一致的,同时组织化学结果也支持了随着笋高度增加,木质化程度加深,基因表达上调的结论。
3.4 次生细胞壁生物合成的复杂性
研究表明,植物具有调节次生细胞壁生物合成的特定转录开关,这些开关主要是NAC和MYB转录因子家族成员,NAC转录因子是第一层次的主要开关。本研究中,综合表达分析表明15个与次生细胞壁合成相关的PeNACs都随着笋高的增加而上调,其中PeNAC37是唯一具有持续增加的表达趋势的基因,其他14个PeNACs的表达量在达到峰值之后开始下降,而且不同PeNACs达到峰值时毛竹笋的高度是不同的,表明它们在时间和空间上的功能可能是不同的。miR164的表达趋势与和次生细胞壁合成和木质素合成相关的3个PeNACs的表达趋势相反,这与miRNA及其靶基因之间具有负向共表达是一致的,表明在毛竹次生细胞壁合成过程中可能存在一个miRNA-NAC调控网络。
另有一些研究表明,MYB转录因子作为次生细胞壁合成过程中的第2层调控开关,基于NAC-MYB调控网络来实现对参与次生细胞壁合成的下游结构基因的调控。本研究中,在7个PeMYBs启动子中含有NAC结合元件,这些PeMYBs与7个PeNACs呈现正向共表达,并且它们在不同高度的毛竹笋中具有与共表达的PeNACs相似的表达模式,这表明PeNACs可能正向调控PeMYBs的表达,这与先前的研究是一致的。而且PeMYB3与AtMYB46和AtMYB83同源,这表明在次生细胞壁合成过程中,PeMYB3作为第二层调控开关,与AtMYB46和AtMYB83功能是相似的。这些结果表明,在次生细胞壁的合成中NAC-MYB的调控是一个渐进的调控过程。此外,在共表达网络中共预测出396个与15个PeNACs共表达的基因,这表明毛竹次生细胞壁生物合成过程中可能涉及转录因子、结构基因和miRNA等复杂网络的调控。但要揭示该网络对竹材材性的调控机制仍需大量的研究工作。
4 结论
竹子是很好的木材替代品。NAC转录因子可以调节次生细胞壁的生物合成和木质化沉积,从而影响竹材材性,因此研究竹子NAC转录因子的功能对揭示竹材形成机制具有重要意义。在毛竹中有94个PeNACs,其表达模式具有多样性,其中15个PeNACs的表达量随着笋高度的增加均明显上调,这与竹笋木质化程度加深相一致。本研究对深入研究PeNACs参与次生细胞壁生物合成调控,解析其在竹材形成中的生物学功能具有重要价值。
猜你喜欢
中国农业科学(2022年11期)2022-06-27
安徽农业科学(2022年1期)2022-02-14
蔬菜(2021年2期)2021-11-27
江苏农业学报(2021年2期)2021-06-30
广东农业科学(2020年9期)2020-11-10
三农资讯半月报(2020年18期)2020-10-14
意林·少年版(2020年13期)2020-08-02
东坡赤壁诗词(2020年2期)2020-06-04
阅读与作文(小学高年级版)(2019年3期)2019-04-20
湖北农业科学(2017年10期)2017-06-22
