
紫云英季土壤固氮微生物对外源碳氮投入的响应
2020-02-27 03:52杨璐曾闹华白金顺周兴周国朋高嵩涓聂军曹卫东
中国农业科学 2020年1期
杨璐,曾闹华,白金顺,周兴,周国朋,高嵩涓,聂军,曹卫东,
紫云英季土壤固氮微生物对外源碳氮投入的响应
杨璐1,2,曾闹华2,白金顺2,周兴3,周国朋1,2,高嵩涓4,聂军5,曹卫东2,4
(1中国农业科学院研究生院,北京 100081;2中国农业科学院农业资源与农业区划研究所/农业农村部植物营养与肥料重点实验室,北京 100081;3湖南省农业科学院作物研究所,长沙 410125;4南京农业大学资源与环境科学学院,南京210095;5湖南省农业科学院土壤肥料研究所,长沙 410125)
【】分析不同外源有机物料(稻草、葡萄糖)及氮素投入对紫云英季土壤固氮微生物的调控作用,为我国南方紫云英-水稻轮作体系中秸秆还田及化肥减施增效提供支持。采用盆栽试验,共设7个处理,即CK(对照,不添加有机物料和氮)、稻草等量添加并配施不同量氮素(分别表示为Rs、RsN1和RsN2,对应C/N比分别为66、25和13);等秸秆碳量添加葡萄糖并配施不同量氮素(即Glc、GlcN1、GlcN2)、调整C/N比与秸秆添加相应处理保持一致。采集紫云英快速生长期土壤样品,利用Illumina Miseq PE300高通量测序和绝对定量PCR技术分析固氮功能基因及固氮微生物群落特征。单独添加外源秸秆或葡萄糖处理的土壤C/N与对照无明显差异,但增施氮肥后C/N比呈下降趋势,GlcN2处理土壤C/N显著低于对照;对于土壤速效养分,Rs和RsN1处理土壤NO3--N含量与CK类似,但RsN2处理明显增加了60%;而添加葡萄糖处理土壤NO3--N含量整体较高(增幅为35%—79%);稻草单独添加或与氮素配施对土壤速效磷含量无明显影响,添加葡萄糖处理则显著降低其含量,降幅为16%—24%,但不同氮水平之间无明显差异。不同处理土壤基因拷贝数为80.4×106—140.5×106g-1土,稻草和葡萄糖添加处理基因拷贝数较CK呈增加趋势,而增施氮素后有所下降。外源碳、氮添加导致固氮微生物α-多样性较CK整体降低,但α-多样性对氮素的响应因碳源种类而异,即稻草添加后增加氮素供应(RsN1和RsN2)处理土壤固氮微生物物种数目和Chao 1指数较Rs显著降低,降低幅度分别为6%—11%和13%—15%,而添加葡萄糖后增施氮肥则对α-多样性有一定促进作用。PCoA结果显示土壤固氮微生物群落结构主要因碳源种类差异而聚集为不同组别,受氮水平供应的影响相对较小。属水平上不同物种对外源碳、氮添加的响应存在明显差异:慢生根瘤菌()相对丰度最高,而外源碳、氮添加后较CK明显降低,且稻草添加处理的降低幅度(12.3%—19.7%)小于葡萄糖添加处理(31.6%—40.5%);第二优势菌属(地杆菌,r)对碳源添加的响应趋势与相反,与CK相比,葡萄糖添加处理相对丰度的增加幅度(170%—270%)明显大于秸秆添加处理(25.0%—54.6%)。同时,多元回归树、RDA及相关性分析表明土壤固氮微生物丰度、多样性和群落结构组成主要受土壤NO3--N和速效磷的影响较大。氮素供应对固氮微生物多样性和群落结构的调控作用受碳源种类(或秸秆和葡萄糖中碳源有效性差异)影响;同时,不同碳源添加后造成土壤速效磷含量的差异也可能是影响紫云英季土壤固氮微生物群落组成的重要环境因子。
固氮微生物;水稻秸秆;葡萄糖;氮;紫云英;
0 引言
【研究意义】紫云英(L.)一直以来是我国南方普遍种植的豆科绿肥,在水稻休闲期种植紫云英,能够充分利用光、水、热等资源;同时,利用其生长阶段生物固氮可补充土壤氮库,减少水稻季氮素投入[1]。据报道,紫云英体内氮素有78%来自生物固氮[2]。生物固氮是农田生态系统氮素投入的重要途径之一,据估计每年生物固氮量可达4 000— 7 000万t[3-4]。固氮微生物是土壤中参与生物固氮的重要成员,系统发育和代谢过程复杂而多样化[5-7],且对环境变化十分敏感[8]。维持土壤固氮微生物的多样性并充分发挥其固氮能力,增加绿色氮源投入,对农田化肥减施和农业绿色发展具有重要意义。【前人研究进展】固氮微生物多样性及群落结构的变化,能够较好地反应其生物固氮能力[9],但外源化肥、有机肥投入、秸秆还田等带来的土壤环境变化,都会对固氮微生物生长和功能发挥造成影响[10-13]。目前关于不同外源物料投入对土壤固氮微生物的促进或抑制作用均有报道。长期施用化肥能够显著影响固氮微生物丰度和群落组成[14]或并无明显作用[15]。由于土壤中多数参与N2固定的微生物属于化能异养型,外源有机物料添加能够为其提供大量碳源和能源,故一般认为有机物料添加对固氮微生物具有促进作用[16]。但因有机物料质量(C/N)不同,该影响效果各异,如农家肥、城市生活废弃物堆肥、水稻秸秆3种不同C/N比物料相比,农家肥C/N比相对较低,对土壤固氮酶活性的影响相对较小,比不施肥对照降低14%;城市生活垃圾堆肥氮含量相对较高,较大程度地抑制了生物固氮(降低50%);而高C/N比的稻草秸秆,则使土壤固氮酶活性比对照增加了40%[17]。稻草秸秆还田可以增强生物固氮能力[18-19]或增加固氮基因()丰度[20-21],可能与秸秆还田后短期内导致土壤速效氮含量降低有关。此外,底物有效性是导致微生物群落竞争的重要因子[22],微生物对不同碳源的利用能力也存在差异,如固氮厌氧菌对葡萄糖和纤维素添加响应不同[23]。【本研究切入点】上述研究结果表明,土壤固氮微生物对外源有机、无机物料添加或底物的数量及质量的响应存在差异。近年来,随着机械化生产进程的推进,我国南方紫云英-水稻轮作体系中,稻草和紫云英联合还田成为稻田重要的作物残体利用方式,二者代表了两种不同质量的有机物料投入;同时,化肥施用也能够调节还田物料的C/N比及有机物料的腐解过程。目前,关于不同有机无机物料投入对紫云英季土壤固氮微生物对的影响已有相关报道[24],但上述措施(如还田物料的碳源有效性及C/N比)对紫云英生长季土壤固氮微生物的调控机制尚不清楚。【拟解决的关键问题】拟通过盆栽试验,探讨紫云英生长期间土壤固氮微生物对不同碳源有效性的有机物料(秸秆和葡萄糖)配合氮素添加的响应规律,为我国南方紫云英-水稻轮作体系中秸秆和氮肥管理及豆科绿肥固氮提供理论依据。
1 材料与方法
1.1 试验设计
盆栽试验于2017年10月至2018年4月年在湖南省长沙市(湖南省土壤肥料研究所网室)进行。供试土壤采自稻田0—20 cm耕层,土壤类型为洞庭湖地区典型水稻土(河流沉积物发育而来的紫潮泥)。土壤基础理化性状为:pH (H2O) 8.00,土壤有机碳(SOC)18.7 g·kg-1,土壤全氮(TN)2.1 g·kg-1,速效磷19.0 mg·kg-1,速效钾84.9 mg·kg-1。
试验共7个处理,分别为:(1)对照CK,不添加外源有机物料、不施氮肥;(2)Rs,仅添加水稻秸秆,C/N = 66;(3)RsN1,添加等量水稻秸秆+低量氮肥,将底物调至C/N=25;(4)RsN2,添加等量水稻秸秆+高量氮肥,将底物调至C/N = 13;(5)Glc,等秸秆碳量添加葡萄糖,并加微量氮素调至底物C/N= 66;(6)GlcN1,等秸秆碳量添加葡萄糖+低量氮肥,将底物调至C/N=25;(7)GlcN2,等秸秆碳量添加葡萄糖+高量氮肥,将底物调至C/N=13。每个处理重复4次,完全随机排列。紫云英品种湘紫1号。试验所用稻草碳(C)、氮(N)、磷(P)和钾(K)含量分别为40%、0.60%、0.055%、2.07%。试验每盆装风干土10 kg,稻草添加量为40 g/盆,装盆时将秸秆剪碎至1—2 cm。葡萄糖添加量为40 g/盆,与秸秆等碳量添加,并补充氮素调至相应C/N比。所有处理等磷(P2O5)、等钾(K2O)添加。不同处理具体外源物料添加量如表1所示。
1.2 样品采集及指标测定
于2018年3月10日紫云英快速生长期取样,将去除紫云英植株后的土壤混合均匀。一部分4℃保存,待测完无机氮后,风干磨细用于其他土壤基本理化性状测定,其中NO3--N和NH4+-N采用1 mol·L-1KCl 浸提,AA3连续流动分析仪测定;土壤速效磷用0.5 mol·L-1NaHCO3浸提,钼锑抗比色法测定;土壤速效钾用1 mol·L-1醋酸铵浸提,原子吸收仪测定;土壤pH用快速pH测定仪测定(土水比=1﹕2.5),土壤全氮和有机质分别采用凯氏定氮法和K2Cr2O7氧化外加热法[25]。另一部分存于-80℃,用于土壤DNA提取。
1.3 DNA提取,PCR扩增和克隆
采集的土壤样品送至北京奥维森基因科技有限公司利用Illumina Miseq PE300平台进行测序分析。采用PowerSoil® DNA Isolation kit(MO BIO Laboratories, Inc., CA, USA)试剂盒,按操作步骤说明提取土壤DNA。用1%琼脂糖凝胶进行检测DNA提取质量和浓度。以稀释后的基因组DNA 为模板,根据测序区域的选择,使用带8-bp Barcode 的特异引物,进行PCR反应。上游引物为AAA GGY GGW ATC GGY AAR TCC ACC AC,下游引物为TTG TTS GCS GCR TAC ATS GCC ATC AT。PCR扩增程序:94℃预变性5 min后,94℃变性30 s,58℃退火30 s,72℃延伸60 s,循环32次,循环结束后72℃保持7 min,4℃条件下结束,收集数据。将同一样本3次PCR反应产物的混合物用2%琼脂糖凝胶电泳检测,AxyPrepDNA凝胶回收试剂盒(AXYGEN公司)切胶回收,Tris_HCl洗脱,2%琼脂糖电泳检测,QuantiFluor™-ST蓝色荧光定量系统(Promega公司)定量检测,最后按照每个样本的测序量要求,进行相应比例的混合。混合后的样品进行Miseq测序,测序数据已上传至NCBI SRA数据库(SRA accession: PRJNA542599)。
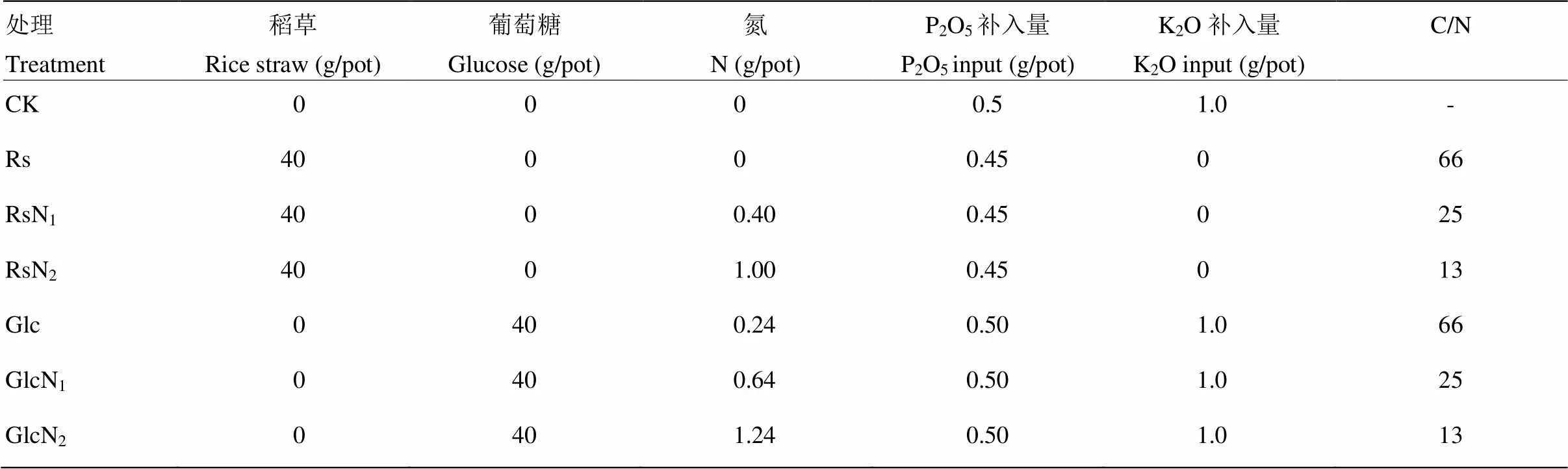
表1 各处理外源有机物料及氮磷钾添加量(每盆装10 kg风干土)
1.4 绝对定量PCR (qPCR)
本研究采用绝对定量PCR分析土壤DNA样品基因拷贝数。先用目的基因引物进行PCR扩增(引物序列同上),目的片段进行琼脂糖凝胶回收,回收的PCR产物进行TA连接,菌落PCR鉴定阳性克隆,并提取质粒作为绝对定量的标准品。再将标准品从101—105进行10倍梯度稀释,每个梯度取2 μL做模板建立标准曲线。测定样品时,标准品与未知样品同时进行PCR 循环,PCR反应体系配置如下:2×Taq Plus Master Mix 10 μL,10 mmol·L上游引物和下游引物各0.5 μL,加水至18 μL。加入2 μL DNA后,按以下程序进行PCR:95℃,30 s;40个PCR循环(95℃,5 s;60℃,40 s(收集荧光))结合标准曲线计算获得待测样品中基因拷贝数。
1.5 生物信息学分析
生物信息学分析等由北京奥维森基因科技有限公司完成,原始数据下机后,首先进行数据质控,通过序列拼接、过滤和去嵌合体后得到优化序列,然后进行OTU聚类及注释,其中OTU划分基于核苷酸序列97%相似性水平。
1.6 数据处理
不同处理对土壤理化性状、基因拷贝数、Alpha多样性的影响采用SAS package 9.1 (SAS Institute,Cary,NC,USA)进行ANOVA分析,LSD(<0.05)进行差异比较;土壤理化性状对nifH基因拷贝数的相对影响作用采用R中的“relweights”包[26]进行权重分析;土壤理化性状与功能菌属Spearman相关关系采用R中的“PerformanceAnalytics”包分析;群落结构差异及环境因子对群落结构影响的主坐标(PCoA(基于Bray-Curtis距离矩阵))和冗余分析(RDA)由R中的“Vegan”包完成;R中‘mvpart’包[27]用于多元回归树分析。做图软件为Microsoft Excel、Sigmaplot和R中“ggbur”包。
2 结果
2.1 外源碳、氮添加对土壤理化性状的影响
由表2可知,秸秆、葡糖糖和氮素添加各处理土壤pH与对照相比无明显差异。与对照相比,RsN2、Glc和GlcN1处理土壤有机质含量以及RsN2和GlcN2处理土壤全氮含量明显增加;尽管各处理土壤C/N与对照无明显差异,但增施氮肥后C/N比呈下降趋势。对于土壤速效养分,Rs和RsN1处理NO3--N含量与CK类似,但RsN2处理明显增加了60%;而添加葡萄糖处理土壤NO3--N含量整体较高(增幅为35%—79%);稻草单独添加或与氮素配施对土壤速效磷含量无明显影响,添加葡萄糖处理则显著降低16%—24%,但不同氮水平之间无明显差异。秸秆或葡萄糖添加对土壤速效K影响相对较弱,但随着氮素添加量的增加,速效钾含量呈下降趋势。

表2 外源C、N添加对紫云英快速生长期土壤理化性状的影响
表格中表格数据为平均值± 标准误,= 4;同一列不同字母表示处理之间差异显著(<0.05)
Data are means ± SE,=4. Means with no letter in common are significantly different among treatments at<0.05 level
2.2 固氮微生物nifH基因拷贝数及α-多样性
与CK 相比,Glc处理基因拷贝数增加了1.1倍,其他添加稻草或葡萄糖处理拷贝数也均一定程度增加,但差异不显著;增施氮肥后拷贝数呈下降趋势(表3)。对于固氮微生物α-多样性,稻草单独添加处理(Rs)有利于增加固氮微生物α-多样性,其中Chao 1指数增加明显;添加稻草后增施氮肥明显抑制了固氮微生物α-多样性,RsN1和RsN2处理chao 1指数比Rs处理分别降低12.5%和15.1%,物种数目降低6.5%和11.2%。添加葡萄糖处理与CK相比,固氮微生物α-多样性整体较低。与添加稻草处理相比,添加葡萄糖后增施氮肥固氮微生物α-多样性呈增加趋势,其中GlcN2与Glc相比增加幅度达显著水平(表3)。

表3 外源碳、氮添加对土壤固氮微生物nifH基因拷贝数及α-多样性的影响
数据为平均值±标准误(= 4)。图中不同字母表示各处理之间差异显著(<0.05)
Data are means±SE (=4). Means with no letter in common indicate significant differences among treatments (<0.05)
权重分析结果表明,所检测的土壤环境因子中,土壤速效氮、有机质、速效钾和C/N对基因拷贝数的相对影响作用大于其他理化性质(图1);多元回归分析表明,土壤NO3--N是导致不同处理之间α-多样性差异的主导因子(图2)。
2.3 土壤固氮微生物群落结构
基于Bray-cruits 距离的PCoA分析结果表明,不同碳、氮添加处理土壤固氮微生物群落聚集为4组(Group),其中Group 1包含CK、Rs和RsN2处理,Group 2包含RsN1处理,Group 3包含Glc和GlcN1处理,Group 4包含GlcN2处理(图3-A)。相似性分析(ANOSIM)表明,不同组别之间群落结构差异显著(=0.001)。群落结构组成同环境因子的冗余分析(RDA)结果表明,所检测的环境因子对土壤固氮微生物群落结构差异解释量为39.9%,RDA1和RDA2解释量分别为23.9%和4.9%;土壤环境因子中速效磷(=4.70,=0.001)和pH(=2.02,=0.05)的影响作用达到显著(图3-B)。
2.4 土壤固氮微生物优势功能菌属
所有处理土壤在属水平上共鉴定出160种固氮菌,包含26个优势功能均属(相对丰度大于1%),其中20个菌属对外源碳、氮添加响应明显,包括多种α-型、β-型、δ-型和γ-型变形菌和一种蓝藻细菌,如眉藻属(图4)。慢生根瘤菌()为最丰富菌属,CK中占23.6%,添加稻草处理后降低至19%—21%,而添加葡萄糖后明显降低至14%—16%,但受氮水平影响较小。秸秆或葡萄糖添加明显增加了和等菌属的相对丰度,其中秸秆添加处理相对丰度较CK平均增加了36.5%,而葡萄糖添加平均增加了2倍,增加幅度远远大于秸秆添加处理,且施氮量增加(C/N降低)相对丰度明显降低;也表现出类似的规律。此外,与CK比较,秸秆添加处理的相对丰度整体呈增加趋势,随氮素添加量增加而呈增加趋势,而葡萄糖添加后相对丰度降低了约22%。

图1 土壤理化性状对nifH基因拷贝数影响的权重分析
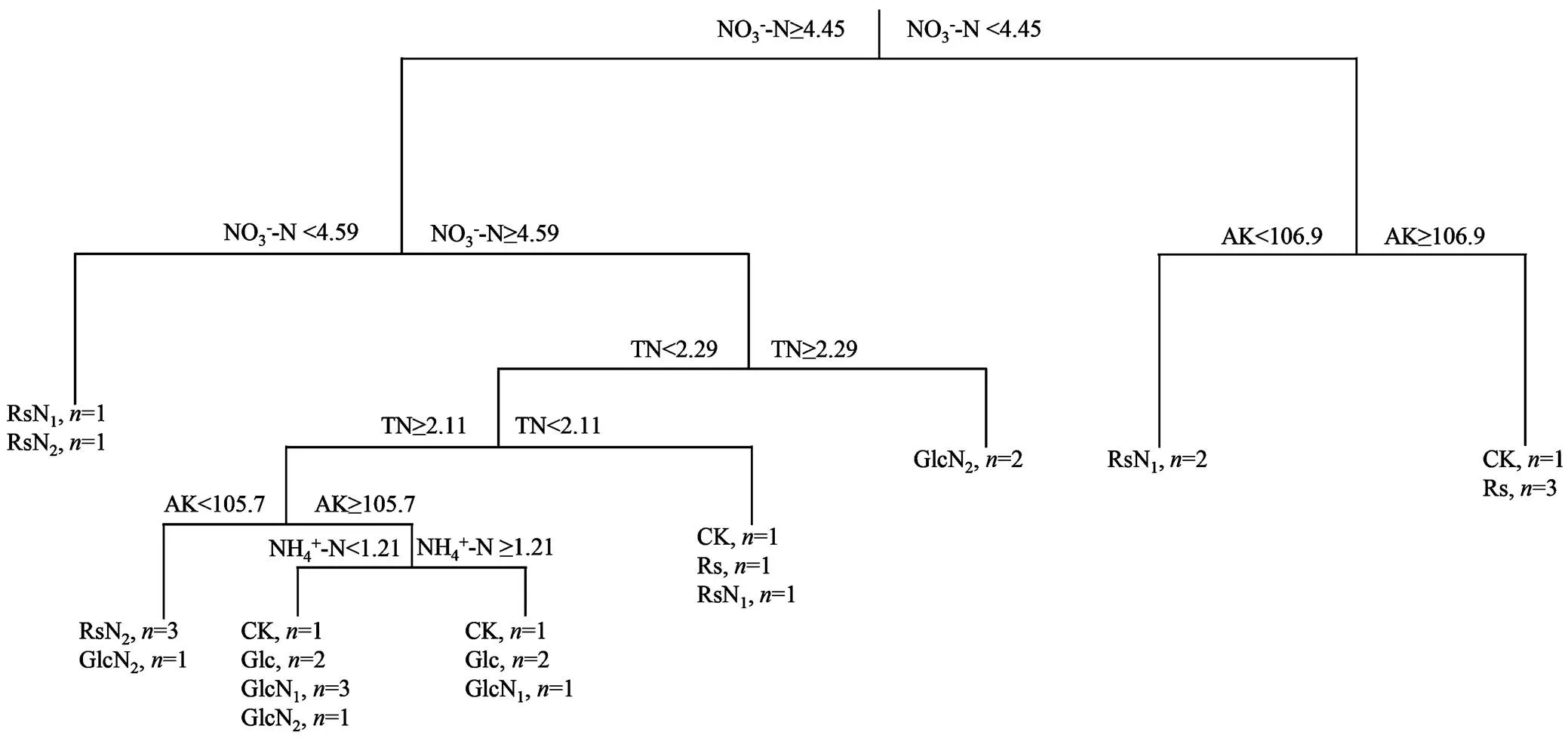
图2 固氮微生物α-多样性与土壤理化性状的多元回归树分析
相关性分析表明,属水平上大多数优势土壤固氮微生物相对丰度与土壤速效磷显著相关(图5)。、、和等优势功能菌属的相对丰度均与土壤速效磷呈显著正相关,而r等则与其呈显著负相关;土壤pH及C/N与属水平相对丰度的相关性趋势和速效磷类似,但仅有少数菌属达到显著水平。属相对丰度与土壤SOM、TN、NO3--N含量的响应也较为敏感,但趋势则与对速效磷的响应相反,如、和相对丰度与SOM显著负相关。属相对丰度与土壤速效钾和NH4+-N含量的相关性较弱(图5)。多元回归树分析进一步表明,稻草和葡萄糖添加处理后土壤固氮微生物属水平相对丰度的差异主要受速效磷驱动(图6)。

图3 土壤固氮微生物群落结构差异的 PCoA分析(A)及其与环境因子的RDA分析(B)
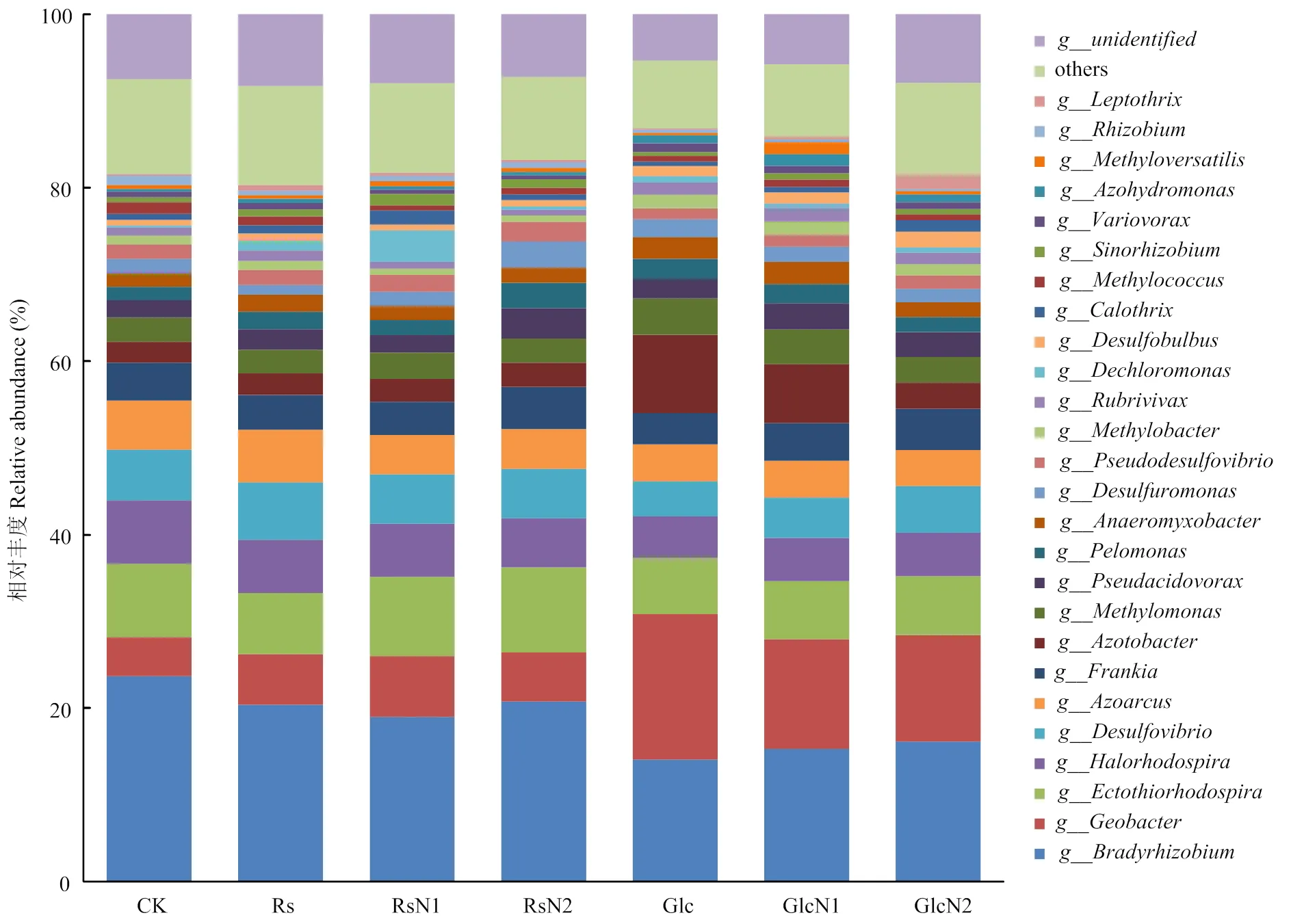
图4 不同处理土壤固氮微生物优势功能菌属相对丰度

图5 土壤固氮微生物优势功能菌属与土壤理化性状的Spearman相关性分析

图6 土壤固氮微生物优势功能菌属与土壤理化性状的多元回归树分析(MRT)
3 讨论
由于土壤中固氮微生物多为化能异养型,故外源有机物料添加能够为其提供大量碳源和能源,有利于生物固氮[16]。与此一致,本研究中秸秆和葡萄糖添加处理基因拷贝数呈增加趋势(表3);但氮素添加(即C/N降低)减弱了这种趋势,这可能与增加氮素有效性一定程度抑制固氮活性有关[24, 28]。与拷贝数相反,外源碳、氮添加降低了固氮微生物α-多样性(Rs单独添加处理Chao 1指数明显高于CK除外),但是α-多样性对氮素的响应因碳源不同(秸秆和葡萄糖)而表现不同,即增加氮素供应显著降低了秸秆处理中α-多样性,但添加葡萄糖后增施氮肥则一定程度增加了α-多样性(表3)。上述结果表明,碳源种类及C/N比是影响土壤固氮微生物α-多样性的关键因素。
尽管高C/N更有利于微生物固氮活性[17,29],但C/N比并不能代表添加物料对微生物生长所需C源和N源的数量及有效性[30]。底物有效性是导致微生物群落竞争的重要因子[22],很大程度决定了有机物料微生物活性和结构组成[31-32]。尽管不少研究指出,有机物料添加有利于促进基因丰度及固氮功能的维持[20-21,33],但有机物料对固氮微生物的影响可能也与有机物料的数量和质量相关[33-34]。LIN等[35]指出,花生、水稻和萝卜等植物残体投入对固氮微生物结构没有显著影响,但猪粪配施化肥后群落组成明显异于对照和单施化肥处理。WU 等[36]也指出,微生物对堆肥和生物炭的响应差异,一定程度上由生物炭添加处理中可溶性碳较低、不利于微生物利用所致。这些研究表明,微生物对不同碳源利用能力存在差异,因而对不同碳源的响应也有所区别。
本研究中PCoA结果表明,土壤固氮微生物群落因添加碳源不同(葡萄糖、秸秆)而聚集为不同组别(图3-A)。通常,革兰氏阳性菌倾向于利用相对复杂的碳底物[37],而革兰氏阴性菌则喜好利用简单结构的碳源[38]。葡萄糖较秸秆是一种更容易利用的C源,更容易被微生物吸收利用,微生物对其响应也可能更为敏感。与对照(CK)比较,葡萄糖添加处理相对丰度平均降低了35.8%,降低幅度明显大于添加秸秆处理(15.3%);而第二优势菌属相对丰度的增加幅度却远远大于秸秆添加处理(124.8%26.7%);同时,秸秆添加有增加相对丰度的趋势,而葡萄糖添加则表现为降低。其他研究也发现厌氧固氮菌对葡萄糖和纤维素的响应不一致[21],进一步表明秸秆和葡萄糖添加后,由于二者所含碳水化合物结构复杂程度悬殊,导致碳源有效性明显不同,进而影响土壤固氮微生物群落结构和组成。
外源物料投入对固氮微生物的影响,可直接通过添加的碳源、能源和养分实现,或通过影响土壤环境因子来间接调控。氮素是微生物生长和代谢所必须,施肥等措施能够提升土壤有效氮含量,但一般抑制土壤固氮酶活性[10-11];而氮素缺乏又可能限制微生物的生长[14,29]。本研究中外源碳氮添加导致土壤NO3--N增加或显著增加,无论在添加稻草还是葡萄糖处理中,增加氮素供应均导致土壤固氮基因丰度呈降低趋势(表3);但是α-多样性和优势功能菌属相对丰度对氮素添加的响应因碳源而异。如添加秸秆和葡萄糖后增加氮素供应,固氮微生物α-多样性响应规律完全相反(表3)。秸秆添加处理的相对丰度整体呈增加趋势,随氮素添加量增加而呈增加趋势,而葡萄糖添加后相对丰度降低了约22%(图4)。这些结果表明,氮素供应对固氮微生物多样性和群落结构的调控作用受碳源种类(或秸秆和葡萄糖中碳源有效性)影响。生物固氮是一个十分耗能的过程,固氮微生物需要消耗大量的ATP来完成生物固氮[39],因而磷养分的供应在固氮微生物的生长和功能发挥中起着着重要作用。本研究RDA分析、Mantel检验及相关性分析表明,土壤固氮微生物群落结构的差异与速效磷含量紧密相关;而PCoA结果表明土壤固氮微生物群落因葡萄糖添加和秸秆添加而明显聚集为不同组别,这与添加葡萄糖处理后土壤速效磷含量显著低于秸秆处理的结果相吻合(表2)。而添加秸秆处理组内,RsN1处理固氮微生物单独聚为一类,可能与该处理土壤速效磷含量较高有关(表2,图3-B)。同时,由于、等相对丰度与土壤速效磷显著正相关,而土壤等则与其呈负相关,因此,添加葡萄糖处理后相对丰度较秸秆处理明显降低,而相对丰度却大幅增加,表明土壤速效磷含量对固氮微生物群落组成具有较强的调控作用。其他研究通过研究不同形态磷素及其与葡萄糖添加对固氮微生物的影响发现,磷有效性决定了固氮微生物的总体固氮潜力,但不同固氮微生物的响应可能不同[40]。结合多元回归树分析,上述结果表明土壤速效磷含量的差异可能是不同碳源(稻草和葡萄糖)添加后驱动土壤固氮微生物属水平相对丰度的差异另一个重要环境因子[40](图6)。本研究中,各处理均为外源养分供应均为等磷、等钾量添加,但添加葡萄糖处理后土壤速效磷含量明显低于秸秆和对照处理,可能与添加葡萄糖促进了其他微生物繁殖,进而固持土壤速效磷有关。
此外,在我国南方紫云英-水稻轮作体系中,稻草还田及种植翻压紫云英是当前提升稻田土壤地力的主要措施之一。禾本科和豆科残体质量差异较大,二者还田或与氮肥配施后对土壤固氮微生物丰度和群落结构的促进、抑制或无明显影响均有报道[14-15,24],这可能与土壤类型、矿质养分有效性等环境因子相关。本研究结果进一步表明,土壤固氮微生物多样性及群落结构对碳源种类或有效性具有明显响应,对稻草和紫云英等有机物料联合还田利用调控土壤固氮微生物群落结构具有重要指导意义。然而,由于固氮微生物对环境变化十分敏感[8],田间条件下碳源有效性对固氮微生物的调控作用尚需进一步证实。
4 结论
不同碳源(稻草、葡萄糖)配合氮素添加对紫云英季土壤固氮微生物群落具有明显影响。添加秸秆和葡萄糖后增施氮肥,土壤固氮微生物基因拷贝数呈下降趋势。固氮微生物α-多样性对氮素供应的响应在秸秆和葡萄糖处理中不同,即增加氮素供应显著降低了秸秆处理中α-多样性,而添加葡萄糖后增施氮肥则一定程度增加了α-多样性。固氮微生物群落结构因碳源类型差异而聚集为不同组别,受氮水平的影响较小。因此,氮素供应对固氮微生物多样性和群落结构的调控作用主要受碳源种类(或秸秆和葡萄糖中碳源有效性差异)影响。此外,固氮微生物群落结构及优势固氮菌属相对丰度与土壤速效磷含量紧密相关,表明不同碳源添加后造成土壤速效磷含量的差异也可能是驱动紫云英季土壤固氮微生物群落组成的重要环境因子。
[1] 李福夺, 尹昌斌. 南方稻区绿肥生态服务功能及其生态价值评估研究. 中国生态农业学报, 2019, 27(2): 327-336.
LI F D, YIN C B. Assessment of the functions and ecological services values of green manure in paddy fields in South China., 2019, 27(2): 327-336. (in Chinese)
[2] CAI S Y, PITTELKOW C M, ZHAO X, WANG S Q. Winter legume-rice rotations can reduce nitrogen pollution and carbon footprint while maintaining net ecosystem economic benefits., 2018, 195: 289-300.
[3] GALLOWAY J N, TOWNSEND A R, JAN WILLEM E, MATEETE B, ZUCONG C, FRENEY J R, MARTINELLI L A, SEITZINGER S P, SUTTON M A. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions., 2008, 320(5878): 889-892.
[4] HERRIDGE D F, PEOPLES M B, BODDEY R M. Global inputs of biological nitrogen fixation in agricultural systems., 2008, 311(1/2): 1-18.
[5] RAYMOND J, SIEFERT J L, STAPLES C R, BLANKENSHIP R E. The natural history of nitrogen fixation., 2004, 21(3): 541-554.
[6] GABY J C, BUCKLEY D H. A global census of nitrogenase diversity., 2011, 13(7): 1790-1799.
[7] DOS SANTOS P C, FANG Z, MASON S W, SETUBAL J C, DIXON R. Distribution of nitrogen fixation and nitrogenase-like sequences amongst microbial genomes., 2012, 13: 162.
[8] JANGID K, WILLIAMS M A, FRANZLUEBBERS A J, SANDERLIN J S, REEVES J H, JENKINS M B, ENDALE D M, COLEMAN D C, WHITMAN W B. Relative impacts of land-use, management intensity and fertilization upon soil microbial community structure in agricultural systems., 2008, 40(11): 2843-2853.
[9] DELUCA T H, ZACKRISSON O, GENTILI F, SELLSTEDT A, NILSSON M C. Ecosystem controls on nitrogen fixation in boreal feather moss communities., 2007, 152(1): 121-130.
[10] ROESCH L F W, OLIVARES F L, PASSAGLIA L M P, SELBACH P A, de Sá E L S, de CAMARGO F A O. Characterization of diazotrophic bacteria associated with maize: effect of plant genotype, ontogeny and nitrogen-supply., 2006, 22(9): 967-974.
[11] COELHO M R R, de VOS M, CARNEIRO N P, MARRIEL I E, PAIVA E, SELDIN L. Diversity ofgene pools in the rhizosphere of two cultivars of sorghum () treated with contrasting levels of nitrogen fertilizer., 2008, 279(1): 15-22.
[12] 刘骁蒨, 涂仕华, 孙锡发, 辜运富, 张先琴, 张小平. 秸秆还田与施肥对稻田土壤微生物生物量及固氮菌群落结构的影响. 生态学报, 2013, 33(17): 5210-5218.
LIU X Q, TU S H, SUN X F, GU Y F, ZHANG X Q, ZHANG X P. Effect of different fertilizer combinations and straw return on microbial biomass and nitrogen-fixing bacteria community in a paddy soil.,2013, 33(17): 5210-5218. (in Chinese)
[13] GEISSELER D, SCOW K M. Long-term effects of mineral fertilizers on soil microorganisms - A review., 2014, 75: 54-63.
[14] WANG C, ZHENG M M, SONG W F, WEN S L, WANG B R, ZHU C Q, SHEN R F. Impact of 25 years of inorganic fertilization on diazotrophic abundance and community structure in an acidic soil in southern China., 2017, 113: 240-249.
[15] MARTENSSON L, DIEZ B, WARTIAINEN I, ZHENG W, EL-SHEHAWY R, RASMUSSEN U. Diazotrophic diversity,gene expression and nitrogenase activity in a rice paddy field in Fujian, China., 2009, 325(1-2SI): 207-218.
[16] RAHAV E, GIANNETTO M J, BAR-ZEEV E. Contribution of mono and polysaccharides to heterotrophic N2fixation at the eastern Mediterranean coastline., 2016, 6: 27858.
[17] TANAKA H, KYAW K M, TOYOTA K, MOTOBAYASHI T. Influence of application of rice straw, farmyard manure, and municipal biowastes on nitrogen fixation, soil microbial biomass N, and mineral N in a model paddy microcosm., 2006, 42(6): 501-505.
[18] HARADA N, NISHIYAMA M, MATSUMOTO S. Inhibition of methanogens increases photo-dependent nitrogenase activities in anoxic paddy soil amended with rice straw., 2001, 35(3): 231-238.
[19] KONDO M, YASUDA M. Effects of temperature, water regime, light, and soil properties on N-15(2) fixation associated with decomposition of organic matter in paddy soils., 2003, 37(2): 113-119.
[20] WAKELIN S A, GREGG A L, SIMPSON R J, LI G D, RILEY I T, MCKAY A C. Pasture management clearly affects soil microbial community structure and N-cycling bacteria., 2009, 52(4): 237-251.
[21] TANG Y F, ZHANG M M, CHEN A L, ZHANG W Z, WEI W X, SHENG R. Impact of fertilization regimes on diazotroph community compositions and N-2-fixation activity in paddy soil., 2017, 247: 1-8.
[22] HUBBELL S P. Neutral theory in community ecology and the hypothesis of functional equivalence., 2005, 19(1): 166-172.
[23] ADACHI K, WATANABE I, KOBAYASHI M, TAKAHASHI E. Effect of application of glucose, cellulose, and rice straw on nitrogen fixation (acetylene reduction and soil-nitrogen components) in anaerobic soil., 1989, 35(2): 235-243.
[24] YANG L, BAI J S, ZENG N H, ZHOU X, LIAO Y L, LU Y H, REES R M, NIE J, CAO W D. Diazotroph abundance and community structure are reshaped by straw return and mineral fertilizer in rice-rice-green manure rotation., 2019, 136: 11-20.
[25] 鲍士旦. 土壤农化分析. 第3版. 北京: 中国农业出版社, 2000.
BAO S D..Beijing: China Agriculture Press, 2000. (in Chinese)
[26] KABACOFF R.. Manning Publications Co., Shelter Island, NY, USA. 2015.
[27] DE'ATH G. Multivariate regression trees: a new technique for modeling species-environment relationships., 2001, 83(4): 1105-1117.
[28] RAO D N, MIKKELSEN D S. Effect of rice straw incorporation on rice plant growth and nutrition1., 1976, 68: 752-756.
[29] MIRZA B S, POTISAP C, NUESSLEIN K, BOHANNAN B J M, RODRIGUES J L M. Response of free-living nitrogen-fixing microorganisms to land use change in the amazon rainforest., 2014, 80(1): 281-288.
[30] MTAMBANENGWE F, KIRCHMANN H. Litter from a tropical savanna woodland (miombo): chemical composition and C and N mineralization., 1995, 27(12): 1639-1651.
[31] BRANT J B, SULZMAN E W, MYROLD D D. Microbial community utilization of added carbon substrates in response to long-term carbon input manipulation., 2006, 38(8): 2219-2232.
[32] TORRES I F, BASTIDA F, HERNÁNDEZ T, GARCÍA C. The effects of fresh and stabilized pruning wastes on the biomass, structure and activity of the soil microbial community in a semiarid climate., 2015, 89: 1-9.
[33] LIAO H, LI Y, YAO H. Fertilization with inorganic and organic nutrients changes diazotroph community composition and N-fixation rates., 2018, 18(3): 1076-1086.
[34] SCHUTTER M, DICK R. Shifts in substrate utilization potential and structure of soil microbial communities in response to carbon substrates., 2001, 33(11): 1481-1491.
[35] LIN Y X, YE G P, LIU D Y, LEDGARD S, LUO J F, FAN J B, YUAN J J, CHEN Z M, DING W X. Long-term application of lime or pig manure rather than plant residues suppressed diazotroph abundance and diversity and altered community structure in an acidic Ultisol., 2018, 123: 218-228.
[36] WU H P, ZENG G M, LIANG J, CHEN J, XU J J, DAI J, LI X D, CHEN M, XU P, ZHOU Y Y, LI F, HU L, WAN J. Responses of bacterial community and functional marker genes of nitrogen cycling to biochar, compost and combined amendments in soil., 2016, 100(19): 8583-8591.
[37] HALVERSON L J, JONES T M, FIRESTONE M K. Release of intracellular solutes by four soil bacteria exposed to dilution stress., 2000, 64: 1630-1637.
[38] HOLDING A J. The properties and classification of the predominant Gram-negative bacteria occurring in soil. ID - 19621900287., 1960, 23: 515-525.
[39] VITOUSEK P M, CASSMAN K, CLEVELAND C, CREWS T, FIELD C B, GRIMM N B, HOWARTH R W, MARINO R, MARTINELLI L, RASTETTER E B. Towards an ecological understanding of biological nitrogen fixation., 2002, 57/58(1): 1-45.
[40] XUN W B, LI W, HUANG T, REN Y, XIONG W, MIAO Y Z, RAN W, LI D C, SHEN Q R, ZHANG R F. Long-term agronomic practices alter the composition of asymbiotic diazotrophic bacterial community and their nitrogen fixation genes in an acidic red soil., 2018, 54(3): 329-339.
Responses of Soil Diazotroph Community to Rice Straw, Glucose and Nitrogen Addition During Chinese Milk Vetch Growth
YANG Lu1,2, ZENG NaoHua2, BAI JinShun2, ZHOU Xing3, ZHOU GuoPeng1,2, GAO SongJuan4, NIE Jun5, CAO WeiDong2,4
(1Graduate School, Chinese Academy of Agricultural Sciences, Beijing 100081;2Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences/Key Laboratory of Plant Nutrition and Fertilizer, Ministry of Agriculture and Rural Affairs, Beijing 100081;3Crop Research Institute of Hunan Province, Hunan Academy of Agricultural Sciences, Changsha 410125;4College of Resources and Environmental Sciences, Nanjing Agricultural University, Nanjing 210095;5Soil and Fertilizer Institute of Hunan Province, Hunan Academy of Agricultural Sciences, Changsha 410125)
【】This study was to reveal the regulating roles of carbon (C) sources (rice strawglucose) and nitrogen (N) addition in soil diazotroph community during growth of Chinese milk vetch (L.), which is crucial for the management of crop residue and mineral fertilizer application in Chinese milk vetch – rice rotation system in southern China. 【】A pot experiment was conducted with seven treatments, including CK (no exogenous C and N addition), rice straw (Rs) plus various rates of N inputs (Rs, RsN1and RsN2, corresponding to C/N ratios of 66, 25 and 13, respectively), and glucose (Glc) addition plus different N rates (Glc, GlcN1, and GlcN2) with same C quantity and C/N ratios in Rs-included treatments. Soils were sampled during the fast-growing phase of Chinese milk vetch, and destined for characterization ofgene marker and diazotroph community by using the Illumina Miseq PE300 sequencing and PCR techniques.【】Soil C/N ratios were similar between the CK and the treatments with straw or glucose addition alone, while tended to decrease with increasing N input, and significant decrease was observed in the GlcN2relative to CK. Regarding to the available nutrients, comparable soil NO3--N contents were observed under treatments of CK, Rs, and RsN1, but it was significantly increased by 60% under the RsN2treatment. Compared to the CK, Glc-included treatments increased soil NO3--N content by 35%-79%. There were limited variations of soil available phosphorous (P) content under the CK and Rs-included treatments. 16%-24% decrease of soil available P content was found in the Glc-included treatments than that under CK, but not affected by N rates. The copy number ofgene ranged from 80.4×106—140.5×106g-1soil under all treatments. Compared to the CK,gene copy number tended to increase under treatments with both Rs and Glc addition, while a downward trend was observed with increasing N inputs. Exogenous C and N addition resulted in an overall decrease of diazotroph α-diversity than that under the CK. The responses of diazotroph α-diversity to N supply differed between C sources (RsGlc). Compared to Rs alone, RsN1and RsN2had less observed species (decreased by 6%-11%) and Chao 1 index (decreased by 13%-15%), however, GlcN1and GlcN2enhanced α-diversity to some extent relative to Glc alone. PCoA showed that diazotroph community structure was clustered into different groups depending upon C sources, and was marginally affected by N inputs.was the most abundant genus in all treatments, and its relative abundance was significantly reduced by C and N addition in comparison with CK, however, the magnitude of reduction was obviously less in Rs-included treatments than in Glc-included treatments (12.3%-19.7%31.6%-40.5%). In contrasting to, the relative abundance of the second most dominant genus () was markedly increased by C addition relative to the CK, with greater magnitude observed in Glc-includedRs-included treatments (by 170%-270% vs. 25.0%-54.6%, respectively). Meanwhile, Multivariate regression tree analysis, RDA, and Mantel analysis revealed that the diazotroph abundance, diversity and community structurewere closely associated with soil NO3‑-N and available P concentrations. 【】The results suggested that effects of N supply on soil diazotroph abundance, diversity and structure were regulated by C sources or the C availability of rice straw and glucose amendments. Meanwhile, the resulted differences of soil available P availability by various C additions might be a key driving factor of reshaping soil diazotroph community during Chinese milk vetch growth.
soil diazotroph; rice straw; glucose; nitrogen; Chinese milk vetch;
10.3864/j.issn.0578-1752.2020.01.010

2019-04-19;
2019-05-29
国家绿肥产业技术体系(CARS-22)、中国农业科学院科技创新工程、现代农业人才支撑计划(Grant Ref: BB/N013484/1)
杨璐,E-mail:luckyyl_520@163.com。通信作者曹卫东,E-mail:caoweidong@caas.cn,caoweidongnj@njau.edu.cn
(责任编辑 李云霞)
猜你喜欢
右江民族医学院学报(2022年2期)2022-05-19
山东农业科学(2022年3期)2022-04-27
农业科技与信息(2021年16期)2021-12-07
中国农业科学(2021年6期)2021-03-25
安徽农学通报(2021年1期)2021-02-07
上海农业科技(2020年2期)2020-04-02
江苏农业科学(2019年14期)2019-09-23
科学与技术(2019年21期)2019-04-27
中学时代(2018年6期)2018-11-20
海峡姐妹(2018年7期)2018-07-27
