
雌雄异株地钱(Marchantiapolymorpha)对镉胁迫生理响应的性别差异
2020-01-21 05:59杨茹画金瑭代玉烜王丽
广西植物 2020年11期
杨茹画 金瑭 代玉烜 王丽


摘 要: 为探究雌雄地钱在镉胁迫下性别响应差异,该文采用室内培养法,对野外采集的地钱雌雄配子体进行CdCl2(100 mg·L-1)溶液胁迫处理,定期取样观测其生理指标并在雌雄间进行比较。同时,利用隶属函数法计算雌雄地钱所有指标的综合评价值,评估雌雄地钱耐镉能力差异。结果表明:雌雄地钱在镉胁迫后,各个指标的变化趋势基本一致,超氧化物歧化酶(SOD)、谷胱甘肽(GSH)、丙二醛(MDA)均表现为显著上升,过氧化物酶(POD)呈现为先下降后上升,脯氨酸(Pro)、可溶性蛋白均表现为先上升后下降,叶绿素a(Chla)、叶绿素b(Chlb)均表现为显著下降;雌性地钱叶绿素a/叶绿素b(Chla/Chlb)显著上升,雄性地钱仅在胁迫8 d显著上升,其余天数下无显著变化;综合评价值显示耐镉能力为雄性地钱>雌性地钱。该研究结果为深入探讨雌雄异株植物对胁迫的性别响应差异提供了参考。
关键词: 地钱, 镉胁迫, 性别差异, 隶属函数
中图分类号: Q949.351.108
文献标识码: A
文章编号: 1000-3142(2020)11-1671-10
Abstract: The physiological differences between male and female Marchantia polymorpha under cadmium stress were studied with incubation method in lab. Male and female gametophytes were collected from wild and regularly treated with 100 mg·L-1 of CdCl2 solution, then the cadmium treated samples were collected regularly for determination of physiological indexes. Meanwhile, the comprehensive evaluation value of male and female gametophytes, which calculated by subordinate function method, was used to evaluate the differences in cadmium tolerance. The results were as follows: Each physiological index had a similar trend of change, the activity of SOD, the contents of MDA and GSH increased significantly with stress day, the POD activity increased markedly after an initial decrease, the contents of soluble protein and proline enhanced significantly under short-term stress and then decreased gradually, and the contents of chlorophyll a(chla)and chlorophyll b(chlb)reduced dramatically with stress days; The value of chlorophyll a/chlorophyll b(chla/chlb)of female gametophyte had a significant increase, while it increased significantly only for 8 d in male gametophyte; The comprehensive evaluation value showed the cadmium tolerance ability of male M. polymopha gametophyte was better than female M. polymopha gametophyte. The results provide a reference for the further study of gender differences of dioecious plants under the different stress.
Key words: Marchantia polymorpha, Cd stress, gender differences, subordination function
鎘(Cd)主要来源于废气、污灌、污施,随着工业进程的加快,镉污染也日益严重。镉对植物是非必须元素,在植物体内累积到一定程度会对植物造成毒害,严重阻碍植物生长发育,镉胁迫主要影响植物的细胞分裂、细胞结构、生理生化等(张金彪和黄维南, 1998)。有研究发现低浓度的Cd2+能够刺激 RNA 和蛋白质合成酶的活性,促进细胞分裂进而促进植物生长(曹德菊和汤斌,2004)。高浓度的Cd2+会引起细胞分裂周期紊乱(张瑞凯等, 2018)、染色体发生畸变(赵锦慧等, 2016)等,最终导致细胞分裂受到严重影响。植物受到镉胁迫时,可以调节自身的生理代谢应对胁迫带来的不利影响。低浓度的Cd2+能诱导铜绿微藻(Microcystis aeruginosa)SOD、CAT活性的升高,清除氧自由基进而保护膜系统,而高浓度的Cd2+则会破坏酶结构,使酶失活导致藻体失去清除氧自由基的能力,出现生长受抑制甚至死亡(张海涛等, 2019)。对不同耐镉性的小麦(Triticum aestivum)进行镉胁迫发现,SOD、POD、CAT、MDA含量均随着Cd2+处理浓度的增加而显著上升,胁迫后耐性最低的小麦品系抗氧化酶的活性低于耐性高的两个品种,而MDA含量却高于两个耐性高的品种(夏雪姣等, 2018)。
雌雄异株植物是生态系统的重要组成部分,对保持物种多样性和维持生态系统稳定有重要作用(陈娟和李春阳, 2014)。被子植物中约有5%的雌雄异株植物,分布于900多个属(Ricklefs, 1995),但在野外不同环境下会表现出不同的性别比例,如在水分、养分充足或自然状态下的生境中,其性别比例偏雌性,而在干旱、贫瘠或受到人为因素干扰的生境中,则雄性偏高(陈娟和李春阳, 2014)。并且在不同的胁迫下,雌雄会表现出不同的敏感性和耐受性,这种性别差异性适应在种子植物中已有广泛的研究,如在干旱胁迫下雌性沙棘(Hippophae rhamnoides)(刘瑞香等, 2005)、葎草(Humulus scandens)(刘金平和段婧, 2013)累积更多的脯氨酸、可溶性糖从而表现出比雄性更高的抗旱性。在铅和干旱复合胁迫下,雄性青杨(Populus cathayana)表现出更高的耐性(Han et al., 2013)。苔藓植物独特的构造和生理特征使其对环境敏感性极高,具有很强的吸附、保留重金属的能力(Sun et al., 2007),常用作环境中重金属污染的监测器(李琦等, 2014)。苔藓植物中雌雄异株约有60%( Steere & Schuster, 1984),但在生物监测领域多集中在不同种苔藓对同一重金属或同一种苔藓对不同重金属的富集(朱秀敏等, 2012),对雌雄异株的苔藓植物响应重金属胁迫的性别差异鲜见报道,弄清重金属胁迫下雌雄异株的苔藓植物的性别响应差异,对生物监测的应用、生物多样性的保护、生态系统稳定性的维持都有重要的意义。
地钱(Marchantia polymorpha)隶属地钱科(Marchantineae)地钱属(Marchantia),是苔纲的代表植物(李鹏等, 2017)。对地钱的研究最早主要集中在化学成分,主要是药用成分的研究(Gokbulut et al., 2014),随着基因组数据的发布,对地钱的研究开始转向分子领域,如对Y染色体结构(Okada et al., 2001)、叶绿体基因组(Ohyama et al., 1988)的研究,基因编辑技术的应用(Sugano et al., 2018)等,在胁迫领域,有对酸雨(柴淑丽等, 2018)、光强(樊晓旭等, 2018)、铅(Pb)(张光飞等, 2011)胁迫的报道,但对性别间的响应差异目前仅有酸雨胁迫有涉及(柴淑丽等, 2018),重金属胁迫下雌雄地钱配子体的生理响应及性别间的差异性未见报道。
模糊综合评判是根据模糊数学的理论,通过隶属函数(隶属度)确定各指标间的模糊关系,从而对多个指标进行综合评定,既能消除个别指标的片面性,又由于各指标的隶属函数值是 [0,1]闭区间上的纯数,使不同指标之间及各样本之间的综合抗性差异具有可比性,将它应用于抗性鉴定,可以提高抗性筛选的可靠性(张海燕等, 2005)。目前在抗逆评价上得到了广泛的运用,如赵佳伟等(2019)利用该方法对5个不同品种的北美豆梨(Pyrus calleryana)进行耐盐性评价得出“殿级堂”品种耐盐性最高;陈丽丽等(2019)对小麦(Triticum aestivum)和两种杂草的耐镉、耐铅能力进行综合评价得出在萌发期,两种杂草耐镉、耐铅能力大于小麦(Triticum aestivum);王慧慧等(2018)对10个砧用南瓜(Cucurbita moschata)品种耐镉能力评价得“津油108”耐镉性最强。
本研究采用室内培养法,定期用氯化镉(CdCl2)溶液进行处理,对雌雄地钱配子体的叶绿素含量、SOD、POD、MDA、GSH、Pro、可溶性蛋白等指标进行测定,并在雌雄间进行分析比较,探讨地钱配子体对镉胁迫产生的生理响应差异性;同时,利用模糊数学的隶属函数法,对所有指标进行综合评价,分析雌雄配子体对Cd2+的耐受差异性,以期为深入研究雌雄异株植物对环境的性别响应差异提供参考。
1 材料与方法
1.1 试验材料
供试材料为雌雄野生地钱(Marchantia polymorpha),采自于四川省成都市温江区踏水站林场,经四川大学生命科学学院王丽教授鉴定。根据生殖托挑选雌雄地钱配子体,定植于培养皿中,于25 ℃,光照强度70 μmol·m-2· s-1,光照与黑暗间隔8 h/16 h条件下适应培养一周后开始Cd2+的胁迫处理。
1.2 试验方法
镉胁迫处理液为浓度100 mg·L-1的CdCl2溶液(杜泽萱等,2019),对适应培养后的雌雄配子体喷洒胁迫溶液,每个培养皿每次喷洒10 mL,喷洒时间为每天早晨9:00左右,每隔一天喷洒一次,持续喷洒20 d,每4 d取一次样,每种处理每个取样天数每种性别各设置3个生物学重复,以蒸馏水处理的作为对照组。
1.3 生理指标测定
SOD、POD、 GSH、MDA、Pro、可溶性蛋白测定均参照试剂盒(南京建成生物工程研究所)说明书进行测定。
1.4 叶绿素含量的测定
取处理后的新鲜雌雄配子体,于流水下冲洗干净,并用滤纸吸取表面水分,去除生殖托、假根,用电子天平称取0.1 g用于叶绿素a、叶绿素b含量的测定。将0.1 g配子体与0.9 mL 95%乙醇混合于预先冷冻处理过的研钵中,充分研磨至组织残渣变白,转移至1.5 mL离心管中,低温离心取上清弃去沉淀,将每个上清分别加150 μL于96孔板中,每個样品加3个孔,取95%乙醇作为空白对照,于全波长酶标仪(Spectrophotometer 1510, SN:1510-03010)中测定665、649 nm处的吸光度,根据以下公式计算出叶绿素a(Chla)及叶绿素b(Chlb)的含量。
1.5 数据处理
所有测定的原始数据均用Microsoft Excel 2010处理转化成目标的含量或活性值,采用 SPSS 20.0软件对数据进行统计分析及相关性分析,用平均值和标准误(SEM)表示测定结果,采用单因素方差分析(ANOVA)检验各个处理天数下雌雄配子体各个指标的差异显著性,用LSD法对同一指标测得的数据进行多重比较,统计显著性水平为P<0.05。采用软件GraphPad Prism 5、Microsoft Excel 2010辅助作图。
对雌雄地钱配子体的耐镉能力采用隶属函数综合分析法,与耐镉能力呈正相关的指标,用以下公式进行计算。
2 结果与分析
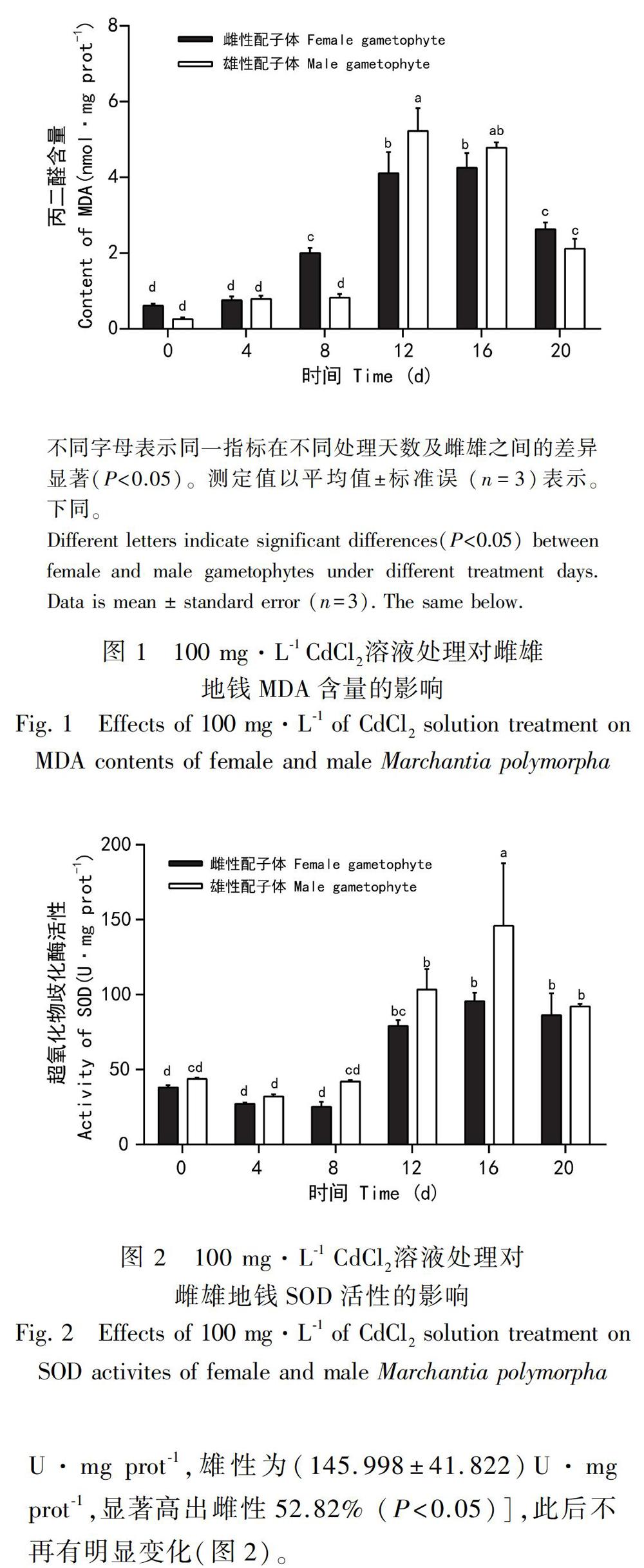
2.1 镉胁迫对雌雄地钱配子体MDA的影响
如图1所示,Cd2+胁迫下,与对照组相比,雌雄地钱配子体体内MDA含量均呈现先上升后下降的趋势,但较对照组均呈现显著上升的趋势(P<0.05)。胁迫8 d时,雌性地钱MDA含量即出现了显著上升达到(2.006±0.132)nmol·mg prot-1,高出对照组229.51%,胁迫16 d时MDA含量达到最大值并显著高出对照组598.36%(P<0.05);雄性地钱在胁迫12 d时MDA含量出现了显著上升并达到了最大值(5.222±0.610)nmol·mg prot-1,高出对照组1947.84%(P<0.05);雌雄地钱在Cd2+胁迫8~12 d时,MDA含量出现了显著的性别差异,但16 d后差异不明显。8 d时雌性显著高出雄性143.15%(P<0.05), 12 d时雄性显著高出雌性27.08%(P<0.05)。
不同字母表示同一指标在不同处理天数及雌雄之间的差异显著(P<0.05)。测定值以平均值±标准误(n=3)表示。下同。
2.2 镉胁迫对雌雄地钱配子体抗氧化酶的影响
雌雄地钱在受到Cd2+胁迫0~8 d后,SOD活性与对照无明显差异,但12 d时显著上升(P<0.05),16 d时达到最高值[雌性为(95.54±3.96)U·mg prot-1,雄性为(145.998±41.822)U·mg prot-1,显著高出雌性52.82%(P<0.05)],此后不再有明显变化(图2)。
与SOD的变化趋势不同,在受到Cd2+胁迫处理后,雌雄地钱体内POD活性均呈现先下降后上升的趋势(图3)。雌性地钱在Cd2+胁迫4 d后,POD活性显著下降到(56.047±2.474)U·mg prot-1(P<0.05),与对照组相比下降了49.22%,在胁迫12 d后POD活性出现了显著上升(P<0.05),并在处理20 d时POD活性上升到最高,显著高出对照组280.23%(P<0.05);雄性地钱在胁迫4、8 d时出现了显著下降(P<0.05),处理8 d达到最低,低于对照组65.09%,在胁迫12 d后POD活性出现显著上升,并且也在处理20 d时上升到最高,显著高出对照组110.82%(P<0.05);雌雄地钱在Cd胁迫处理12~20 d时POD活性出现性别差异,雌性地钱POD活性显著高于雄性地钱(P<0.05)。
2.3 镉胁迫对雌雄地钱配子体Pro、GSH、可溶性蛋白的影响
如图4所示,雌雄地钱在受到Cd2+胁迫之后,Pro含量均出现了先上升后下降的趋势。雌性地钱在Cd2+胁迫处理4~16 d后,Pro含量较对照组均显著上升(P<0.05),并在处理12 d时达到最大值(82.903±1.860)μg·g-1,高出对照组288.74%(P<0.05),在胁迫20 d时,Pro含量下降到显著低于对照组水平(P<0.05);雄性地钱在Cd2+胁迫4~20 d后,Pro含量较对照组均发生了显著上升,并在胁迫8 d时出现了最大值(48.409±0.759)μg·g-1,高出对照组201.75%(P<0.05);在Cd2+胁迫处理0~12 d时,雌雄地钱的Pro含量出现了显著差异,且均表现为雌性含量高于雄性,仅在处理20 d时,雄性地钱Pro含量显著高于雌性(P<0.05)。
由图6可知,地钱在受到Cd2+胁迫后,雌雄地钱体内可溶性蛋白含量均呈现先上升后下降的变化趋势,但在12 d后下降趋势变缓且雌雄间无显著差异。雌性地钱在胁迫4、8 d时,可溶性蛋白含量较对照组显著上升,并在胁迫4 d时达到最大值(0.834±0.025)g prot·L-1,高出对照组51.64%,在胁迫12~20 d持续显著下降,并在20 d时下降到最小值(0.113±0.010)g prot·L-1,显著低于对照组79.45%(P<0.05);雄性地钱在处理4、8 d时,可溶性蛋白含量显著上升,在胁迫8 d时达到最大值(1.022±0.099)g prot·L-1,高出对照组127.62%,在12~20 d时持续显著下降,并在胁迫20 d时下降到最小值(0.161±0.006)g prot·L-1,显著低于对照组64.12%(P<0.05);在胁迫4、8 d时,可溶性蛋白含量在雌雄之间出现显著的性别差异,胁迫4 d时,雌性显著高出雄性20.35%,胁迫8 d时,雄性显著高出雌性27.43%(P<0.05)。
2.4 镉胁迫对雌雄地钱配子体叶绿素含量的影响
雌雄地钱配子体在受到Cd2+胁迫后叶绿素含量发生了显著变化。如表1所示,雌雄地钱Chla含量均呈现显著下降的趋势(P<0.05)。雌性地钱在Cd2+胁迫8 d后,Chla的含量显著低于對照组45.38%,随着处理时间的增加Chla显著下降程度不断加大,在处理20 d时,Chla含量下降到最小值(0.084±0.001 3)mg·g-1,较对照组下降了64.71%(P<0.05);雄性地钱在Cd胁迫4 d时,Chla的含量发生显著下降,达到(0.214±0.000 2)mg·g-1,低于对照组10.83%,在处理20 d时,Chla含量下降到最小值(0.080±0.002 8)mg·g-1,较对照组下降了66.67%(P<0.05);Chla的含量在Cd2+处理4、16 d后雌雄地钱之间均出现了显著差异,雌性地钱在胁迫4 d时显著高出雄性地钱11.21%,雄性地钱在胁迫16 d时显著高出雌性地钱14.13%(P<0.05),到20 d下降到同一水平。
雌雄地钱Chlb的含量同样呈现显著下降的趋势。雌性地钱在Cd2+胁迫4 d时Chlb的含量显著低于对照组27.34%,并随着处理天数的增加显著下降的程度不断加大,到胁迫20 d时,Chlb的含量显著下降到最小值(0.052±0.000 7)mg·g-1,较对照组下降了79.69%(P<0.05);雄性地钱Chlb的含量在Cd胁迫4 d显著低于对照组31.34%,同样到胁迫20 d时,Chlb的含量显著下降到最小值(0.056±0.005 4)mg·g-1,较对照组下降了72.14%(P<0.05);在胁迫0、4 d时,雌雄地钱Chlb的含量出现了显著差异,雌性地钱Chlb的含量显著高出雄性地钱27.36%、34.78%(P<0.05)。
与对照组相比,雌性地钱Chla/Chlb呈现出显著上升趋势;最大值显著高出对照组97.63%(P<0.05);而雄性地钱除了在胁迫8 d时显著高于对照外 [(1.836±0.005 2),高出对照组53.64%(P<0.05)],其余与对照无明显差异;处理0~20 d雌雄间Chla/Chlb无显著差异。
2.5 雌雄地钱配子体耐镉性综合评价
根据隶属函数法计算雌雄地钱每个指标的隶属函数值及所有指标的综合评价值,结果见表2。雌性Chla、Chlb、Pro、可溶性蛋白、POD的隶属函数值均大于雄性,雄性MDA、GSH、SOD、Chla/Chlb隶属函数值均大于雌性,综合评价值雄性(0.529)>雌性(0.483)。
3 讨论与结论
植物在受到胁迫时,在一定范围内可调整自身代谢消除胁迫对植物体造成的伤害从而维持正常生理活动。雌雄异株植物在生长特性、 生理生化特征、 资源分配、 生殖格局、 遗传特性等方面具
有一定的性别差异(Zhang et al., 2012),进而对胁迫的应对能力也会有差异性(刘金平和段婧, 2013)。通过测定这些生理生化指标,可以分析植物的抗逆能力。
脂质过氧化是膜上不饱和脂肪酸发生的一系列活性氧反应,产物丙二醛的含量可以反映脂质过氧化程度(卢倩云等, 2018)。雌雄地钱MDA含量随着胁迫天数的增加,较对照组均显著上升(P<0.05),与镉胁迫小麦的结果一致(夏雪姣等, 2018)。雌性地钱在胁迫8 d时出现MDA的累积,并显著高出雄性143.15%(P<0.05),而此时雄性地钱MDA含量并未出现显著变化,说明雌性地钱膜脂受到氧化伤害早于雄性;16 d后雌雄的MDA与12 d时无太大差异,在20 d时均显著下降(P<0.05),且性别间无明显差异,说明随着胁迫时间的增加,雌雄地钱体内的保护系统受到诱导均能在一定程度上减轻氧化伤害;虽然胁迫12 d雄性地钱MDA含量显著高于雌性地钱,但到20 d,雌雄MDA均下降到同一水平,说明雄性保护系统能更大程度地消除氧化伤害。
植物體内清除ROS自由基的抗氧化系统有2种,一种是植物体的酶保护系统,由SOD、POD、CAT等酶组成;另一种是植物谷胱甘肽抗氧化系统,由GSH、谷胱甘肽还原酶(GR)和谷胱甘肽过氧化物酶(GSH-Px)组成。雄性地钱SOD活性在胁迫16 d时显著高于雌性(P<0.05),说明该胁迫天数下雄性SOD诱导表达程度大于雌性地钱,清除活性氧自由基保护膜系统的能力更强;高浓度的Cd2+能直接取代SOD活性中心的金属元素Fe、Zn等,或是直接与酶的半胱氨酸残基结合,抑制这些酶的活性(程雨卉等, 2017),导致酶活性下降;综合20 d的胁迫,与对照组相比,SOD最大增幅雄性地钱(234.17%)>雌性地钱(150.83%)。POD不仅除能通过氧化还原清除活性氧外,还可参与木质素和木栓质的合成,使细胞壁交联加厚(José Diaz et al., 2001),阻碍重金属离子进入细胞质,从而起到解毒作用;Cd胁迫0~8 d,雄地钱POD活性显著下降(P<0.05),雌性地钱仅在胁迫4 d显著下降(P<0.05),说明短期胁迫下雌雄地钱POD活性表现出下降趋势,可能是地钱配子体体内POD大量消耗,但能够缓解Cd2+胁迫带来的氧化伤害,这与镉胁迫油菜毛状根的结果一致(卢倩云等, 2018);12 d后随着胁迫时间延长,氧化胁迫加剧,POD受到强烈诱导,活性显著上升,在胁迫12~20 d时,POD活性表现出显著的雌性优势(P<0.05)。雌雄地钱GSH含量在胁迫12 d时显著增加,16 d时出现性别显著差异(P<0.05),与对照组相比,雄性地钱的最大增幅(254.49%)>雌性地钱(244.07%),GSH能够与Cd2+ 螯合、沉淀,避免Cd2+在植物体内迁移进而避免与细胞器接触解除Cd的毒性(孙婕妤等, 2018)。因此,依靠GSH消除氧化伤害保护机体的能力雄性地钱强于雌性地钱。
Pro、可溶性蛋白是重要的渗透调节物质,可保护酶的活性(Hare & Cress, 1997)。雌性地钱在镉胁迫前12 d,Pro含量均显著高于雄性(P<0.05),说明雌性地钱累积Pro抗Cd2+胁迫能力强于雄性,Cd2+胁迫时间延长,植物细胞和酶受到破坏,因此,Pro 含量逐渐下降。可溶性蛋白的增加一方面可与Cd2+结合形成Cd-BP固定Cd2+减弱毒性(Salt et al., 1995),另一方面可增加功能蛋白的数量,升高细胞渗透势(李元等, 1990),胁迫4 d雌性地钱可溶性蛋白含量显著高于雄性,胁迫8 d雄性显著高于雌性(P<0.05),综合20 d的结果,雌性地钱可溶性蛋白的最大增幅(51.64%)<雄性地钱(127.62%),到胁迫20 d可溶性蛋白较对照组的降幅雌性地钱(79.45%)>雄性地钱(64.12%),说明雄性地钱依赖可溶性蛋白保护机体的能力强于雌性地钱。
叶绿素a、叶绿素b与植物的光合作用紧密相关,Cd2+胁迫后,雌雄地钱均显著下降(P<0.05),可能是由于Cd2+胁迫使植物体内ROS大量累积,累积的ROS会抑制叶绿素合成相关酶的活性,同时促进叶绿素降解,进而使叶绿素含量降低(生吉萍等, 2009)。胁迫后雌性地钱Chla/Chlb的值较对照组均显著上升(P<0.05),说明雌性地钱体内叶绿素b对镉胁迫更敏感,叶绿素b受到的影响大于叶绿素a,这可能与前人研究发现的叶绿素a/叶绿素b的值上升代表着类囊体的垛叠程度加大,能够缓解光抑制作用(王飞翔等, 2012)有关;地钱属于林下苔藓,对光的捕获能力决定光合作用的强弱,与对照组相比叶绿素a的下降幅度雄性(66.67%)>雌性(64.71%),叶绿素b的下降幅度雄性(72.14%)<雌性(79.69%),可能是由于Cd2+胁迫促进了雄性地钱叶绿素a向叶绿素b的转化,以维持补光能力,而雌性地钱叶绿素b对Cd胁迫更敏感,其捕光能力受到的影响更大,因而光合作用更易受到Cd2+胁迫的影响,且受到的光抑制也比雄性更强。
由上述各个指标可以发现,虽然Cd2+胁迫后,雌雄地钱的各个指标总体变化趋势基本一致,但不同指标在雌雄之间表现出不同的差异性,如POD、Pro表现出雌性优势,但其他各个指标无法看出单一的性别优势,故该研究利用隶属函数法计算了雌雄地钱每个指标的隶属函数值,并将雌雄地钱所有指标的隶属函数值分别加和求均得出雌雄各自的综合评价值,结果表明雄性综合评价值大于雌性地钱,说明雄性抗Cd2+能力强于雌性。
在该研究中,Cd2+胁迫使地钱雌雄配子体抗氧化系统、渗透调节物质、叶绿素含量等生理指标均受到显著影响。虽然地钱雌雄配子体在各指标的总体变化趋势基本一致,但在一些指标上的变化量上存在显著的性别差异。综合评价认为雄性地钱配子体耐镉能力强于雌性,但植物的抗性是多个指标共同作用的结果,既受到遗传因素的影响,又受到诸多外界环境因子影响,同时各指标间又具有一定的相关性,故有关地钱抗Cd2+胁迫表现出的性别差异还应该结合遗传特性、环境因子等进行深入研究。
参考文献:
CAO DJ, TANG B, 2004. Effect of plumbum, cadmium and the combined pollution actate on root tip cell of Vicia faba [J]. Acta Laser Biol Sin, 13(4): 302-305. [曹德菊, 汤斌, 2004. 铅、镉及其复合污染对蚕豆根尖细胞的诱变效应 [J]. 激光生物学报,13(4): 302-305.]
CHAI SL, CHEN YF, FAN XX, et al., 2019. Gender differences of Marchantia polymorpha in tolerance to acid rain stress [J]. Chin J Appl Environ Biol, 25(1):100-107. [柴淑丽, 陈亚菲, 樊晓旭, 等, 2019. 地钱(Marchantia polymorpha)雌雄配子體对酸雨胁迫的耐受性差异 [J]. 应用与环境生物学报, 25(1):100-107.]
CHEN J, LI CY, 2014. Sex-specific responses to environmental stresses and sexual competition of dioecious plants [J]. Chin J Appl Environ Biol, 20(4): 743-750. [陈娟, 李春阳, 2014. 环境胁迫下雌雄异株植物的性别响应差异及竞争关系 [J]. 应用与环境生物学报, 20(4): 743-750.]
CHEN LL, FU YY, WANG YP, et al., 2019. Effects of Cd2+ and Zn2+ on seed germination and seedling growth of Triticum aestivum and 2 weed species [J]. J SW For Univ(Nat Sci Ed), 39(1):50-57. [陈丽丽, 付媛媛, 王艳萍, 等, 2019. 镉锌胁迫对小麦和2种杂草种子萌发及幼苗生长的影响 [J]. 西南林业大学学报(自然科学), 39(1): 50-57.]
CHENG YH, JIANG JW, DONG Y, et al., 2017. Effects of divalent metal ions on activities of immune-related enzymes in Strongylocentrotus nudus [J]. Fish Sci, 36(1): 22-28. [程雨卉, 蒋经伟, 董颖, 等, 2017. 二价金属离子对光棘球海胆免疫相关酶活力的影响 [J]. 水产科学, 36(1): 22-28.]
DU ZX, WU JC, YANG RH, et al., 2019. Single and combined effects of vanadium, titanium and cadmium on the physiology and biochemistry of Marchantia polymorpha L. [J]. N Hortic,(8): 73-81. [杜泽萱, 吴敬陈, 杨茹画, 等, 2019. 钒、钛、镉单一和联合胁迫对地钱生理生化的影响 [J]. 北方园艺,(8):73-81.]
FAN XX, DU ZX, CHAI SL, et al., 2018. Comparison on chlorophyll fluorescence kinetic response of six bryophytes to different light intensity [J]. N Hortic,(20):84-90. [樊晓旭, 杜泽萱, 柴淑丽, 等, 2018. 光强胁迫下六种苔藓植物的叶绿素荧光动力学响应差异 [J]. 北方园艺,(20):84-90.]
GOKBULUT A, SATILMIS B, BATCIOGLU K, et al., 2012. Antioxidant activity and luteolin content of Marchantia polymorpha [J]. Turkish J Biol, 36(4):381-385.
HAN Y, WANG L, ZHANG X, et al., 2013. Sexual differences in photosynthetic activity, ultrastructure and phytoremediation potential of Populus cathayana exposed to lead and drought [J]. Tree Physiol, 33(10):1043-1060.
HARE PD, CRESS WA, 1997. Metabolic implications of stress-induced proline accumulation in plants [J].Plant Growth Regul, 21(2):79-102.
JOS D, BERNAL A, POMAR F, et al., 2001. Induction of shikimate dehydrogenase and peroxidase in pepper(Capsicum annuum L.)seedlings in response to copper stress and its relation to lignification [J]. Plant Sci, 161(1): 179-188.
LI P, LI PF, LI L, et al., 2017. Advances in research of Marchantia [J]. Popular Sci Technol, 19(10):47-49. [李鵬, 李平凤, 黎理, 等, 2017. 地钱属植物研究进展 [J]. 大众科技, 19(10): 47-49.]
LI Q, JI X, WANG EH, et al., 2014. Using bryophytes as biomonitor atmospheric heavy metal deposition in the city of Qingdao [J]. Bull Bot, 49(5):569-577 [李琦, 籍霞, 王恩辉, 等, 2014. 苔藓植物对青岛市大气重金属污染的生物监测作用 [J]. 植物学报, 49(5):569-577.]
LI Y, WANG HX, WU YS, 1990. Effects of Cd and Fe on several physiological indexes of tobacco leaves [J]. Acta Ecol Sin, 12(2):147-154. [李元, 王焕校, 吴玉树, 1990. Cd、Fe及其复合污染对烟草叶片几项生理指标的影响 [J]. 生态学报, 12(2):147-154.]
LIU JP, DUAN J, 2013. Humulus scandens gender differences in response to water stress in the vegetative growth stage [J]. Acta Pratac Sin, 22(2):243-249. [刘金平, 段婧, 2013. 营养生长期雌雄葎草表观性状对水分胁迫响应的性别差异 [J]. 草业学报, 22(2):243-249.]
LIU RX, YANG J, GAO L, 2005. Changes in contents of proline soluble saccharin and endogenous hormone in leaves of chinese seabuckthorn and Russian seabuckthorn under different soil water content [J]. J Soil Water Conserv, 19(3):148-151. [刘瑞香, 杨劼, 高丽, 2005. 中国沙棘和俄罗斯沙棘叶片在不同土壤水分条件下脯氨酸、可溶性糖及内源激素含量的变化 [J]. 水土保持学报, 19(3):148-151.]
LU QY, CAO YC, CHEN YM, et al., 2018. The physiological response and iron and potassium contents in the hairy roots of Brassica rape L. [J]. Chin J Appl Environ Biol, 24(6):1382-1389. [卢倩云, 曹宇棽, 陈友明, 等, 2018. 镉胁迫下油菜毛状根的生理响应及铁钾含量 [J]. 应用与环境生物学报, 24(6):1382-1389.]
OHYAMA K, FUKUZAWA H, KOHCHI T, et al., 1988. Structure and organization of Marchantia polymorpha chloroplast genome: I. Cloning and gene identification [J]. J Mol Biol, 203(2): 281-298.
OKADA S, SONE T, FUJISAWA M, et al., 2001. The Y chromosome in the liverwort Marchantia polymorpha has accumulated unique repeat sequences harboring a male-specific gene [J]. Proc Natl Acad Sci USA, 98(16): 9454-9459.
RICKLEFS RRE, 1995. Dioecy and its correlates in the flowering plants [J]. Am J Bot, 82(5): 596-606.
SALT DE, PRINCE RC, RASKIN PI, 1995. Mechanisms of cadmium mobility and accumulation in indian mustard [J]. Plant Physiol, 109(4):1427-1433.
SHENG JP, LIU KL, SHEN L, 2009. Effects of exogenous nitric oxide on chlorophyll in cadmium-induced tomato seedlings [J]. Spectrosc Spectr Anal, 29(3):762-764. [生吉萍, 劉开朗, 申琳, 2009. 外源一氧化氮对镉胁迫下番茄幼苗叶绿体保护作用的光谱学分析 [J]. 光谱学与光谱分析, 29(3):762-764.]
STEERE WC, SCHUSTER RM, 1984. New manual of bryology [J]. J Ecol, 87(3).
SUGANO SS, NISHIHAMA R, SHIRAKAWA M, et al., 2018. Efficient CRISPR/Cas9-based genome editing and its application to conditional genetic analysis in Marchantia polymorpha [J]. PLoS ONE, 13(10): e0205117.
SUN JY, LIU YQ, LI BL, et al., 2018. Research progress on mechanism of plant tolerance to cadmium and remediation of cadmium contaminated [J]. Jiangsu Agric Sci, 46(7):12-19. [孙婕妤, 刘艳秋, 李佰林, 等, 2018. 植物对镉的耐性机制以及对镉污染土壤修复的研究进展 [J]. 江苏农业科学, 46(7):12-19.]
SUN SQ, WANG DY, HE M, et al., 2007. Retention capacities of several bryophytes for Hg(Ⅱ)with special reference to the elevation and morphology of moss growth [J]. Environ Monit Assess, 133(1-3):399-406. ]
TIAN X, 2018. Evaluation of cold and drought resistance of american popla clones based on subordinate function [D]. Yangling: Northwest Agriculture & Forestry University:7-18. [田星, 2018. 基于隶属函数法的美洲黑杨无性系抗寒、抗旱性评价 [D]. 杨凌: 西北农林科技大学: 7-18.]
WANG FX, WANG MY, LI D, et al., 2012. Effects of Pb and Cd stress on photosynthetic characteristics and contents of pigments in the leaves of Loropetalum chinense var. rubrum [J]. Hunan Agric Sci,(15):103-105. [王飞翔, 王妙媛, 李达, 等, 2012. 土壤铅和镉胁迫对红花檵木叶片光合特性及色素含量的影响 [J]. 湖南农业科学,(15):103-105.]
WANG HH, WANG C, ZHANG MH, et al., 2018. Study on tolerance and accumulation ability of cadmium in pumpkin seedlings for rootstocks [J]. Acta Bot Boreal-Occident Sin, 38(12): 2257-2266. [王慧慧, 王晨, 张明华, 等, 2018. 砧用南瓜幼苗对镉的耐性和积累能力研究 [J]. 西北植物学报, 38(12): 2257-2266.]
XIA XJ, JIAN MY, HAN YC, et al., 2018. Effects of cadmium stress on morphological development and physiological metabolism in wheat(Triticum aestivum)[J]. J Agric Biotechnol, 26(9):1494-1503. [夏雪姣, 菅明阳, 韩玉翠, 等, 2018. 镉胁迫对小麦形态发育及生理代谢的影响 [J]. 农业生物技术学报, 26(9):1494-1503.]
ZHANG GF, DUAN ZZ, LUO XJ, et al., 2011. Response of tolerance and chlorophyll fluorescence characteristics of two bryophyte species to Pb2+ concentration [J]. Environ Poll Control, 33(1):36-40. [张光飞, 段仲昭, 罗晓娟, 等, 2011. 2种苔藓的叶绿素荧光特性及耐受性对Pb2+浓度的响应 [J]. 环境污染与防治, 33(1):36-40.]
ZHANG HT, GUO XY, ZHANG J, et al., 2019. Physiological response of Microcystis aeruginosa to Zn2+ and Cd2+ stresses [J]. Jiangsu J Agric Sci, 35(1):33-41. [张海涛, 郭西亚, 张杰, 等, 2019. 铜绿微囊藻对锌、镉胁迫的生理响应 [J]. 江苏农业学报, 35(1):33-41.]
ZHANG HY, JIAO BC, LI G, 2005. Evaluation of soybean drought resistance [J]. Soybean Sci, 24(3): 183-188. [張海燕, 焦碧婵, 李贵, 2005. 大豆抗旱性鉴定指标评价的研究 [J]. 大豆科学, 24(3):183-188.]
ZHANG JB, HUANG WN, 2000. Advances on physiological and ecological effects of cadmium on plants [J]. Acta Ecol Sin, 20(3):514-523. [张金彪, 黄维南, 2000. 镉对植物的生理生态效应的研究进展 [J]. 生态学报, 20(3):514-523.]
ZHANG RK, ZHANG YH, LIU QQ, et al., 2018. Cadmium induces centrosome amplification through oxidative DNA damage pathway [J]. Lett Biotechnol, 29(2):227-232. [张瑞凯, 张艳花, 刘琴琴, 等, 2018. 重金属镉通过DNA氧化损伤途径诱导细胞中心体扩增 [J]. 生物技术通讯, 29(2):227-232.]
ZHANG S, CHEN LH, DUAN BL, et al., 2012. Populus cathayana males exhibit more efficient protective mechanisms than females under drought stress [J]. For Ecol Manag, 275(4): 68-78.
ZHAO JH, WANG ZQ, HUO SJ, 2016. Effects of heavy metals on seed germination and somatic cell division of broad bean [J]. J Zhoukou Norm Univ, 33(5): 106-110. [赵锦慧, 王智巧, 霍少杰, 2016. 重金属对蚕豆种子萌发及体细胞分裂的影响 [J]. 周口师范学院学报, 33(5):106-110.]
ZHAO JW, LI QY, LU B, et al., 2019. Physiological characteristics and comprehensive evaluation of Pyrus calleryana Decne. under NaCl stress [J]. Plant Physiol J, 55(1):23-31. [赵佳伟, 李清亚, 路斌, 等, 2019. 不同品种北美豆梨对NaCl胁迫的生理响应及耐盐性评价 [J]. 植物生理学报, 55(1):23-31.]
ZHU XM, WANG SH, LI L, 2012. Enrichment capacity of five heavy metals in several bryophytes [J]. N Hortic,(23):88-90. [朱秀敏, 王僧虎, 李丽, 2012. 几种苔藓植物对五种重金属富集能力的比较 [J]. 北方园艺,(23):88-90.]
(责任编辑 周翠鸣)
猜你喜欢
少儿科学周刊·少年版(2021年17期)2021-01-17
少儿科学周刊·儿童版(2021年17期)2021-01-17
阅读(科学探秘)(2020年8期)2020-11-06
小学阅读指南·低年级版(2020年9期)2020-10-12
文萃报·周五版(2019年18期)2019-09-10
儿童故事画报·自然探秘(2019年10期)2019-01-14
小天使·六年级语数英综合(2019年2期)2019-01-12
儿童故事画报·自然探秘(2017年7期)2018-03-14
现代农业研究(2017年10期)2018-01-02
女性天地(2016年10期)2017-04-25
