
种群生态风险评估方法研究进展
2020-02-03 11:33谢锐莉许宜平张慧王子健
生态毒理学报 2020年5期
谢锐莉,许宜平,张慧,王子健
1. 中国科学院生态环境研究中心,中国科学院饮用水科学与技术重点实验室,北京 10008 2. 中国科学院生态环境研究中心,环境水质学国家重点实验室,北京 100085 3. 中国科学院大学,北京 100049
人类活动已经对地球生态系统产生了不可逆转的影响,在众多胁迫因子中,化学物质通过对野生生物的毒害作用导致群落和生态系统结构和能流的改变是一个广为关注的命题。工业化过程中产生的大量未知化学品进入环境,许多低浓度化学物质的生态影响经长期暴露才能显现出来,而区域或流域尺度的种群、群落和生态系统的响应需要更长时间才能显示出来,这就需要建立一种有效的方法体系,来替代传统的依据单一生物毒性响应或物种敏感度分布的风险预测或评估方法[1-3]。
生态风险评估的核心目标之一是评估暴露于毒物中的自然种群的动态变化[4]。传统的生态风险评估方法,如评估因子法(assessment factor, AF)和物种敏感度分布法(species sensitivity distribution, SSD)等,均基于单物种的毒性测试,再经外推得出一个敏感物种响应的最低浓度阈值或基准值[1-2,5-6]。在个体水平的实验室标准毒性测试中,实验周期通常较短,且暴露条件往往保持恒定[7]。然而,进入环境的化学物质会受到各种因素的影响,从而导致暴露环境受到不断扰动,短时间内的单代毒性测试无法反映化学物质对生物体的长期累积效应和对物种繁衍历程的影响。其次,个体水平的毒性数据通常来源于多个物种,而不同物种的生活史特征存在差异,比如寿命、生殖时间以及产生的后代数目。这种差异使得生物对化学物质的敏感性不同,即使在外推过程中进行标准化处理也无法消除,从而增加了风险评估过程中的不确定性[8]。最后,生态系统是一个动态的实体,个体之间、个体与环境之间均存在复杂的相互作用,个体层面的风险评估无法真实反映暴露于化学物质中的种群动态变化和物种之间的捕食关系变化[7,9]。因此,需将生态风险评估受体的生物水平拓展至种群以提高评估结果的真实性。
基于种群水平的风险评估方法整合了较低层次生物组织的影响,可反映化学压力源对群落结构的累积效应,减少了生态系统响应外推过程的不确定性[10-11],并将生态系统的复杂性纳入到风险评估中,有助于降低生态风险评估结果的不确定性,从而为环境管理者提出更有效的生态保护条例提供基础参考[12]。本文阐明了生态风险评估从个体水平向种群水平的发展历程,并综述了国内外种群层面生态风险评估的常用技术及模型,希望能够对我国生态风险评估方法和技术发展提供借鉴。
1 种群生态风险评估发展历程(Historical Perspective of population level ecological risk assessment)
生态风险评估的最终目的是保护种群、群落以及生态系统免受污染胁迫因子的威胁[4]。然而传统的基于个体水平的风险评估方法与种群乃至群落层面的风险管理之间存在着持续的脱节[7,12-13]。早期的生态风险评估研究多针对单一风险源或风险因子,评估尺度也局限在单物种水平。这一时期,环境管理中采用的多数化学物质的预测无效应浓度(predicted no-effect concentration, PNEC)均依据实验室产生的单物种毒性测试结果,进而外推到风险熵或概率风险水平。评估因子法和物种敏感度分布曲线法是常用的2类外推方法。然而,由于外推过程中是基于单一物种对胁迫因子敏感度的原理,这2种方法在评估化学物质暴露引起的生态系统响应时存在很大的不确定性。
2005年,美国环境保护局(US EPA)风险评估论坛风险评估委员会(Risk Assessment Forum)成立了一个专门的技术小组,探讨与种群级别生态风险评估相关的一系列问题,最终确定了3项旨在加强种群风险评估的措施,包括扩展种群水平生态风险评估的试验、召开种群水平生态风险评估的方法技术研讨会以及为种群水平生态风险评估制定合适的指南。2008年6月,US EPA风险评估委员会再次召开了一次关于种群水平生态风险评估的技术研讨会,探讨了种群水平生态风险评估的必要性并提出了一些技术方法和模型[14]。随着有毒有害化学品的日益增多以及环境条件的日趋复杂,人们逐渐意识到传统的生态风险评估方法不能有效预测种群、群落以及生态系统所经受的真实风险。与此同时,国内外学者通过风险相关的生物调查、野外毒性毒理实验和生态学模型等方法开展了大量种群水平的生态风险研究,种群水平的生态风险评估也从传统的单一野外观测和生态毒理模型逐渐发展到中宇宙模拟、野外模拟试验和对种群的动态模拟,发展了如生命表分析、Logistic曲线等,模拟种群的分布格局及丰度,进而将生物生理参数纳入模型以预测种群的年龄、阶段、遗传结构和灭绝时间等[15-16]。
2 种群生态风险评估框架(A general framework for population level ecological risk assessment)
Landis和Kaminski[17]提出种群生态风险评估终点应为生态实体和其属性的结合。选择评估种群时应纳入环境保护目标,优先考虑对胁迫源敏感、对生态系统功能影响大的种群。种群属性是种群内个体属性,包括人口统计学特性、栖息地使用、生理和基因属性的综合特性。种群丰度是核心的属性参数,包括种群大小、种群密度、自然状态下的平衡丰度以及环境容纳量。种群增长率描述了种群丰度的时间变化,可反映种群的健康状况,通常用内禀增长率(r)表示。种群结构表征了个体的年龄、大小、性别和发育阶段的分布。空间分布和栖息地属性则反映了种群和环境的交互作用。由于种群属性和个体属性间存在区别,因此评估终点也存在差异(表1)。个体水平的评估终点主要反映个体的存活、生长和繁殖情况,种群水平的评估终点包括种群丰度、密度、内禀增长率、灭绝风险和恢复概率等,更注重种群在胁迫状态下的可持续发展能力。

表1 个体属性和种群属性的对比[18]Table 1 Comparison of attributes between individuals and populations[18]
虽然环境管理者一直将种群作为保护目标,但鲜有研究将种群水平的风险评估和生态风险评估框架结合。Shaffer[19]于1990年提出了种群生存力分析(population viability assessment, PVA)框架,能够识别胁迫源并预测种群持续生存特定时间的可能性,但该框架主要用于濒危物种的保护。Efroymson等[20]于2004年初步提出了种群水平生态风险评估的框架,并应用该框架成功预测了脊椎动物暴露于石油中时种群密度的变化和灭绝的可能性。Barnthouse等[18]在US EPA颁布的生态风险评估框架的基础上正式提出了种群水平生态风险评估(population level ecological risk assessment, PERA)的框架,评估过程与个体水平生态风险评估框架的思路一致。在Barnthouse基础上进一步补充细化的种群水平生态风险评估框架如图1所示。一个系统的种群生态风险评估过程,主要包括风险管理规划、提出问题、风险评估、风险表征和风险管理5个步骤。风险管理规划阶段必须明确管理目标,特定的管理目标能够作为问题提出的起点,为后续的风险分析提供导向。在提出问题阶段,需要确定待评估的种群和评估终点,并选择合适的概念模型和评估种群风险的工具。按照研究对象,种群水平的风险评估方法主要划分为以下2类:(1)直接以种群作为研究对象的评估方法;(2)个体水平毒性数据结合种群模型的外推方法。分析阶段主要是开展实验或输入数据运行模型,判断实验结果和模型输出结果的可靠性。风险表征阶段即利用分析阶段所得的结果进行风险评估,并考虑所用工具和分析过程中的优点和不足。最后将评估结果应用于实际的风险管理中,促进管理目标的实现。

图1 种群水平生态风险评估的框架[18]Fig. 1 A general framework for population level ecological risk assessments[18]
3 直接以种群为研究对象的评估方法(Direct assessment methods based on population attribute)
直接以种群为研究对象的评估方法包括微宇宙、基于eDNA技术的野外群落调查以及关注生物种群结构的非结构化模型和结构化模型,它们可直接通过种群属性对暴露于胁迫源中的种群进行风险评估。
3.1 微宇宙
由于以单物种毒性测试为基础的生态风险评估不能直接反映自然界中的种内和种间关系,且野外规模试验耗费大量人力、物力,微宇宙试验法逐渐受到广泛关注。微宇宙可视为一个小型的生态系统,包含了天然生态系统中的主要物种和生化过程。按照构建的场所,可分为室外微宇宙和室内微宇宙[21]。室外微宇宙与室内微宇宙相比体积更大,营养结构更为复杂,更接近真实的生态系统。室内微宇宙可进一步分为混合培养微宇宙和人工组合微宇宙。混合培养微宇宙内的环境介质和生物种群均来自天然生态系统,可支持多种藻类、微型动物甚至鱼类的生存,而人工组合微宇宙中的环境介质由人工配制而成,在接种生物后,经培养演化,成为小型的生态系统,其优点在于最初的生物种群可以人为控制。
通过微宇宙实验对种群进行生态风险评价时,首先要选择受试生物,在适宜的光照强度和温度下进行微宇宙的构建和驯化,一般应包括3个及以上营养级组分,以反映种群间的捕食和竞争关系。然后设置受试物浓度梯度,进行微宇宙毒性测试。最后在不同时间采集样品,测定理化参数及受试物浓度,并对受试生物进行计数,通过微宇宙内种群密度或丰度的变化反映受试物对种群的毒性效应,通过理化因子的波动指示微宇宙结构和功能的变化。刘建梅等[22]构建了包括浮游藻类、轮虫和大型溞的微宇宙系统,通过定期测定铜离子浓度和种群丰度的变化曲线并结合物种敏感度分布预测了铜离子对该系统的63 d无效应浓度(no observed effect concentration, NOEC)。袁丙强和李少南[23]为探究有机磷杀虫剂三唑磷的水生态风险,构建了室内微宇宙系统,通过冗余分析、蒙特卡罗检验、显著性分析和多重比较得出了三唑磷对于大型溞、萼花臂尾轮虫和绿色湖湾介这3类浮游动物种群的最高无效应浓度和最低有效浓度(the lowest effective concentration, LOEC)。Haegerbaeumer等[24]构建了土壤微宇宙系统,探究了杀真菌剂氟虫腈对线虫的毒性效应。
微宇宙可模拟自然环境中化学物质暴露下的种群动态,以此为基础的生态风险评价更具有真实性。然而由于构建微宇宙所选择的生物通常是易于饲养的,未涵盖全部生态位的生物,因此对受试物的危害评估考虑不足。
3.2 基于环境DNA技术的野外群落调查
环境DNA(environmental DNA, eDNA)技术于20世纪80年代被提出,最早应用于微生物群落研究中,随着第二代测序技术(next-generation sequencing, NGS)的发展,研究对象逐渐拓展,由最初的单物种过渡到了种群、群落水平,现已成为生物监测、物种多样性保护和风险评估领域的非侵入性技术方法[25-28]。eDNA技术是指从环境样品,包括土壤、沉积物、水体和空气等中提取DNA片段,进而利用PCR扩增和高通量测序对环境样品中的目标群落进行定性和定量分析的方法。
利用该技术进行种群生态风险评估时,主要包括环境样品采集、DNA提取和DNA分析3个部分[29-31]。首先,采集环境样本。与传统采样方法不同,浮游植物、浮游动物及鱼类的采集可直接过滤水样,将滤膜保存于无菌冻存管中,采集底栖动物时直接收集沉积物于无菌冻存管中。然后,使用DNA提取试剂盒提取环境样品中的DNA,通过PCR扩增和高通量测序进行可操纵分类单元(operational taxonomic units, OTU)的聚类及物种信息注释。根据后续的数据处理,可得到样本中的物种组成信息,并进一步计算出每个物种的相对丰度。此时,将环境因子或化学物质浓度与种群密度进行相关性分析,可评估种群密度随暴露浓度的波动情况,由此得出该物质的生态风险。Yang等[32]利用eDNA技术鉴定出了太湖流域的浮游动物种群,评估了pH、化学需氧量(COD)、总氮、总磷和硝酸盐等环境因素对浮游动物种群结构的影响,并结合物种敏感度分布推导了特定位点的总氨氮水质基准。该研究团队也将微宇宙实验与eDNA宏条形码技术结合,鉴定出了澳大利亚墨尔本某湿地的939个原核分类单元和878个真核分类群,评估了沉积物中的铜对种群密度、种群结构、生物多样性和生物相互作用的影响[33]。
利用eDNA技术可快速从环境样品中鉴定出多个物种,克服了传统形态学分类的诸多限制。其次,还可与生态学理论结合建模,纳入物种特定的人口统计学特征、污染源的时空变化以及预测种群在污染物胁迫下的反应。但由于该技术费用较高且环境中eDNA技术获得的每个物种的测序序列数与生物量之间的准确定量关系难以确定,因此该技术的使用仍然有限。
3.3 非结构化模型
非结构化模型将种群内个体视为相同个体,用少数变量描述特定种群的动态变化。Malthus模型由英国人口统计学家Malthus在研究历史人口统计资料后提出,最初用于预测人口的变动情况,后被广泛应用于生态学中种群数量波动的研究[34-35]。该模型假设种群内禀增长率(r)保持恒定,种群数量可由式(1)表示,当时间趋于无穷大时,只要r>0,种群数量(Nt)最终一定会趋于无穷大。
Nt=N0er(t-t0)
(1)
式中:Nt表示t时刻的种群数量,N0表示种群初始数量,t表示结束时间,t0表示初始时间。
然而,由于自然界中资源有限且存在种内竞争等因素,种群增长率不可能一直保持恒定,因此该模型的应用存在限制。1838年Verhulst在考虑密度制约的情况下,提出了更符合种群增长规律的Logistic方程[36],如式(2)所示,纳入环境容纳量(K),将种群增长分为起始期、增速期、转折期、减速期和饱和期5个阶段。在此情况下,当时间t趋于无穷大时,种群数量会接近稳定值K。
Nt=KN0/[(N0+KN0)e-rt]
(2)
Hallam和de Luna[37]假设在种群增长率和种群对有毒物质的吸收率之间存在线性关系,提出了生态毒理学上的种群增长模型,运用积分均值法得出了种群平均持续生存的阈值。金香琴[38]利用Logistic模型探究了多环芳烃胁迫对淡水生物种群生长及种间关系的影响。Barnthouse[39]利用离散的Logistic模型预测了枝角类和桡足类等水生生物在农药扰动下种群的恢复时间。
上述Malthus模型和经典Logistic模型多用在单种群的动态分析中,Lotka-Volterra模型以种间竞争关系为基础,表征了2个种群之间互利共生、捕食-被捕食以及相互竞争的关系,能评估多个种群在化学物质暴露条件下的种群动态。Deines等[40]利用Lotka-Volterra模型预测了在外来物种入侵、毒物胁迫和环境变化的条件下,本地物种的种群大小随时间的波动情况以及种群的扩散迁移趋势。由于非结构化模型中,种群内个体均视为相同个体,所以可通过少数变量描述种群特点,但简化的同时也忽略了个体间的差异,导致模型准确度和现实性低。
3.4 结构化模型
结构化模型将种群内个体按照年龄、大小、性别和发育阶段等进行分类,用多组变量描述各类别属性,最经典的是矩阵模型和种群生命表。
Leslie[41]开发的Leslie矩阵是根据个体生命阶段的繁殖力和存活率预测种群增长的模型。该模型将生物种群所有成员按年龄大小等间隔划分为n个年龄组,通过繁殖率和存活率构成的矩阵预测种群的大小、繁殖力和存活率信息。应用该模型进行种群水平生态风险评估时,可通过文献中的历史数据或多代标准毒理学测试获得模型构建的初始参数。Hanson和Stark[42]构建了棉鳚和鲈鱼的Leslie矩阵模型,在密度依赖性和环境随机性条件下,预测了2类种群暴露于壬基酚(NP)、三丁基锡(TBT)、氯化汞(MC)和吡啶硫酮锌(ZnP)中灭绝的可能性及种群的平均恢复时间,结果显示,与传统急性和慢性毒性数据相比,Leslie矩阵方法分别将评估的不确定性降低了80%和61%。Chandler等[43]利用Leslie矩阵构建了桡足类生物暴露于氟虫腈的三代种群模型,结果表明,氟虫腈具有生殖毒性,影响了个体的生殖发育和种群的性别比例。与个体水平的毒性效应评估相比,矩阵模型能够利用有限的数据提高种群水平风险评估的现实性,在种群生态风险评估领域被广泛使用。
生命表是系统记录种群死亡率、存活率和生殖率的一种工具,是最清楚、最直接地展示种群死亡和存活过程的一览表。根据研究对象和数据类型主要分为动态生命表和静态生命表,动态生命表假定种群所经历的环境条件没有变化,真实记录生物同生群个体在各年龄段和发育阶段的存活情况,同时还可以查明和记录死亡原因,从而可以分析种群发育的薄弱环节,找出造成种群数量下降的关键因素,适用于世代不重叠的生物。静态生命表是根据特定时间和区域内种群的年龄分布和存活数据编制而成,种群内个体出生于不同年,经历的环境条件也有所差异,适用于开放、复杂或非人工控制的环境下种群大小和年龄结构趋向稳定的种群,但该模型无法分析死亡原因,也不适用于出生或死亡变动很大的种群。An等[44]基于野生蟑螂的生命周期参数构建了暴露于内分泌干扰物质的种群生命表,结合年龄结构矩阵评估了内分泌干扰物对蟑螂性别变异的影响,并预测双性个体的出现会显著增加种群的灭绝风险。Ducrot等[45]对暴露于锌中的苏氏尾鳃蚓(Branchiurasowerbyi)进行了完整的生命周期测试实验,使用贝叶斯公式拟合出生命表参数,结合矩阵模型预测了种群每日的渐近增长率。运用种群生命表能对种群进行最真实的风险评估,但由于野外观测耗费大量人力物力,因此该方法如今应用较少。
3.5 直接以种群为研究对象的评估方法的优缺点
直接以种群为研究对象的评估方法历经多年的发展,在种群生态风险研究中的应用已较为成熟,在表2中总结了各类方法的优缺点及适用范围。微宇宙能反映化学物质在真实生态系统中的毒性效应,但由于人为驯化和饲养条件的限制,一般只适用于植物、浮游动物、底栖动物和藻类。eDNA技术能快速检测物种在环境中的分布,实验过程简单快速,但费用昂贵。非结构化模型对数据要求低,一般应用于昆虫类种群生态风险的初步评估。一般动物种群的生态风险评估均可采用结构化模型,但由于数据要求高,因此需要大量历史实测数据。
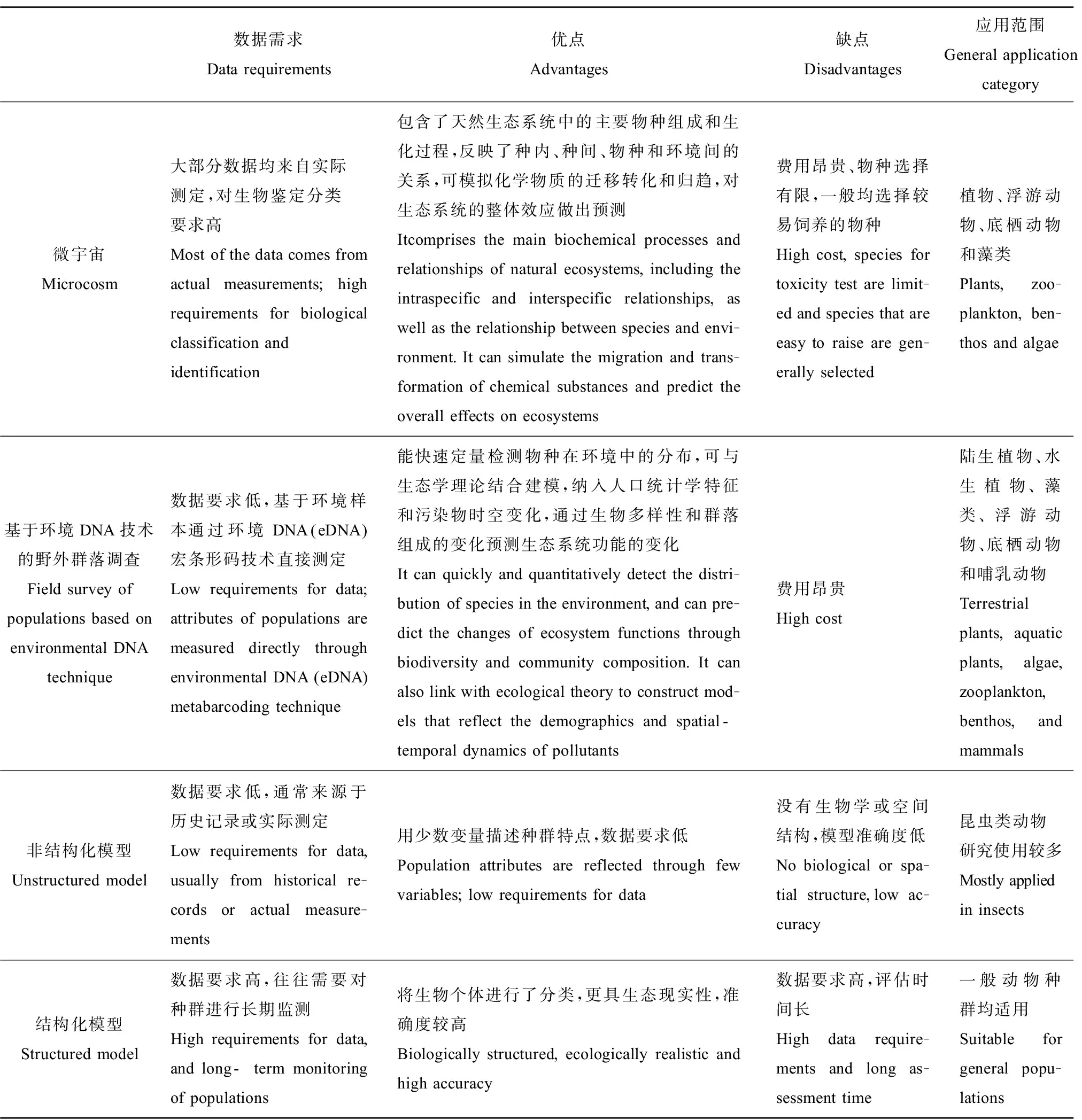
表2 种群水平生态风险评估(PERA)直接评估技术方法和模型比较[2,18]Table 2 The direct techniques and models for population level ecological risk assessment (PERA) [2,18]
4 个体水平毒性数据结合种群模型的外推方法(Extrapolation assessment methods based on individual-level toxicity data and population models)
种群生态风险评估的另一类方法,是以动态能量收支模型(dynamic energy budget, DEB)、生态有害结局路径(ecological adverse outcome pathway, eAOP)以及其他综合生态风险模型等种群生态模型为核心,基于生物个体水平的毒性测试数据,考虑生物和环境介质以及生物之间的相互作用,通过种群模型的动态模拟将个体水平的毒性效应外推至种群水平。
4.1 动态能量收支模型
DEB模型由Kooijman[46]于1986年提出,该模型基于能量代谢的物理、化学特性,量化了能量在生物体整个生活史阶段的分配情况。如图2所示,在标准的DEB模型中,生物体从食物中摄取能量,同化后储存在储存库中,用于生长、成熟和繁殖等生命过程。能量的利用遵循Kappa规则,即储存的能量分为2个部分,其中,固定比例的储存能量(κ)用于体细胞维持和生长(体细胞维持优先于生长),剩余部分的能量(1-κ)用于成熟度维持、成熟和繁殖(成熟度维持优先于成熟和繁殖)。胚胎和幼体利用储备的能量构建生理系统和生殖器官。如果储备的能量不足以维持正常生命活动的能耗,则个体可能会减轻重量或死亡。随着发育停止,当幼体到达成年阶段时,能量将重新分配[47]。

图2 基于动态能量收支(DEB)模型的评估方法[47]注:IBM表示基于个体的模型,f、L0、Lp、Lm、rB和Rm分别表示摄食率、出生体长、成熟时的体长、最大体长、呼吸率和最大繁殖率,CBR表示临界体残留,CTO表示临界靶位点占据,CAUC表示曲线下临界面积,DAM表示损害评估模型,TDM表示阈值损害模型。Fig. 2 The assessment method based on dynamic energy budget (DEB) models[47]Note: IBM stands for individual-based model; f, L0, Lp, Lm, rB and Rm stand for feeding rate, length at birth, length at puberty, maximum length, respiration rate, and maximum reproduction rate, respectively. CBR stands for critical burden residue; CTO stands for critical target occupation; CAUC stands for critical area under the curve; DAM stands for damage assessment model; TDM stands for threshold damage model.
DEBkiss模型是标准DEB模型的简化形式,生物体用于生长和繁殖的所有能量均直接从食物中获取,而不是首先储存在储存库中,该模型具有更多的物种和背景特异性,在理解复杂暴露场景下随时间变化的毒性效应方面非常有用[48]。Billoir等[49]在分析标准大型溞繁殖慢性毒性测试的数据时,在标准DEB模型的基础上,提出DEBtox(dynamic energy budget in toxicology)模型。该模型纳入了毒代动力学(toxicokinetics, TK)和毒效动力学(toxicodynamics, TD)过程,将污染物的暴露浓度作为协变量,生物生活史特征(存活、生长和繁殖)作为时间和暴露浓度的连续变量,通过各种反应机理评估污染物的毒性效应。TK模型能定量反映外源性化学物质在体内的吸收、分布、代谢和排泄等动力学过程,从而预测目标位点的浓度,主要划分为经验房室模型和生理毒代动力学模型两大类[50]。TD模型引入损害动力学,用于描述靶位点浓度变化对特定毒性终点的影响[51]。目前,已开发出许多反映暴露时间和毒性效应之间机制关系的TK-TD模型,包括临界体残留(CBRs)、临界靶位点占据(CTO)、曲线下临界面积(CAUC)、损害评估模型(DAM)和阈值损害模型(TDM)等。DEB模型与TK-TD模型的结合可反映不同化学物质的生理作用模式和毒性作用机制,能对种群所经历的生态风险做出更精准的预测。
利用DEBtox软件及相关毒性数据库,输入个体生理学参数和化学物质毒性参数,运行模型,可得到个体水平的浓度响应曲线和半数致死浓度(LC50)或无效应浓度(NEC)值,结合种群模型,可将个体水平的效应外推到种群水平。首先,DEB模型可以和矩阵模型相结合。在矩阵模型中,个体根据年龄、大小和发育阶段被分为离散的类别,可通过DEB模型中的个体数据来推导每个类别的存活率和生殖率等信息。张慧[52]利用DEB模型建立了稀有鮈鲫在镉暴露下存活、生长和繁殖的胁迫响应关系及能量分配策略,获得了稀有鮈鲫在重金属镉胁迫下的种群安全浓度阈值。Lopes等[53]将摇蚊暴露于甲硫威中进行毒性实验,所得的存活数据作为DEB模型的输入参数,利用DEBtox预测了自然死亡率、致死率、清除率和NEC的值。之后结合Leslie矩阵证实甲硫威浓度超过21 μgL-1时会对摇蚊的种群动态造成影响。其次,DEB模型可与基于个体的模型(individual-based model, IBM)相结合。DEB-IBM模型由Martin等[54]于2012年开发,DEB模型描述个体层面的生理过程,而IBM模型则能够追踪每个个体在不同时间的特征并模拟胁迫源对生理过程或者行为产生效应的机理。它们可以将个体新陈代谢与种群动态联系起来,分析压力源对个体代谢的影响并推断到种群水平。Vlaeminck等[11]利用DEB模型解释个体水平上由于不同食物来源所导致的蜗牛敏感度的差异,通过DEB-IBM模型将食物源效应外推到种群水平,结合生长和繁殖的终点信息,发现抑制能量同化是铜最合理的生理作用模式。
利用DEB模型进行生态风险评估时,不仅能实现个体到种群水平的外推,还能实现不同物种和化学物质间的外推,同时还能评估混合物的毒性效应,但是,构建该模型所需数据量大,物种信息数据库及毒性数据库有待进一步更新。
4.2 生态有害结局路径
生态有害结局路径(ecological adverse outcome pathway, eAOP)是根据化学物质环境暴露的已知信息,结合暴露评估和毒性鉴别,描述分子起始事件与生物组织水平的有害结局之间联系的概念性框架,在化学品环境风险识别及管控中发挥了重要作用。如图3所示,环境中的污染源通过各种介质迁移转化后转移到生物体内,经吸收、分布、代谢和排泄等实现生物体内部暴露,当靶位点浓度达到激活剂量时,分子起始事件经细胞、器官响应后产生个体、种群及生态系统水平的有害结局[55-56]。个体水平的有害结局包括传统毒性终点和非传统毒性终点。传统生物毒性测试可以测定存活、生长和繁殖等毒性终点,它们可以直接用于种群模型的计算中。其他类型的终点,例如与活动、行为、发育和免疫功能相关的参数,只能通过数学关系和机理将其与种群水平相联系,在个体水平有害结局的基础上,通过种群建模阐明高阶群体水平效应。研究者通过AOP将乙酰胆碱酯酶活性的降低与鲑鱼摄食行为、食物消耗和生长联系起来,将个体数据整合到生活史种群矩阵中,从而预测到摄入农药降低了鱼体细胞的生长速率,并使种群数量减少[56-57]。Doering等[58]构建了类二噁英化合物芳香烃受体激活的eAOP框架,可定量预测鱼类及鸟类暴露于此类物质时生命早期阶段的剂量-反应曲线。Hines等[59]基于AOP框架系统收集有关暴露、化学物质性质和毒代动力学的信息,构建总体暴露途径(aggregate exposure pathway, AEP)框架,预测了多种脊椎动物及无脊椎动物暴露于高氯酸根中的有害结局。
在eAOP框架下整合多种暴露途径的累积毒性效应及不同生物组织水平的毒性效应数据,有助于毒理机制研究和风险评估,但目前对于大多数化学物质的生物大分子的分子起始事件以及关键事件的识别仍存在挑战。
4.3 其他综合生态风险模型
综合生态风险模型纳入了环境因子,模拟化学物质在生态系统中的迁移、转化和归趋过程,能在更大的尺度上进行种群水平的风险评估。按照评估尺度可分为基于食物网的模型、基于生态系统的模型和基于社会生态系统的模型[61]。
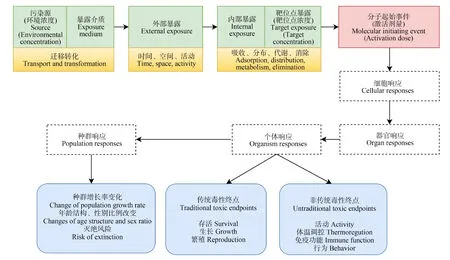
图3 生态有害结局路径(eAOP)概念图[60]Fig. 3 Conceptual framework of the ecological adverse outcome pathway (eAOP) [60]
食物网分析是生态风险评估的重要组成部分,特定的生态食物网所反映的物种间的复杂关系以及时空变化信息能用来评估有毒化学物质以及其他胁迫源的累积效应。常用的模型包括:多介质箱体模型(multimedia box models, MBMs)[62]、AQUATOX模型[63]、三相生态食物网模型(three-phase ecological food web analysis, TEFWA)[64]、基于逸度的食物网模型(fugacity-based food web model, FFWM)[65]。AQUATOX模型由US EPA发布,描述生态系统的概念模型如图4所示,该模型将生产者、消费者、分解者以及相关环境变量纳入其中,构建食物网,模型输入参数包括化学物质的物化性质和毒性参数、环境因子参数以及生物参数,这些参数来自野外实际监测和数据库检索。该模型能模拟氮循环、营养盐的吸收排泄、化学物质的生物积累与转化以及生物的生理生化等过程,能预测由食物网引起的间接生态效应,现已被广泛应用于溪流、池塘、湖泊、河口和水库等各种场景的水生态风险评估中[66]。根据模型输出结果,可预测种群在特定胁迫源下生物量的变化情况,结合后续敏感度分析得到种群生物量变化(±20%)的概率。张璐璐等[67]为探究白洋淀多溴联苯醚(PBDEs)的生态风险,根据淀内优势种群特征和持久性有机污染物的时空分布特征构建了AQUATOX模型,得出了各个种群的生物量变化(±20%)的风险水平,为污染物环境基准的建立提供了较为可行的方法。
基于生态系统的风险评估模型不仅能反映生物间的捕食和竞争关系,同样关注污染源胁迫下生物和环境因子间的相互作用。常用的模型包括:综合水生系统模型(comprehensive aquatic systems model, CASM)[68]、区域脆弱性评估(regional vulnerability assessment, ReVA)[69]、生态网络分析(ecological network analysis, ENA)[70]。CASM模型由Dynamic Solution公司开发,是一个通用的质量平衡水生食物网建模平台,已用于各种淡水和沿海水生生态系统中。输入参数包括基本水质和环境参数、系统总初级生产力、系统呼吸作用、营养物质变化和分解参数、污染物毒性参数等,可以模拟多个种群存在条件下的捕食竞争关系,通过微分方程表示每个状态变量的变化情况,从而预测种群暴露于溶解性化学污染物条件下数量变化的可能性。Naito等[71]利用CASM模型评估了诹访湖浮游植物、浮游动物、底栖动物以及鱼类种群暴露于线性烷基苯磺酸盐中的风险,以生物量变化(±20%)作为潜在毒性终点估算了其NOEC值。
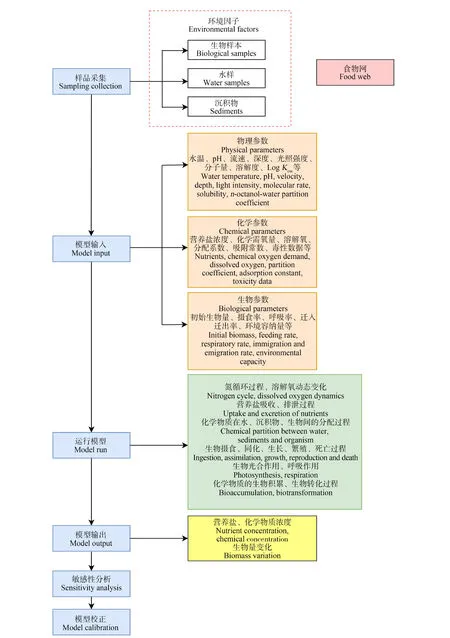
图4 AQUATOX模型的评估流程图[74]Fig. 4 Risk assessment procedure with the AQUATOX model[74]
以上2类模型主要关注种群、群落和生态系统在胁迫源下结构和功能的改变,而社会生态层面的风险评估模型是在法律决策背景下评估自然胁迫源和人为活动的综合风险,包括:净效益分析模型(net benefit analysis model, NBAM)[72]和社会生态系统脆弱性模型(socio-ecological systems vulnerability model, SSVM)[73]。
在生态系统规模上建立的综合生态风险评估模型不仅关注种群间的关系,而且反映了暴露条件下生物与环境因素之间相互作用的改变,在风险管理背景下评估各种生态系统中种群结构和功能响应具有很大潜力。
4.4 个体水平毒性数据结合种群模型的外推方法的优缺点
个体水平毒性数据结合种群模型的外推方法在种群生态风险评估中的应用越来越广泛,笔者总结了上述方法的优缺点和适用范围(表3)。DEB结合矩阵模型和IBM模型能对种群生态风险做出准确的预测,但数据需求量大,模拟过程复杂。eAOP能将毒性效应从分子、细胞水平推至种群水平,利于明确化学物质的毒性作用机制,但由于分子起始事件和细胞关键事件的识别较难,因此适用的化学物质有限。综合生态风险模型适用于大尺度下的种群生态风险评估,更具有生态现实性,但对现场环境因子和生物参数的数据量和准确度要求较高。
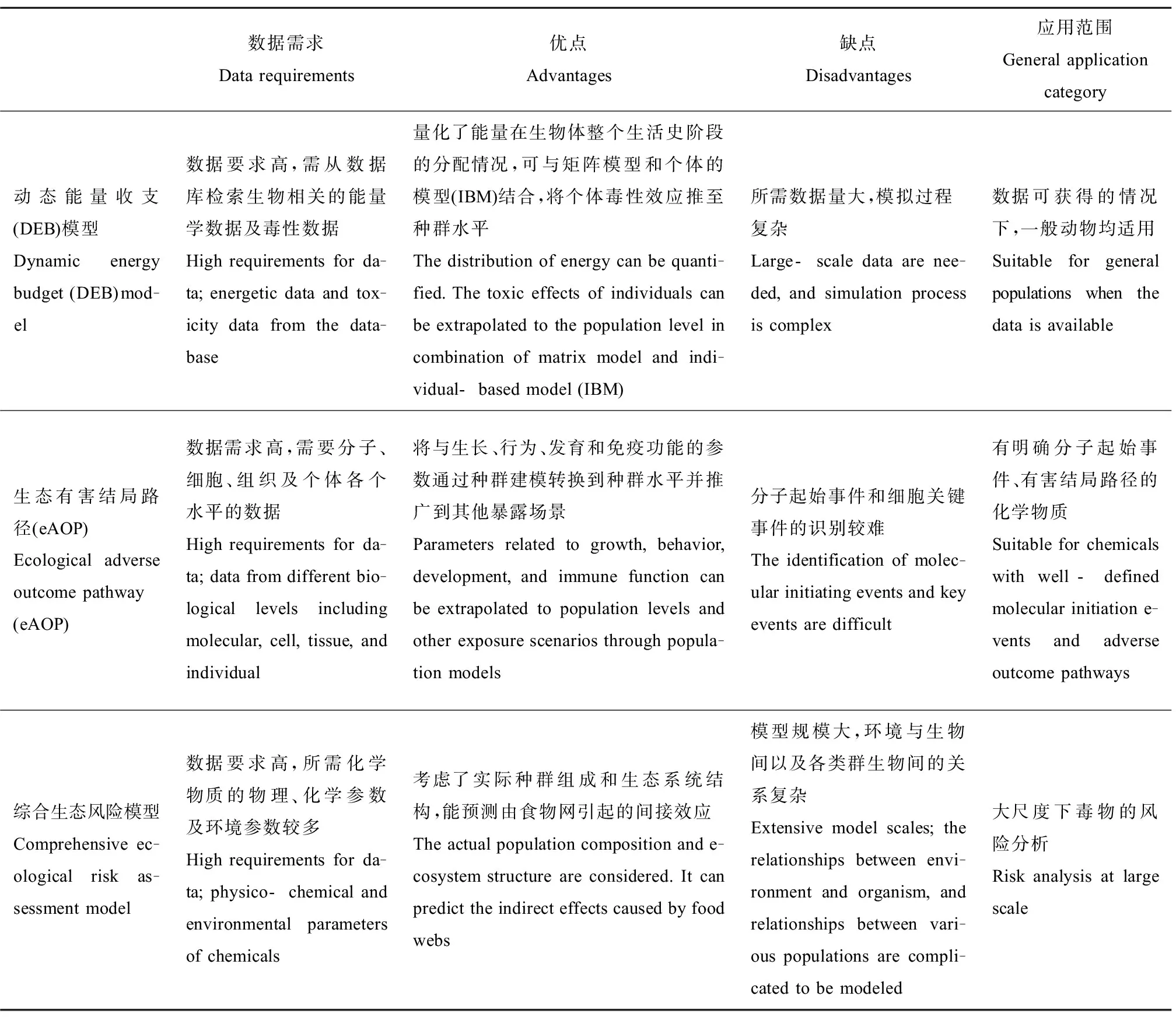
表3 PERA外推模型方法比较[10,47,61]Table 3 The extrapolation techniques and models for PERA[10,47,61]
5 种群生态风险评估的风险表征与不确定性分析(Risk characterization and uncertainty analysis of population level ecological risk assessment)
个体水平的生态风险表征一般采用商值法和概率法。商值法是将物质的环境暴露浓度与表征该物质危害程度的毒性数据相比较,比值>1说明有风险。更高层次的风险评估多采用概率法,通过暴露评估和效应评估的结合,把可能发生的风险依靠统计模型以概率的方式表达出来。目前,种群生态风险表征仍然多采用商值法,根据种群水平的评估终点(例如种群内禀增长率(r)为1、种群生物量变化±20%、种群年龄和性别结构维持稳定)确定种群相应的PNEC值。Lin等[75]通过种群内禀增长率判断青鳉种群暴露于4-壬基酚中的风险,并将r=1时的化学物质浓度作为种群的PNEC值,r<1表明种群受到胁迫,生物量减少。Hanson和Stark[42]将r=1时的化学物质浓度除以评估因子3得到了绵鳚和鲈鱼种群暴露于5种化学物质中的PNEC值,从而评估了种群暴露于化学物质中的风险。
为了对种群所遭受的风险进行综合评估,必须考虑整个评估过程中的不确定性。PERA中的不确定性主要来源于自然变化、模型不确定性和数据不确定性。由于很难在整个时空范围预测所关注变量的真实值,因此自然界本身所固有的不确定性会对评估结果造成影响。针对自然变化的不确定性,可考虑用所有可能的值构建概率分布。通过蒙特卡罗模型将其整合到PERA的模型中,从每个时间阶段的相应概率分布中选择存活率或繁殖力参数,多次运行,最后得到未来种群丰度的概率分布[76]。模型不确定性是由于生态过程或数学分析的简化引入的,数据不确定性包括测量误差和采样实验方法设置不合理等。不确定性的数学表征方法包括概率分析法、区间数学法和模糊理论法。在对不确定性进行分析时,常采用敏感度分析法和采样法。在建模过程中进行敏感度分析时,可多次改变输入参数的值对模型进行扰动,通过模型输出结果的变化鉴别敏感参数。采样法不需要获取模型方程或者模型程序,通过蒙特卡罗和拉丁超立方体抽样法从输入参数的概率密度函数中随机取样,多次运行模型得到模型输出的概率统计分布。
6 种群生态风险评估的应用与展望(Challenges and opportunities of population level ecological risk assessment)
种群水平的生态风险评估历经多年的发展,评估范围已经发展到多种群和群落水平,甚至拓展到景观和区域尺度;评估手段也从传统的对多物种模拟生态系统的监测转化为eDNA分子生物学手段对群落生物多样性的监测以及DEB、综合水生态模型对种群的动态模拟。它可用于环境风险全过程管理体系,不仅能在种群水平上反映突发环境事件的急性风险,而且能用于预测持久性污染物的长期、慢性环境风险,将事件驱动型的环境风险管理转变为主动性的风险预警。种群水平的生态风险评价还可用于环境质量评价,为环境标准的制定和修订提供理论基础。然而,由于种群模型的复杂性和不确定性,以及基础数据的获取需耗费大量物力财力,种群水平的生态风险评估仍停留在科学研究层面,未有效应用到环境风险管理中。随着环境风险管理对种群、群落以及生态系统保护的要求的提高,需从以下几方面开展深入研究,以使种群生态风险评价结果能够更准确地反映生态系统的情况。
(1)拓展PERA的评估框架的理论基础。目前的评估框架侧重于对常规化学物质所导致的生态风险进行评估,缺乏对新型污染物的毒性作用模式研究。通过毒性比率临界值或化学官能团将毒性化学物质区分为反应型、非反应型、非极性麻醉型和极性麻醉型的理论还存在争议,需基于生物体内临界浓度(critical body residue, CBR)衡量外源性化学物质对生物的毒性作用从而建立化学物质分类规则。
(2)优化监测种群属性的技术方法。建立标准化的微宇宙构建方法,包括生物组成、生物培养介质和试验条件,从而提高实验结果的可重复性。优化eDNA的样品的获取及分析方法,避免样品间的交叉污染,使采集样品能全面真实地反映受试种群的属性。进一步发展PCR技术及DNA测序技术,降低eDNA技术的成本。
(3)发展PERA评估模型,优化种群动态模拟技术。在种群模型构建上,需要针对具体问题和数据的可利用度来考虑模型的适用性,模型输出结果应和环境风险管理目标有效结合,表明种群之间、种群和生态系统间的复杂关系,结合毒理学模型,阐明待评估化学物质的作用机制及受体所有可能的暴露途径,结合计算模拟技术,简化模型参数的输入,提高模拟结果的准确度。
(4)判断评估结果的可信度时,应加强证据权重(WoE)方法体系研究。采用证据权重对各证据的相关性、强度和可靠性进行打分,整合生成证据权重主体,以最终确定是否存在不合理的风险。
(5)在风险交流方面,环境风险管理工作者应积极参与评估过程,将评价结果作为风险管理决策的支撑,在不同的环境管理背景下定义可接受的种群风险,补充整合环境风险信息,构建透明有效的环境风险交流平台。
猜你喜欢
今日农业(2022年15期)2022-09-20
中学生数理化·中考版(2022年5期)2022-06-05
中学生数理化·中考版(2021年5期)2021-11-22
当代水产(2021年6期)2021-08-13
湖南电力(2021年1期)2021-04-13
小哥白尼(野生动物)(2019年5期)2019-08-27
中学生数理化·中考版(2019年5期)2019-06-26
中成药(2018年11期)2018-11-24
红土地(2018年7期)2018-09-26
中学生数理化·中考版(2018年3期)2018-05-29
