
耐碳青霉烯类肺炎克雷伯菌毒力基因检测及药敏分析
2020-01-17 01:57邹佳贺刘效伊席健峰
山东化工 2019年24期
邹佳贺,刘效伊,席健峰,徐 义,王 勇*
(1.佳木斯大学附属第一医院,黑龙江 佳木斯 154003;2.深圳市龙岗区妇幼保健院,广东 深圳 518172;3.云台山风景名胜区云台街道卫生院,江苏 连云港 222000)
肺炎克雷伯菌(Klebsiella pneumoniae,KP)属于革兰氏阴性杆菌,近年来已成为引起社区获得性感染和医院感染最重要的病原菌之一,可导致呼吸道、泌尿道、伤口等部位的感染及败血症、肝脓肿等疾病。碳青霉烯类抗菌药物是活性最强、抗菌谱最广的非典型β-内酰胺类抗菌药物,常被认为是治疗革兰阴性杆菌引起感染的最后一道防线[1]。近年来,由于碳青霉烯类抗菌药物的大量使用,导致了耐碳青霉烯类肠杆菌的出现(CRE)。收集2018年6月到2019年6月我院分离出的CRKP菌株42株,检测其毒力基因并对其药敏结果进行分析。
1 材料与方法
1.1 菌种来源
收集2018年6月至2019年6月佳木斯大学附属第一医院从患者标本中分离出的CRE菌株,去除同一患者不同部位相同的菌株共41株。
1.2 仪器与试剂
Vitek-2 Compact全自动微生物分析仪(法国梅里埃公司)。血平板、麦康凯平板(OXOID公司)。
1.3 标准质控菌株
肺炎克雷伯菌ATCC700603购自国家卫生计生委临床检验中心。
1.4 拉丝试验
CRE菌株接种在含5%羊血琼脂平板上,培养18~20h,然后用接种针轻轻挑起菌落,如果挑起菌落过程中出现丝状物并且≥5mm,则为拉丝试验阳性;反之,丝状物长度<5cm则为阴性[2]。
1.5 PCR检测毒力基因
用磁珠法提取细菌的模板DNA,根据文献[3-5]委托上海生工生物技术公司合成毒力基因magA、uge、kfu、allS、iroNB、mrkD、kpn、fimH,阳性产物送上海生工生物有限公司测序,结果在NCBI网站上运用BLAST比对确定其基因型(表1)。

表1 毒力相关基因引物序列

表1(续)
1.6 统计分析
数据使用SPSS 22.0 ,Fisher精确检验进行统计分析,计数资料采用X2检验,以P<0.05 为差异有统计学意义。
2 结果
2.1 CRKP的临床科室分布
42株CRKP中有32(76.2%)株是ESBL阳性菌,10(23.8%)株是ESBL阴性的菌。ESBL阳性菌主要见于重症医学科和神经外科,ESBL阴性菌主要见于重症医学科,见表2。
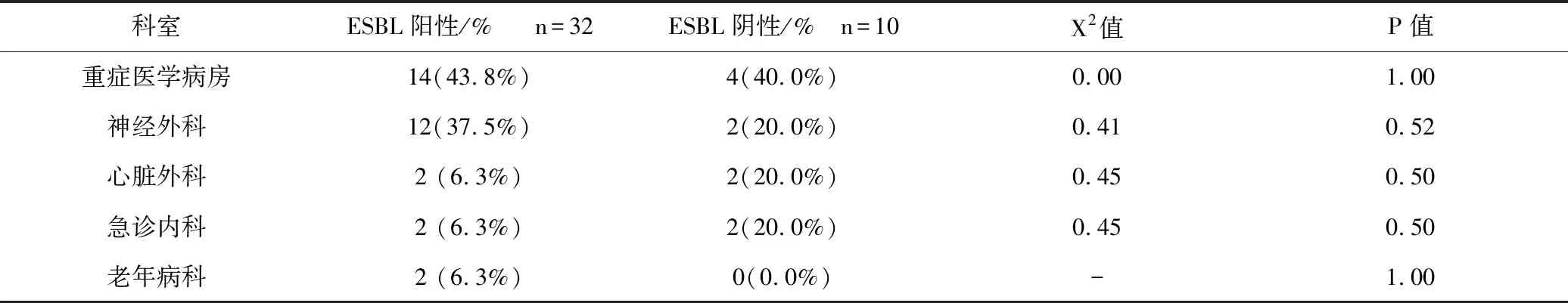
表2 ESBL阳性菌和ESBL阴性菌科室分布
2.2 CRKP拉丝试验结果
对佳木斯大学附属第一医院42株CRKP进行拉丝实验,结果皆为阴性。
2.3 CRKP毒力基因携带情况
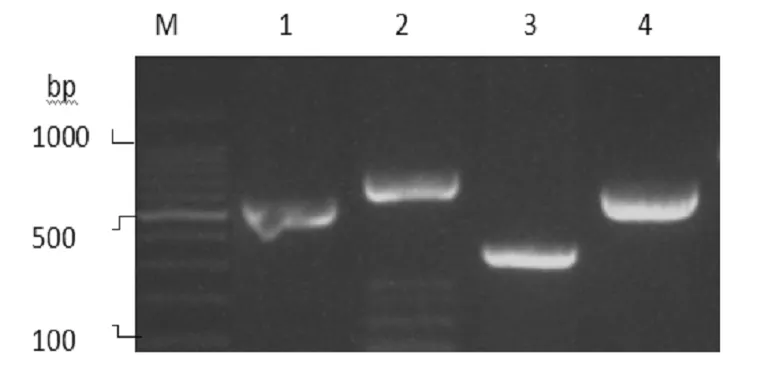
(M:表示mark 1、泳道是uge 2、泳道是fimH 3、泳道是mrkD 4泳道是kpn)
使用PCR方法检测我院42株CRKP,结果42株CRKP经PCR检测fimH携带率为88.1%(37/42),mrkD携带率为92.9%(39/42),kpn携带率为92.9%(39/42);所有菌株均携带uge基因;magA,kfu,iroNB,alls均未检出,见图1。
2.4 CRKP药敏结果
42株CRKP对临常用的抑菌药物如阿莫西林/克拉维酸、哌拉西林/他唑巴坦、头孢呋新、头孢呋辛酯、头孢西丁、头孢他啶、头孢曲松、头孢哌酮/舒巴坦、头孢吡肟、厄他培南、亚胺培南、左旋氧氟沙星、复方新诺明等耐药性显著。对阿米卡星敏感性达100%。以下是 ESBL阳性CRKP菌株和ESBL阴性菌株耐药情况,见表3。
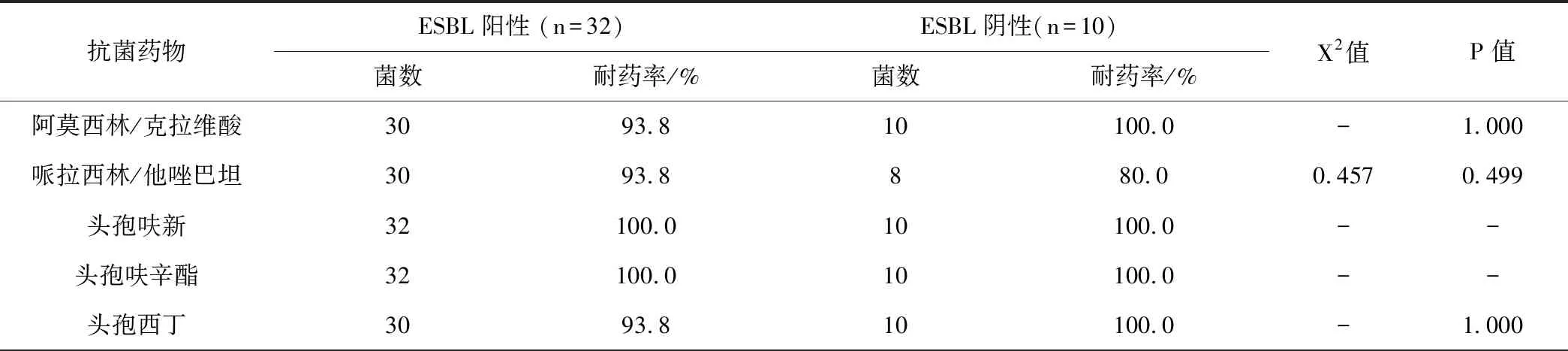
表3 ESBL阳性CRKP菌株和ESBL阴性菌株耐药结果
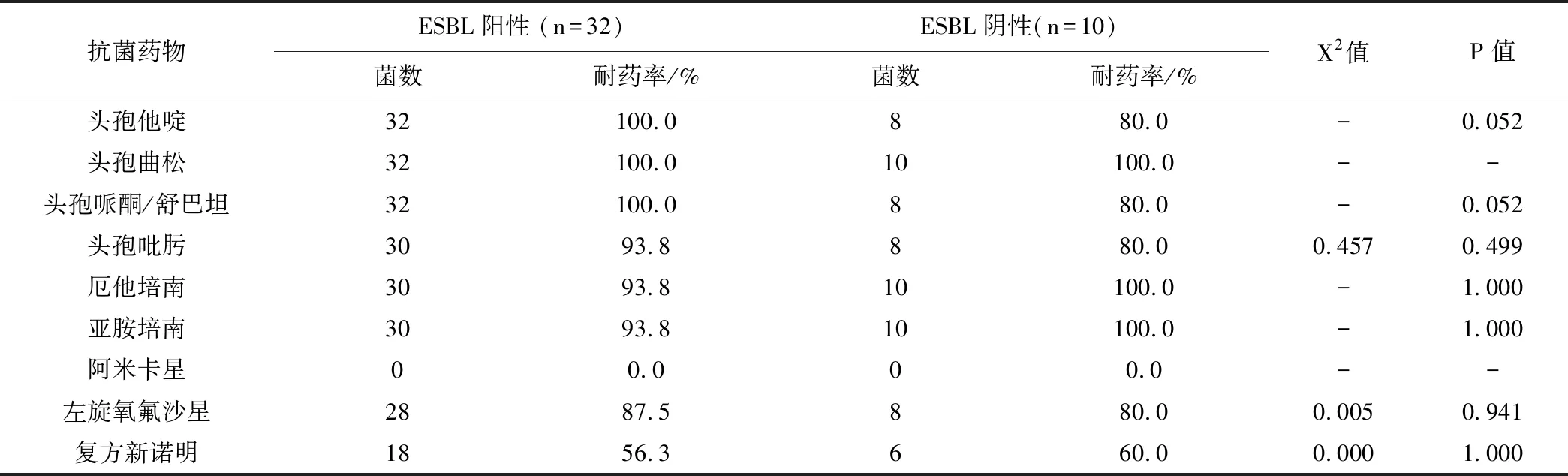
表3(续)
3 讨论
耐碳青霉烯类肺炎克雷伯杆菌(carbapenem-resistantenterobacteriaceae,CRKP)是一种对头孢菌素类、青霉素类甚至是碳青霉烯类抗生素广泛耐药的多重耐药菌,目前CRKP仅对替加环素、多粘菌素敏感。研究表明SARS死亡率为11%,而CRKP感染患者死亡率可达40%~50%[6],感染CRKP的血液病患者死亡率甚至高达60%以上[7],被称为超级耐药菌。由于替加环素在国内还未上市,多粘菌素有很强的毒副作用,应用范围狭窄,CRKP已处于无药可治状态。中国细菌耐药性监测(CHINT)发布报告指出KP对亚胺培南的耐药率由2005年的3%,到2010年的9.2%,再到2016年16.1%,可以看出CRKP成明显增长趋势,且不同地区细菌耐药情况存在差异[8]。由于本地区医院尚未使用阿米卡星这种抗菌药物,所以对阿米卡星敏感性达到100%。肠杆菌科细菌对β-内酰胺类抗菌药物耐药的主要耐药机制之一是产生超广谱β-内酰胺酶(extended-spectrum beta-lactamases,ESBLs)。ESBLs能水解青霉素类、头孢菌素(包含第三代、第四代头孢菌素)及单环酰胺类,如氨曲南,并且可以被β-内酰胺酶抑制剂抑制的由质粒介导的一类β-内酰胺酶。ESBLs主要见于大肠埃希菌和肺炎克雷伯菌,也见于肠杆菌属、变形杆菌属、枸橼酸杆菌属、沙雷菌属等其他肠杆菌科细菌。近年随着抗菌药物大量使用,特别是第三代头孢菌素的广泛使用往往是促使产ESBLs菌株出现及广泛播散的主要原因,各种肠杆菌科细菌甚至都出现了对碳青霉烯耐药的菌株,表现出多重耐药。多重耐药形成的机制比较复杂,主要与产生ESBLs、AmpC酶、碳青霉烯酶等各种重要的灭活酶以及细菌外排系统过度表达、膜通透性改变等机制有关。它们常因携带多种不同的耐药基因而具有多重耐药性,这种耐药性可以通过质粒、转座子等可移动元件在细菌的种内或是种间传播,造成感染爆发流行,控制产酶菌株的流行防止耐药菌谱近一步扩大,已经成为减缓耐药菌众多措施中的重中之重。
CRKP存在高致死率,但它们的毒力因子特征还尚不清楚。Ⅰ型和Ⅲ型菌毛被认为与KP的粘附有关,是另一类较为重要的毒力基因。Ⅰ型菌毛是细菌表面上的线状突起,由fimA-H基因编码。Ⅲ型菌毛是螺旋状的细丝,在KP中由mrkABCD基因簇编码。有研究显示,KP利用环境因素来调节Ⅰ型菌毛的表达,Ⅰ型菌毛在泌尿道中表达,但不在胃肠道或肺中表达,有助于编码Ⅰ型菌毛的hvKP在泌尿道的粘附,引起尿道炎和膀胱[9]。Hornick称Ⅲ型菌毛可以通过mrkD基因介导体外物质与气管、口腔细胞和肺组织结合[10],这与呼吸机相关性肺炎分离的标本中均能检出mrkD基因有关。有研究称无论是hvKP还是cKP都存在Ⅰ型和Ⅲ型菌毛,但也有mrkD基因只在hvKP中检出,并与K2血清型密切相关的报道[11]。究竟编码Ⅰ型和Ⅲ型菌毛的基因是否在特定的KP中检出,需要进一步的研究。根据毒力基因携带情况可知我院的CRKP毒力基因目前只发现了四种。
4 结论
综上所述,应该更加注重对CRKP的耐药性和毒力基因的检测,从而促使临床医生更好的认识和了解CRKP。
猜你喜欢
传染病信息(2022年2期)2022-07-15
现代畜牧兽医(2022年4期)2022-06-09
现代临床医学(2022年3期)2022-06-06
天津医科大学学报(2019年3期)2019-08-13
中国当代医药(2015年26期)2015-03-01
郑州大学学报(理学版)(2014年2期)2014-03-01
中国预防兽医学报(2013年1期)2013-09-11
中国预防兽医学报(2013年10期)2013-09-10
中国动物传染病学报(2013年2期)2013-07-04
