
Anti-microsporidial effect of thymoquinone on Encephalitozoon intestinalis infection in vitro
2020-01-10 06:31lfetetinkayalaySezerArzuvCharyyevaHalilBayraktarHealthVocationalCollegeErciyesUniversityKayseriTurkey
Ülfet Çetinkaya, Gülay Sezer, Arzuv CharyyevaHalil Bayraktar Health Vocational College, Erciyes University, Kayseri, Turkey
2School of Medicine, Department of Pharmacology, Erciyes University, Kayseri, Turkey
3Genkok Genome and Stem Cell Centre, Department of Molecular Biology and Genetics, Erciyes University, Kayseri, Turkey
4Life Science Research Centre, Faculty of Science, Ostrava University, Ostrava, Czech Republic
ABSTRACT Objective: To evaluate the anti-microsporidial effects of the active component of Nigella sativa seeds, thymoquinone, against Encephalitozoon intestinalis using an in vitro model.Methods: Anti-microsporidial effect of thymoquinone against Encephalitozoon intestinalis was evaluated by using various concentrations of thymoquinone (0, 1, 5, 10, 15, 20, 30, 35, and 40 µM) and sterile dimethyl sulfoxide. Real time PCR was used to evaluate the inhibitory effects of thymoquinone on the life cycle of Encephalitozoon intestinalis.Results: The cytotoxic effect of thymoquinone on HEK293 cell line was observed with 30, 35, and 40 µM concentrations of thymoquinone after 24, 48, and 72 hours of incubation. It was observed that 10, 15, 20, and 30 µM concentrations of thymoquinone decreased the spore density compared with the control; however, it was significant only at 30 µM. Conclusions: Thymoquinone shows potent anti-microsporidial effects against Encephalitozoon intestinalis in the in vitro model; however, the toxic concentrations of thymoquinone are also toxic to the host cells.
KEYWORDS: Microsporidia; Thymoquinone; Nigella sativa; Antiparasitic; Encephalitozoon intestinalis
1. Introduction
Microsporidia are obligate intracellular, spore-forming parasites that infect both vertebrates and invertebrates. There are 14 microsporidia species that cause infection in humans. Encephalitozoon spp. and Enterocytozoon bieneusi are considered to be the clinically important pathogens and cause opportunistic infection in immunocompromised individuals[1-3].
Treatment of microsporidia infections is not easy since they are intracellular parasites and have natural resistance from a spore wall. Albendazole and fumagillin are the two most widely used drugs in the treatment of microsporidiosis since they are more effective than other alternative therapeutic agents and less toxic to living cells[4,5]. Albendazole binds to tubulin and inhibits the polymerization of tubulin to microtubules. It remains the drug of choice in treatment although it does not have desirable activity against some microsporidians and in some cases, recurrences are reported[6]. Fumagillin is effective in the treatment of Enterocytozoon bieneusi infections in AIDS patients, so, its use is limited to people with HIV in some countries and has been associated with undesirable effects such as thrombocytopenia and neutropenia[7,8].
Nigella sativa (N. sativa), grown in the Eastern Europe, Middle East, and Western and Central Asia, is used to treat numbers of conditions and diseases such as eczema, asthma, diabetes, hypertension, inflammation, bronchitis, cough, fever, headache, influenza, and dizziness[9,10]. Thymoquinone is an active compound extracted from N. sativa essential oil[11], and has been shown to have anti-bacterial[12], anti-fungal[13], and anti-viral[14] characteristics in various studies. Its anti-parasitic activity has been also investigated against many parasites[15-18]. However, there is no information about the effectiveness of thymoquinone against Encephalitozoon intestinalis (E. intestinalis) or any other microsporidia.
The aim of this study was to investigate whether the active component of N. sativa seeds, thymoquinone, has any antimicrosporidial effects against E. intestinalis using an in vitro model.
2. Materials and methods
2.1. Parasite strain and in vitro infection
The E. intestinalis reference strain (50506) was obtained from the American Type Culture Collection (ATCC). The parasites were cultured in HEK293 cells (human kidney epithelial cells, ATCC) as previously reported[19].
2.2. Chemicals
Thymoquinone was obtained from Sigma-Aldrich (St. Louis, MO) and was dissolved in sterile dimethyl sulfoxide (DMSO, Sigma-St. Louis, MO). After filtration, aliquoted solutions were stored at -20 ℃. The final concentration of DMSO did not exceed 0.1% in the DMSO treated or thymoquinone treated experimental groups. Albendazole (Sigma-St. Louis, MO) was used as a positive control for assaying the life cycle of the E. intestinalis.
2.3. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
HEK293 cells were cultured in DMEM-HG (Sigma-Aldrich) supplemented with 10% fetal bovine serum (Gibco), 1% L-glutamine (Gibco), 1% streptomycin (10 mg/mL)/penicillin (10 000 units/ mL) (Gibco) medium, and incubated under humidified conditions at 37 ℃ and 5% CO2. To establish experimental groups, 3×103cells were seeded into the wells of the 96-well plate with 100 µL fresh medium and incubated overnight. The cytotoxic effect of thymoquinone against HEK293 cells was evaluated by using various concentrations of thymoquinone (0, 5, 10, 15, 20, 30, 35, and 40 µM), and DMSO after incubation for 24, 48, and 72 h. At the end of each period, 10 µL MTT (5 mg/mL stock) solution was added to each well and incubated for 2 h at 37 ℃. Then, 100 µL of DMSO was added to solubilize the purple formazan crystals and the cells were incubated for 20 min at 37 ℃. The absorbance was measured at 560 nm using a microplate reader (Promega Multireader, Glomax, USA).
2.4. Effects of thymoquinone on the life cycle of E. intestinalis
To evaluate the inhibitory effects of thymoquinone on the life cycle of E. intestinalis, HEK293 cells were used as a host cell. HEK293 cells were cultivated in a 6-well tissue culture plate and incubated at 37 ℃ in 5% CO2overnight. After removal of non-adherent cells, E. intestinalis spores were added and incubated at 37 ℃ in 5% CO2overnight for infection. After incubation, non-adherent spores were removed with the medium. Fresh medium containing various concentrations of thymoquinone (0, 1, 5, 10, 15, 20 and 30 µM), albendazole as a positive control (8 and 16 ng/mL), or DMSO was added to each well and incubated at 37 ℃ in 5% CO2for 10 d with the media changed twice a week. Ten days after the infection, spores started to be detectable in the medium and the experiments were terminated. To quantify parasite density, all media and host cells were removed from the cell culture plate with trypsin (Sigma-St. Louis, MO) and collected in a tube. The tubes were then centrifuged at 4 000×g for 10 min and the supernatant was removed. The pellet was used for DNA isolation. DNA was prepared using the GeneAll®Exgene Cell SV Mini Kit (GeneAll Biotechnology, Seoul, South Korea) according to the manufacturer’s recommendation. The parasite load was evaluated quantitatively by real-time PCR using the Roche LightCycler®480 system (Germany).
2.5. Real-time PCR
The previously reported primer pair was used to detect microsporidia[20]. The total volume of the reactions were 20 µL containing 5 µL of extracted DNA, 1 µL of both forward and reverse primers (10 pmol), 10 µL 2×SYBR-Green Master (Roche Diagnostic, Germany), and 3 µL of PCR grade water. The thermal cycles for real-time PCR reaction consisted of the following steps: initial incubation at 95 ℃ for 5 min, 45 cycles at 95 ℃ for 10 s, 60 ℃ for 10 s and 72 ℃ for 10 s, followed by 1 cycle at 95 ℃ for 5 s, 65 ℃ for 1 min, and finally 1 cycle at 40 ℃ for 30 s.
The dilution series prepared from the plasmid encoding E. intestinalis 16S SSU rRNA region were used as a positive control. The number of copies in the main stock dilution was calculated to be 2.8×108. Then, ten different dilutions were prepared by making a ten-fold dilution from the main stock.
2.6. Statistical analysis
All experiments were carried out in triplicate and data was presented as mean ± standard error of the mean (SEM). Statistical analyses were carried out with the SPSS statistical software package (version 21.0, Inc., Chicago, IL, USA) and graphics were prepared with the Prism GraphPad (San Diego, CA, USA). After checking for the homogeneity of variance using the Levene’s test, one-way ANOVA was used, followed by post hoc Tukey test to compare the absorbance and parasite load values of different thymoquinone concentrations with control group or albendazole treatment. P<0.05 was considered statistically significant.
3. Results
3.1. Cytotoxic effect of thymoquinone on HEK293 cells
The cytotoxic effect of thymoquinone on HEK293 cell line was observed at 30, 35, and 40 µM concentrations of thymoquinone after 24, 48, and 72 hours of incubation period. Compared to the control group, DMSO, or thymoquinone at the concentration less than 30 µM did not have significant effect on cell viability (P>0.05) (Figure 1).
3.2. Effects of thymoquinone on life cycle of E. intestinalis
As shown in Figure 2, serial dilutions were appropriate to determine the density of the parasites. After 10 d, the experiments were terminated and the parasite load was assessed by quantitative real time PCR.
It was observed that 10, 15, 20, and 30 µM concentrations of thymoquinone decreased the spore density compared with the control (Figure 3); however, it was significant only at 30 µM (P<0.05) (Figure 3). The effect of 30 µM thymoquinone was similar to 8 ng/mL albendazole (P>0.05).
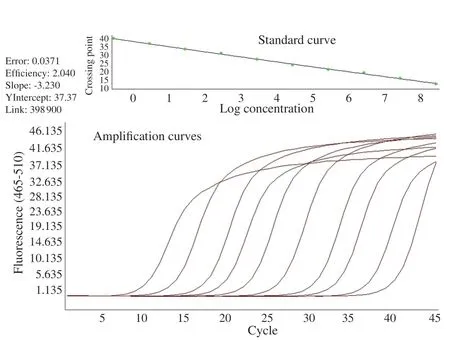
Figure 2. Real-time PCR results. Dilution series prepared from the plasmid encoding of Encephalitozoon intestinalis 16S SSU rRNA region were used as a positive control. The number of copies in the main stock dilution was calculated to be 2.8×108. Then, ten different dilutions were prepared by making a ten-fold dilution from the main stock.
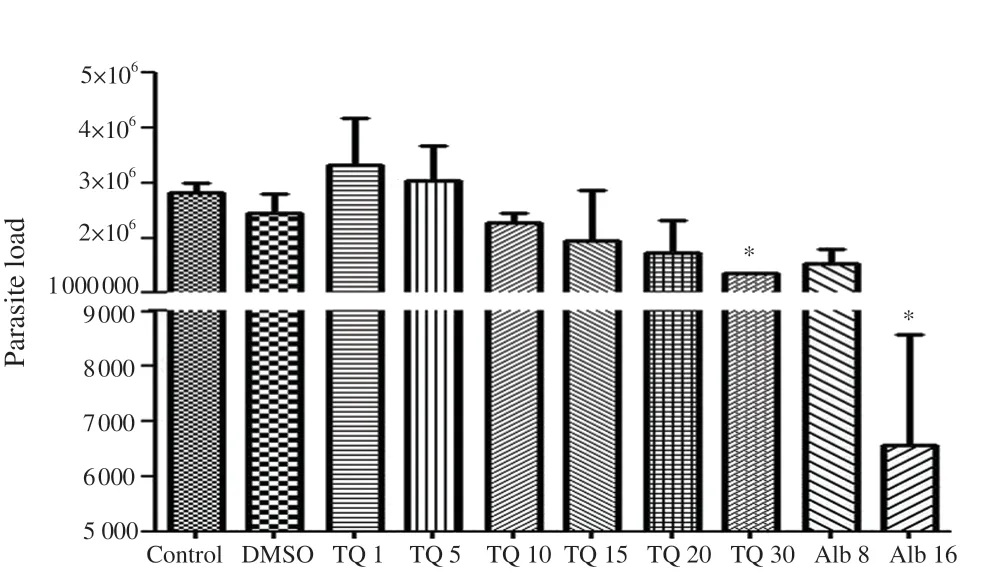
Figure 3. Real-time PCR evaluation of the inhibitory effects of thymoquinone (TQ) on the life cycle of Encephalitozoon intestinalis after ten days of incubation with TQ at 0 (control), 1, 5, 10, 15, 20, and 30 µM concentrations, albendazole (Alb) at 8 and 16 ng/mL of concentrations, and DMSO. DMSO was used to dissolve TQ and albendazole. *P < 0.05 compared to the control on the 10th day.
4. Discussion
Albendazole is the most effective drug in the treatment of microsporidia to date. Recent studies have reported that this drug limits the parasite burden, however, fails in clearance of microsporidia especially in immunosuppressed individuals and can result in the reoccurrence of the disease after the treatment[21,22]. Fumagillin is reported to be effective in the treatment of Enterocytozoon bieneusi but has some side effects[7,8]. Some medications such as azithromycin, metronidazole, paromomycin, nitazoxanide, and cyclosporine have been used in treatment, but no effective results have been obtained[22,23].
Plant extracts and compounds have been used as a valuable natural resource of traditional remedies for centuries. In previous studies, it was reported that thymoquinone shows an anti-parasitic effect against some parasites. In one study[18], it was reported that thymoquinone showed anti-leishmanial activity (Leishmania infantum, IC50: 1.47 mg/mL and Leishmania tropica, IC50: 1.16 mg/mL) on promastigotes after 72 hours of incubation period. In the same study, it was observed that the parasites had less macrophage invasion ability and intracellular survival rate of intramacrophage amastigotes (Leishmania infantum, IC50: 2.6 mg/mL and Leishmania tropica, IC50: 2.1 mg/mL) in the presence of thymoquinone. Thymoquinone showed an inhibitory effect on Babesia and Theileria proliferation in both in vitro and in vivo. These results make thymoquinone a promising candidate for use in therapy. Other authors reported IC50values of thymoquinone were (0.28 ± 0.016), (7.35 ± 0.17), (35.41 ± 3.60), (67.33 ± 0.94), and (74.05 ± 4.55) µM for Babesia divergens, Babesia bigemina, Babesia bovis, Babesia caballi, and Theileria equi, respectively[24]. In addition, Mahmoudvand et al.[17] revealed that thymoquinone can be a new scolicidal agent for use in hydatid cyst surgery. They reported that all of the protoscoleces died after exposure at the concentration of 1 mg/mL at 10 min. In this study, the anti-microsporidial effect of thymoquinone was evaluated for the first time. It was found that 30 µM thymoquinone inhibited the life cycle of E. intestinalis. In addition, the amount of spores was significantly decreased ten days later. In this study, unlike against leishmania[18], the lower concentrations of thymoquinone (30 µM for intracellular forms) were found to be more toxic for intracellular forms than extracellular spores. This can be explained by the spore structure of the parasite. While microsporidian parasites have highly resistant spores, they do not have a wall structure in its intracellular forms. For this reason, higher concentrations of thymoquinone and long term exposures are necessary to change the viability of spores. The cytotoxic effect of thymoquinone on host cell has been reported by many researchers[25-27]. The findings of the present study demonstrated that thymoquinone is cytotoxic on HEK293 cells at concentrations of 30 µM. Since microsporidia are obligatory intracellular parasites requiring host cells for their reproduction, the studies on the host cell and the life cycle of the parasite showed that thymoquinone has toxic effects at 30 µM concentrations. In this case, it is hard to understand whether the decreased number of spore was caused directly by the toxic effect of thymoquinone on the spore or host cells.
Despite studies demonstrating an anti-parasitic effect of thymoquinone, there are some studies reporting that thymoquinone has no significant effect on parasite survival. Khan et al. reported that N. sativa has a weak effect on balantidiasis in donkeys[15]. Similarly, Nasir et al. proved that N. sativa shows poor effect on treatment of Cryptosporidium parvum infection in calves[28]. This can be explained in several ways; (1) In some studies, N. sativa extracts were used instead of thymoquinone. Some studies reported that thymoquinone is more effective than N. sativa extracts, (2) Studies were conducted only in vivo. (3) It may have different effects on different parasites.
In summary, thymoquinone showed potent anti-microsporidial effects against E. intestinalis in the in vitro model, however, the toxic concentrations of thymoquinone are also toxic to the host cells. Thymoquinone in combination with other medications or formulations as nanoparticles can be evaluated for further studies. It should be noted that these are the main limitations in the present study. Despite these limitations, this study gives a general idea of the use of thymoquinone.
Conflict of interest statement
We declare that there is no conflict of interest.
Authors’ contributions
UC, AC and GS designed the study and made the critical revision of the article. UC and GS performed the in vitro experiment. Collecting test data, drafting the article and getting a final approval of the version to be published were done by UC and GS, as well as data analysis and interpretation was done by GS. In addition, UC was responsible for supervision, and project administration.
 Asian Pacific Journal of Tropical Biomedicine2020年1期
Asian Pacific Journal of Tropical Biomedicine2020年1期
- Asian Pacific Journal of Tropical Biomedicine的其它文章
- Hepatoprotection by dandelion (Taraxacum officinale) and mechanisms
- Renoprotective effect of umbelliferone in high-fat diet/streptozotocin-induced type 2 diabetic rats
- Raspberry ketone attenuates high-fat diet-induced obesity by improving metabolic homeostasis in rats
- Evolution of specific RNA aptamers via SELEX targeting recombinant human CD36 protein: A candidate therapeutic target in severe malaria
- Antioxidant and anti-melanogenic activities of ultrasonic extract from Stichopus japonicus