
Identification and modulation of expression of a TNF receptor superfamily member 25 homologue in grass carp(Ctenopharyngodon idella)
2020-02-28 09:45XingxingChengXinyuJiangYunjieSongJingduoGaoYujieXueZeinabHassanQianGaoJunZou
Aquaculture and Fisheries 2020年1期
Xingxing Cheng,Xinyu Jiang,Yunjie Song,Jingduo Gao,Yujie Xue,Zeinab Hassan,Qian Gao,∗,Jun Zou,∗∗
a Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources,Ministry of Education,Shanghai Ocean University,Shanghai,201306,China
b International Research Center for Marine Biosciences at Shanghai Ocean University,Ministry of Science and Technology,Shanghai,201306,China
c National Demonstration Center for Experimental Fisheries Science Education,Shanghai Ocean University,Shanghai,201306,China
Keywords:
ABSTRACT
1.Introduction
The tumor necrosis factor superfamily(TNFSF)of ligands and receptors(TNFRSF)are fundamental for many biological processes such as immunity,inflammation and hematopoiesis(Aggarwal,Gupta,&Kim,2012).TNF was first discovered during attempts to treat mice with cancer with injections of serum pre-challenged withSerratia marcescenspolysaccharide(Aggarwal et al.,2012).TNFRSF25 was discovered during trials to isolate proteins that interact with TNF receptor 1(TNFR1)in the 1990's(Schreiber & Podack,2013).To date,many members of the TNF superfamily have been identified in vertebrates.There are approx.19 TNFSFs and 29 TNFRSFs in humans(Wiens &Glenney,2011).The TNFRSF can be divided into three main groups:1)the TNF receptor associated factor(TRAF)binding domain containing receptors,2)the death domain(DD)motif(approx.80 amino acids)containing receptors,and 3)the decoy receptors(Bodmer,Schneider,&Tschopp,2002).The TNFRSFs contain 1-4 cysteine-rich domains(CRDs)in their ectoplasmic region and each CRD consists of three disulfide bonds with a core motif of CXXCXXC(Tafalla&Granja,2018).The DD is present in a number of TNFR associated proteins and related molecules such as death receptor(DR3-6)and Fas,which are primarily involved in signaling cell death(Wang et al.,2001).Among the TNFRSFs,the isoform TNFRSF25 shares the highest sequence homology with TNFRSF1(Bittner,Knoll,& Ehrenschwender,2017b).TNFRSF1 is ubiquitously expressed whilst TNFRSF25 expression is restricted primarily to lymphocyte-rich tissues such as the spleen,thymus,kidney and colon(Al-Lamki et al.,2003;Schreiber & Podack,2013).Migone et al.have demonstrated that the TNF-like protein 1A(TL1A),also known as the TNF ligand superfamily 15(TNFSF15),acts as the sole ligand for TNFRSF25(Migone et al.,2002).TNFRSF25,once activated by TL1A,activates several cellular signaling pathways mediated by the nuclear factors(NF)-κB,c-Jun N-terminal kinase(JNK)and p38 mitogen activated protein kinases(MAPKs),leading to cell apoptosis or death(Bittner,Knoll,& Ehrenschwender,2017a).TNFRSF25 mediated signaling can be suppressed by decoy receptor 3(DcR3)via competition with TL1A(Siakavellas & Bamias,2015).
Homologues of TNFRSF members have been reported in fish(Secombes,2016).In addition,the functions of TNFRSF1a/TNFR1 and TNFRSF1b/TNFR2 have also been established(Grayfer & Belosevic,2009;Xu,Nie,Sun,& Chang,2010;Zhang et al.,2016).Recombinant goldfish TNFα2 induces the expression of TNFRSF1b/TNFR2 and downregulates its own expression in primary macrophages.In goldfish and grass carp,soluble receptors containing the extracellular region interact with TNFαand antagonize TNFαactivities.However,research into fish TNFRSF25 is scarce.In the present study,an orthologue of TNFRSF25(termedCiTNFRSF25)was identified in grass carp(Ctenopharyngodon idella)and its phylogenetic relationship with other known TNFRSF25s were determined.Expression analysis showed that theCiTNFRSF25 was differentially expressed in tissues of healthy fish and could be modulated after exposure toFlavobacterium columnare(F.columnare).The results suggest that theCiTNFRSF25 is involved in immune response to bacterial infection.
2.Materials and methods
2.1.Fish maintenance and tissue sampling
Grass carp(120±12 g),were purchased from a local fish farm(Shanghai,China).All fish were acclimatized at 25±2℃ for 14 days in aerated tanks and fed with commercial diet twice daily until 24 h before the challenge experiment.Fish were anaesthetized with 0.05%MS-222 prior to experimental procedures.All experiments complied with the national regulations and guidelines of Shanghai Ocean University for use of animals for research(License No.,SHOU-DW-2019-003).
For tissue sampling,fish were anaesthetized with MS-222 and killed by cutting the spinal cord.Tissues were collected immediately,snap frozen in liquid nitrogen and stored at-80℃ until use.
2.2.Challenge experiment
The pathogenic G4strain ofF.columnarewas provided by Dr Haixie Xie,the Institute of Hydrobiology,Chinese Academy of Sciences(Wuhan,China)(Xie,Nie,& Sun,2010).Experimental fish were randomly divided into two groups(12 fish per group)and were injected intraperitoneally(i.p.)withbacterial suspension(1μL/g body weight,1×107cfu/mL)or PBS(1μL/g body weight).Three fish were sampled from each group and killed at 12,24,48 and 72 h post injection(hpi).The head kidney and spleen were collected for total RNA extraction.
2.3.Production and purification of recombinant interleukin(rCiIL-4/13B)
TheCiIL-4/13B was cloned into the expression vector pEHISTEVb using the restriction enzymesNcoI andHindIII.BL21 Star(DE3)competent cells were transformed with the pEHISTEVb-CiIL-4/13B plasmid.The recombinantCiIL-4/13B protein was expressed as inclusion bodies after induction of the transformed BL21 cells with 1 mM IPTG(SIGMA,USA).The inclusion bodies were dissolved in 6 M guanidine hydrochloride and gradually added into the refolding buffer(100 mmol/L Tris-HCl,2 mmol/L EDTA,400 mmol/L L-arginine-HCl,0.5 mmol/L oxidized glutathione and 5 mmol/L reduced glutathione,pH 7.4).After incubation for 20 h at 4℃,the refolded protein solution was centrifuged at 13,000 rpm for 20 min,concentrated by filtration,and purified by HPLC chromatography using a Superdex 75 10/60 column(GE Healthcare Life Sciences,USA).The protein fractions were collected in 1.5 mL tubes and checked by SDS-PAGE.The fractions containing protein of the expected size of recombinantCiIL-4/13B protein was mixed and quantified using a Pierce® BCA Protein Assay Kit(Thermo Fisher Scientific,USA)according to the manufacturer's protocols.
2.4.Isolation of primary leucocytes and stimulation by interleukin(IL)-4/13B
Primary leucocytes were collected from the head kidney of healthy grass carp using Percoll gradient centrifugation,which was carried out as follows.L-15 medium(2% FBS,0.1% heparin,100 U/mL penicillin and 100 U/mL streptomycin)was prepared and pre-cooled before the experiment.The head kidney was transferred to a 70μm nylon mesh and gently pushed through the mesh.The cell suspension was loaded onto a 51% Percoll gradient(GE Healthcare Life Sciences,USA)and centrifuged at 400×gfor 30 min at 4℃ without braking.The cells were collected at the Percoll/medium interface and resuspended in L-15 medium(0.1% FBS,0.1% heparin,100 U/mL penicillin,100 U/mL streptomycin).Finally,the cells were washed twice with L-15 medium(0.1% FBS,100 U/mL penicillin,100 U/mL streptomycin)and centrifuged at 400×gat 4℃ for 10 min.
Following isolation,the cells were cultured in cell culture medium(RPMI with 10% FBS,100 U/mL penicillin and 100 U/mL streptomycin)in 6 well plates(1.5×106cells/well)at 28℃ for 4 h in a 5%CO2incubator.The rCiIL-4/13B protein was added to the wells at a final concentration of 20 ng/mL and at day 3,the same amount of rCiIL-4/13B was added again.The control cells were treated in parallel with an equal volume of protein buffer containing 20 mmol/L His-HCl and 50 mmol/L NaCl.The cells were collected at 12 h,24 h,48 h,72 h and 7d for quantitation of mRNA levels ofCiTNFRSF25 by qPCR as described below.
2.5.RNA isolation and synthesis of cDNA
All tissues were quickly frozen in liquid nitrogen after sampling and kept in a-80℃ freezer.Total RNA was extracted using TRIZOL(SIGMA,USA)according to the manufacturer's instructions.The firststrand cDNA was synthesized using a Prime Script™RT reagent Kit with gDNA Eraser(Perfect Real Time)(TaKaRa,Japan)for cloning and quantitative real-time PCR(qPCR).The synthesized cDNA was stored at-20℃ until use.
2.6.Cloning of the full-length TNFRSF25 sequences
The partial cDNA sequence of grass carp TNFRSF25 was obtained by analyzing the whole-genome sequence database(http://www.ncgr.ac.cn/grasscarp/)(Wang et al.,2015).Based on the partial cDNA sequences,the 5′and 3′untranslated region(UTR)were obtained by Touchdown PCR and Nest PCR.The primers and adaptors used to obtain the full-length cDNA sequence ofCiTNFRSF25 in this study are listed in Table 1.The amplicons were checked by electrophoresis and extracted from the agarose gel using a Gel Extraction Kit(OMEGA,USA),ligated into the pMD19-T vector(TaKaRa,Japan)and transformed into theE.coliDH5αcompetent cells.Positive clones containing inserts of correct size were screened by PCR and three randomly selected clones were sequenced(Sangon shanghai,China).Following the 5′and 3′RACE,the full-length coding sequence was confirmed by PCR using a pair of primers located at the 5′and 3’untranslated region(UTR)(Table 1).
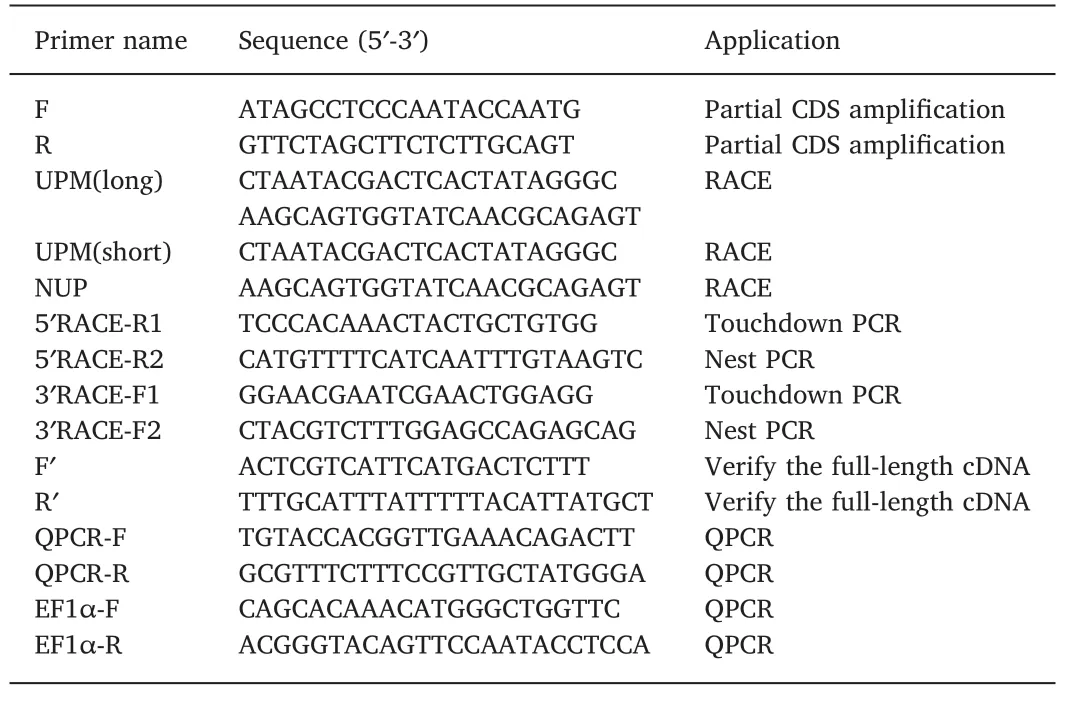
Table 1Primers used in this study.
2.7.Sequence analysis of TNFRSF25
The full-length sequence of TNFRSF25 cDNA was analyzed using the BLAST program in the NCBI database(http://www.ncbi.nlm.nih.gov).The deduced amino acid sequence was analyzed using the programs listed on the ExPASy website(http://www.us.expasy.org/tools/)(Wilkins et al.,1999).The amino acid sequences of vertebrate TNFRSF25s were obtained from the NCBI database(www.ncbi.nih.gov)and used for multiple alignment using the ClustalW 2 program(Chenna,2003).The protein domains were predicted using the Simple Modular Architecture Research Tool(SMART)(http://smart.embl-heidelberg.de/).The signal peptide and the transmembrane region were predicted using the SignalP4.1(http://www.cbs.dtu.dk/services/SignalP/)and TMHMM 2.0 Server(http://www.cbs.dtu.dk/services/TMHMM)respectively.A neighbor joining phylogenetic tree was constructed using the MEGA 5.0 programme with p-distance parameters and complete deletion of gaps.The size of grass carp TNFRSF25 introns and exons was obtained by comparing the genomic and corresponding cDNA sequence.Gene synteny data of human,mouse and zebrafish were taken from the Ensembl database and that of grass carp obtained by analyzing the genome sequences.
2.8.Real-time PCR quantification of gene expression
Quantitative real-time PCR(qPCR)was carried out to examine gene expression.The primers forCiTNFRSF25 and elongation factor-1α(EF1α)are listed in Table 1.The qPCR reaction was set up as follows:5μL of SYBR Premix(Yeasen shanghai,China),4.6μL of the cDNA and 0.2μL of forward and reverse primer(10μmol/L).The qPCR was performed in a Bio-Rad Light Cycler 96 Real-time detection system(Roche,Switzerland)using SYBR Premix under following conditions:a preincubation at 95℃ for 30 s,followed by 40 cycles of 95℃/5 s,62℃/30 s,72℃/10 s,and a melting reaction at 95℃/10 s,75℃/60 s,95℃/1s.Each sample was run in triplicate.In these experiments,EF-1αwas used as the internal control to normalize gene expression.Fold change in expression was calculated by comparing the normalized mRNA levels ofCiTNFRSF25 of treatment group and that of the corresponding control group.
2.9.Data analysis
The qPCR data were analyzed using one-way analysis of variance(One-Way ANOVA)and a LSD post hoc test with the SPSS package 20.0(SPSS Inc.,Chicago,IL,USA).P<0.05 andP<0.01 were considered significantly different between groups.
3.Results
3.1.Amino acid sequence and phylogenetic analysis of CiTNFRSF25
The cDNA sequence ofCiTNFRSF25 was obtained and deposited in the NCBI database(Accession number:MK651506).The full-length cDNA sequence ofCiTNFRSF25 was 1420 bp in length,and contained a 5′-UTR of 60 bp,a 3’-UTR of 208 bp and an open reading frame of 1152 bp(Fig.1).TheCiTNFRSF25 cDNA encoded a peptide of 383 aa with a calculated molecular weight of 43.1 kDa and an isoelectric point of 8.0.A polyadenylation signal(AATAAA)site was located 15 bp upstream from the poly A tail(Fig.1).TheCiTNFRSF25 contained a signal peptide of 22 aa(Met1-Cys22)and a transmembrane region of 23 aa(Leu173-Phe195).A characteristic TNFR DD was identified between Val291and Leu371(Fig.1).Further analysis revealed that theCiTNFRSF25 contained two conserved potential N-glycosylation sites(Asn55-Ser57,Asn242-Thr244)and a sumoylation site(Ala346-Glu349).In addition,there was a single typical TNFR domain,also called the cysteine rich domain(CRD),ranging from 48 to 89 aa,unlike mammalian and avian TNFRSF25s that contains two or three CRDs(Fig.3).

Fig.1.Nucleotide and deduced protein sequence of CiTNFRSF25.The start and stop codon of the open reading frame are in bold.The polyadenylation signal(AATAAA)is underlined.The putative signal peptide is in Italics.Two potential N-glycosylation sites(grey),cysteine rich domains(CRDs)(green),death domain(DD)(blue)and transmembrane domain(yellow)are highlighted.The conserved cysteine residues in the CRD are circled.The predicted sumoylation site is boxed.
A protein sequence alignment of TNFRSF25s was generated.As shown in Fig.2,the CRD and DD were highly conserved in humans,mouse,chicken,zebrafish,crucian carp and grass carp.The CRD of grass carp contained six cysteine residues(Cys48,Cys63,Cys66,Cys69,Cys81and Cys89)and three disulfide bonds(Fig.2).To better understand the evolution of TNFRSF25,an unrooted phylogenetic tree was constructed using the Neighbor-Joining method(Fig.4).It was apparent thatCiTNFRSF25 was clustered with fish TNFRSF25s,also annotated as TNFRSF1ALs in some species in the NCBI and Ensembl database.Fish TNFRSF25s/TNFRSF1ALs formed a sister clade with TNFRSF25s from mammals and birds with a bootstrap value of 83%.The strong grouping demonstrates the homologous relationship of TNFRSF25s between fish and higher vertebrates.

Fig.2.Multiple sequence alignment of TNFRSF25 proteins.The identical aa residues are shaded in black and similar aa residues in grey.The signal peptide(SP),cysteine rich domain(CRD),transmembrane domain(TM)and death domain(DD)are indicated.Homo sapiens:AAO63495.1;Mus musculus:AAH17526.1;Gallus gallus:XP_423315.4;Danio rerio:XP_009295056.1;Carassius auratus:XP_026097489.1.
TheCiTNFRSF25 gene organization was obtained by analyzing the genomic and cDNA sequence.It spanned approx.6.8 kb and was longer than its human and mouse counterparts and was organized into ten exons and nine introns,the same number as found in humans and mouse(Fig.5).TheCiTNFRSF25 gene was located in Scaffold_1000001 and the surrounding genes included CCDC30,UBR4 and RCC2 which were also present in the TNFRSF25 gene locus in humans,mouse and zebrafish(Fig.6).
As shown in Table 2,the deduced aa sequence of theCiTNFRSF25 gene shared 72.1% sequence identity with crucian carp TNFRSF25 and 20%-70% sequence identity with other vertebrate TNFRSF25s.The CRD domain ofCiTNFRSF25 shared 57.1% and 54.8% aa identity with the zebrafish and crucian carp homologues,respectively(Table 3).However,only 21.4% sequence identity was found between grass carp and rainbow trout homologues(Table 3).Table 4 shows that the DD ofCiTNFRSF25 was also highly conserved,sharing 97.3% sequence identity with the DD of zebrafish and crucian carp TNFRSF25.
3.2.Tissue distribution
The expression levels ofCiTNFRSF25 were determined in nine tissues of healthy grass carp using qPCR.As shown in Fig.7,theCiTNFRSF25 transcripts were constitutively expressed in all the tissues examined,including brain,skin,gills,intestine,thymus,spleen,head kidney,trunk kidney and liver.The gills exhibited the highest expression followed by brain,spleen and liver.The lowest expression was detected in the head kidney.
3.3.Gene expression in response to F.columnare
The expression levels ofCiTNFRSF25 were investigated in grass carp after infection byF.columnare.As shown in Fig.8,theCiTNFRSF25 was significantly down-regulated(P<0.01)in both head kidney and spleen at 24 hpi.In the head kidney,theCiTNFRSF25 transcripts increased at 12 hpi,decreased significantly at 24 hpi and thereafter returned to normal level.However,in the spleen,the expression ofCiTNFRSF25 hardly changed after infection except for a transient down-regulation at 24 hpi.
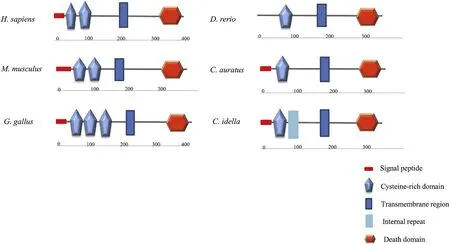
Fig.3.The protein domains of TNFSFR25 predicted using SMART program.
3.4.Gene expression of CiTNFRSF25 in primary head kidney leucocytes after stimulation with rCiIL-4/13B
To determine the effects of rCiIL-4/13B on theCiTNFRSF25 expression,the rCiIL-4/13B protein was purified.Primary head kidney leukocytes were isolated and the effects of rCiIL-4/13B on theCiTNFRSF25 expression examined.The cells were incubated with the rCiIL-4/13B protein for 12 h,24 h,48 h,72 h and 7 d.Fig.9 shows that theCiTNFRSF25 transcripts were up-regulated at 12,24 and 48 hpi and returned to normal level at 72 h.The results demonstrated that theCiTNFRSF25 expression was modulated in kidney leukocytes by rCiIL-4/13B.

Fig.4.Phylogenetic tree analysis of TNFRSF25s from Ctenopharyngodon idella and other species.The tree was constructed using the MEGA 5.0 program with the Neighbor-Joining method.Numbers at tree nodes represent the percentage of bootstrap values after 10,000 replicates.
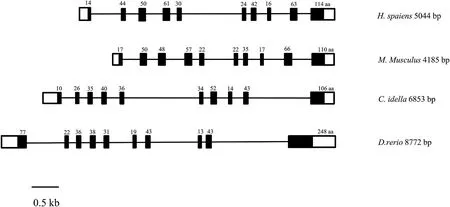
Fig.5.Genomic organization of the TNFRSF25 gene from human,mouse,grass carp and zebrafish.The exons are shown from 5′to 3′end.Black and open boxes represent coding regions and untranslated regions,respectively.The size of the coding region in each exons is numbered.
4.Discussion
In this paper,a homologue of TNFRSF25(termedCiTNFRSF25)was identified in grass carp.Bioinformatics analysis revealed that it possessed the typical structure of other TNFRSF25 molecules,consisting of a cysteine-rich domain in the extracellular region and a death domain in the intracellular region.Moreover,the gene synteny ofCiTNFRSF25 resembled that of human,mouse and zebrafish,suggesting they shared a common ancestor during evolution.TheCiTNFRSF25 was constitutively expressed in tissues and was modulated by rCiIL-4/13B in primary head kidney leucocytes and by bacterial infection in head kidney and spleen.
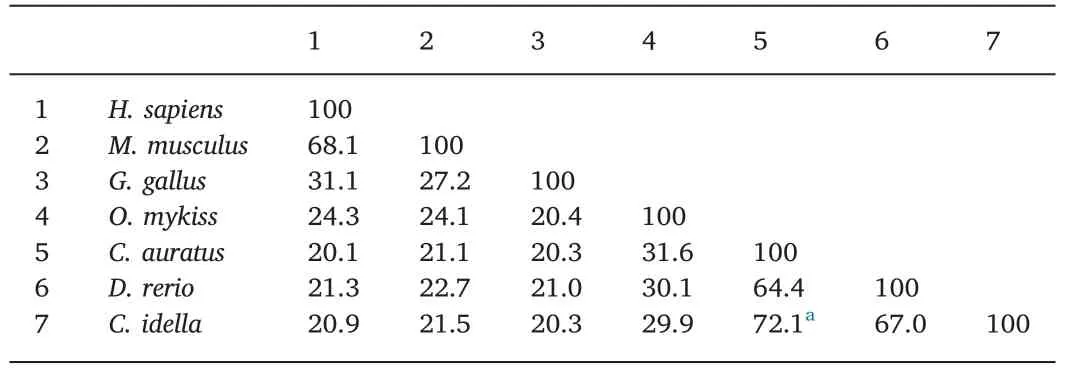
Table 2Percentage of amino acid identity(%)of TNFRSF25.
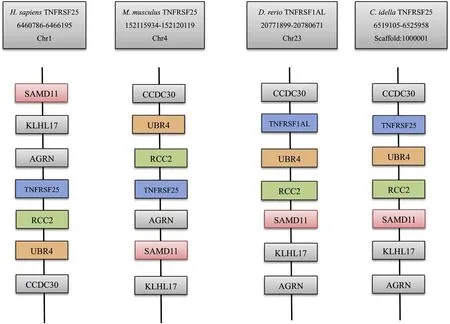
Fig.6.Comparative synteny analysis of TNFRSF25 genes.The genomic data of human,mouse,and zebrafish were retrieved from the Ensembl database.The CiTNFRSF25 data were analyzed using the sequence deposited in the genome database.Gene symbols were taken from the Ensembl database.
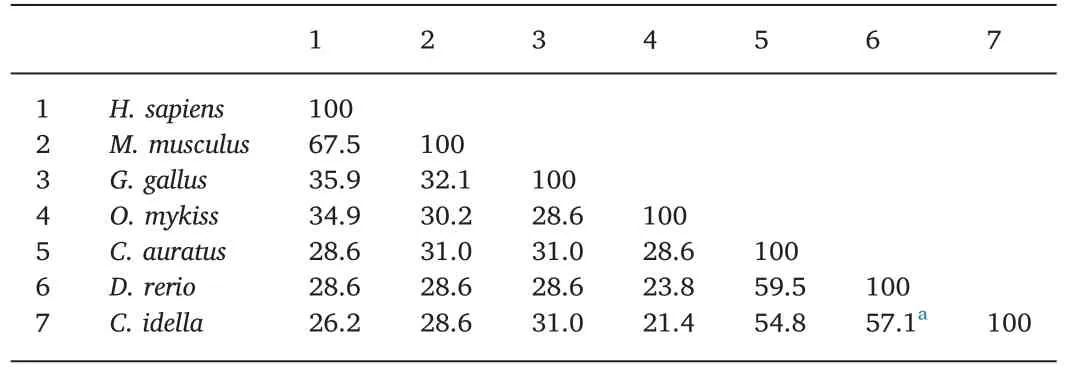
Table 3Percentage of amino acid identity(%)of CRDs in TNFRSF25.
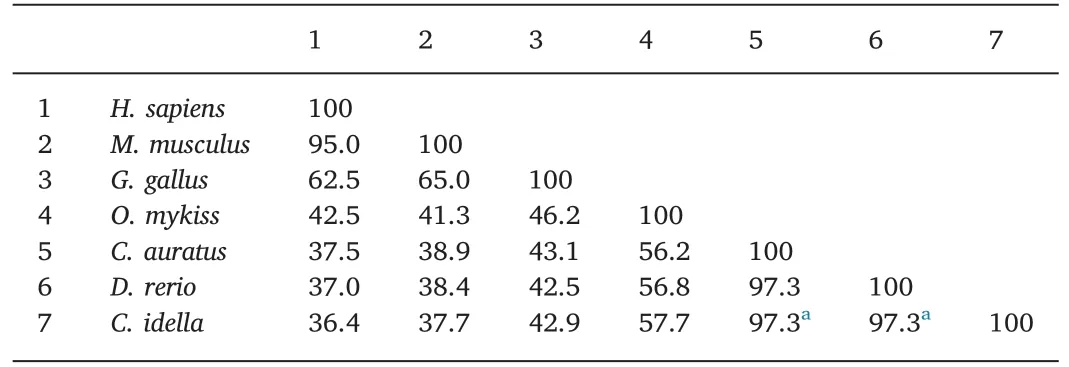
Table 4Percentage of amino acid identity(%)of DD in TNFRSF25.
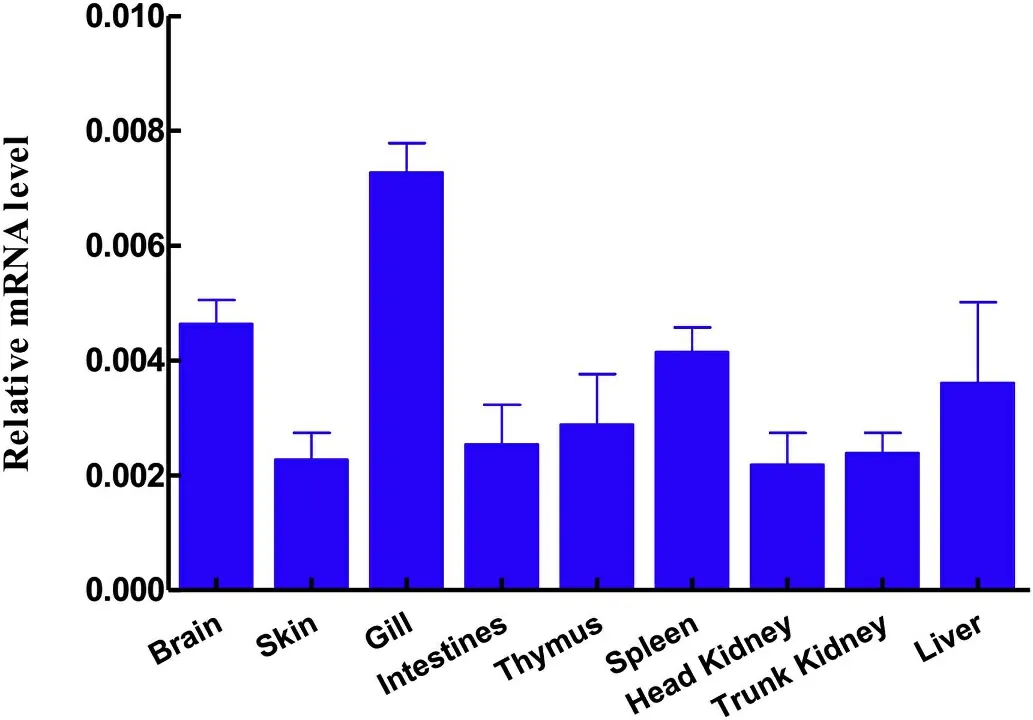
Fig.7.Tissue expression of CiTNFRSF25.The expression levels of CiTNFRSF25 in the brain,skin,gills,intestine,thymus,spleen,head kidney,trunk kidney and liver were determined by qPCR and are presented as relative expression to EF1α(N=3).
The TNFRSF25 is known to be a potent inducer of cell apoptosis and necroptosis in mammals and has similar functions to Fas and TNFRSF1(Bodmer et al.,1997).The functions of TNFRSF25 are mediated via the death domain in the intracellular region,and involves recruitment and activation of downstream signal molecules(Bossen et al.,2006;Ware,2003).Once activated by its ligand(TNF-like molecule 1A)TNFRSF25 initiates two distinct downstream signaling cascades(Siakavellas,Sfikakis,& Bamias,2015),1)interaction of domains with the TNFRassociated death domain protein(TRADD),and 2)the TNFR-associated factor(TRAF)and receptor-interacting(RIP)proteins(Micheau &Tschopp,2003),both elicit pro-inflammatory signals or cell death(Bittner & Ehrenschwender,2017).CiTNFRSF25 has a well conserved death domain which is located between 291 aa and 371 aa(Fig.2),suggesting it may have similar functions to the mammalian counterpart.In fact,cellular signaling molecules interacting with TNFRSF such as TRAF proteins have been reported in fish(Tafalla & Granja,2018).
Phylogenetic tree analysis revealed that theCiTNFRSF25 formed a clade with fish TNFRSF1ALs.This fish TNFRSF25/TNFRSF1ALs clade formed an expanded clade with mammalian TNFRSF25s supported by a high bootstrap value of 83%(Fig.3),indicating they are evolutionarily related.The orthologous relationship was also supported by the conserved gene synteny of genes neighboring the TNFRSF25 gene in fish and mammals(Fig.6).Therefore,we propose that the TNFRSF1AL genes annotated in fish should be re-named,TNFRSF25,to avoid confusion in the future.
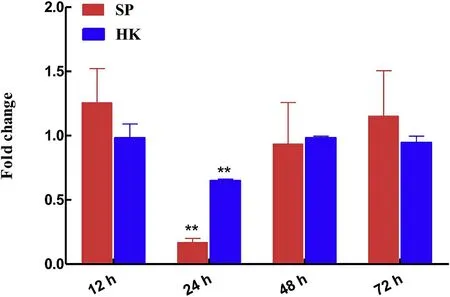
Fig.8.The expression levels of CiTNFRSF25 in head kidney and spleen stimulated with PBS or F.cloumnare for 12,24,48 and 72 hpi.CiTNFRSF25 expression levels were calculated relative to that of EF1αgene(internal control)and presented as fold change relative to the control(=1).The data are shown as mean±S.E(N=3).**P<0.01 or*P<0.05 are considered significantly different.
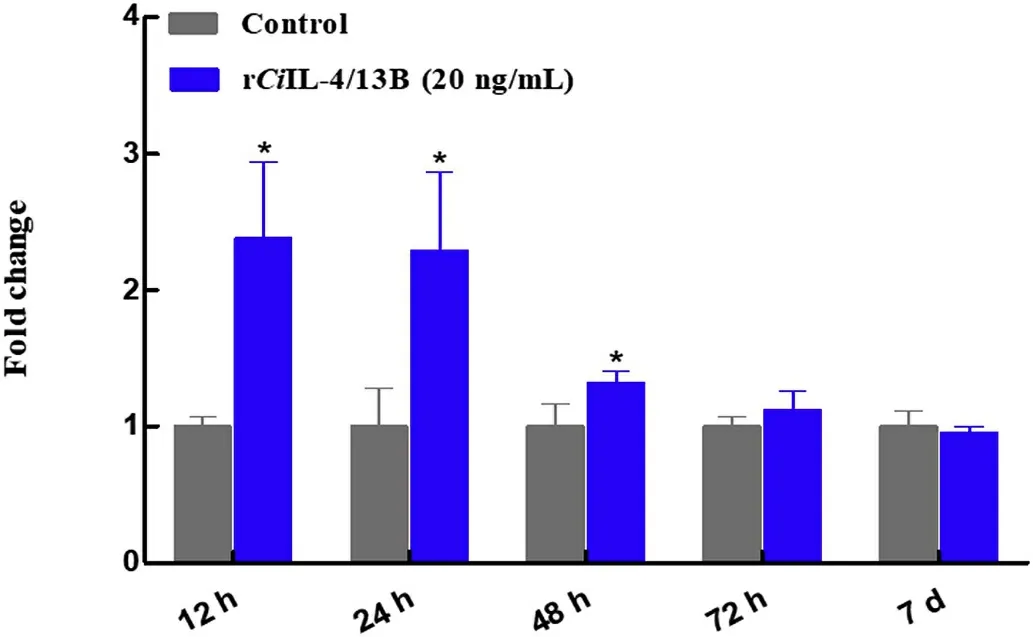
Fig.9.The expression levels of CiTNFRSF25 in primary head kidney leucocytes after stimulation with rCiIL-4/13B.The head kidney cells were isolated from 3 individual fish,and were cultured in 25 cm2 flasks for 4 h and then stimulated with 20 ng/mL of rCiIL-4/13B or protein buffer.At 12 h,24 h,48 h,72 h and 7 d,the cells were harvested for qPCR analysis.The expression levels of CiTNFRSF25 were calculated relative to that of EF1αgene(internal control)and presented as fold change relative to the control(=1).The data are shown as mean+S.E(N=3).*P<0.05 is considered significantly different.
In mammals,TNFRSF25 was expressed mainly in lymphocyte-rich tissues upon activation with inflammatory cytokines(Meylan et al.,2008).In the present study,CiTNFRSF25 mRNA transcripts were detected in a broad range of tissues,but at different levels(Fig.7).For example,the expression of theCiTNFRSF25 gene was markedly higher in gills,brain and spleen than in other tissues(Fig.7).It has previously been shown that TNFRSF25 is restricted to lymphocytes and is highly expressed by T reg cells(Schreiber et al.,2010;Twohig et al.,2012).However,other studies demonstrated that the expression of TNFRSF25 was less restricted(Cretney et al.,2003;Schreiber&Podack,2013).We found that 24 h after i.p.administration ofF.columnare,the expression levels ofCiTNFRSF25 were decreased in both spleen and head kidney,the major systemic immune organs in teleost fish(Fig.8).The inflammatory response toF.columnareinfection was probably caused by migration of immune cells from the spleen and head kidney to the site of infection to eliminate invading pathogens.Additionally,CiTNFRSF25 was up-regulated byCiIL-4/13B in primary kidney leukocytes(Fig.9).This suggests that cytokines such as IL-4/13B may regulate TNFRSF25 mediated responses by altering the numbers of receptor molecules on the cell surface.
To summarize,this is the first report on the TNFRSF25 gene in teleost fish.The results indicate that the TNFRSF25 mRNA was constitutively expressed in a broad range of tissues and could be modulated by infection withF.columnare.The predicted sequence motifs ofCiTNFRSF25 suggests that it may have similar functions to mammalian TNFRSF25s.
 Aquaculture and Fisheries2020年1期
Aquaculture and Fisheries2020年1期
- Aquaculture and Fisheries的其它文章
- Probiotics in shellfish aquaculture
- Identification and histopathological and pathogenicity analysis of Aeromonas salmonicida salmonicida from goldfish(Carassius auratus)in North China
- Comparison of juvenile growth of wild and hatchery-raised olive flounders Paralichthys olivaceus as examined by otolith microstructure
- The effects of on-farm produced feeds on growth,survival,yield and feed cost of juvenile African sharptooth catfish(Clarias gariepinus)
- Biofloc technology(BFT):Adjusting the levels of digestible protein and digestible energy in diets of Nile tilapia juveniles raised in brackish water